
Statistical Optimisation of Diesel Biodegradation at Low Temperatures by an Antarctic Marine Bacterial Consortium Isolated from Non-Contaminated Seawater


 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
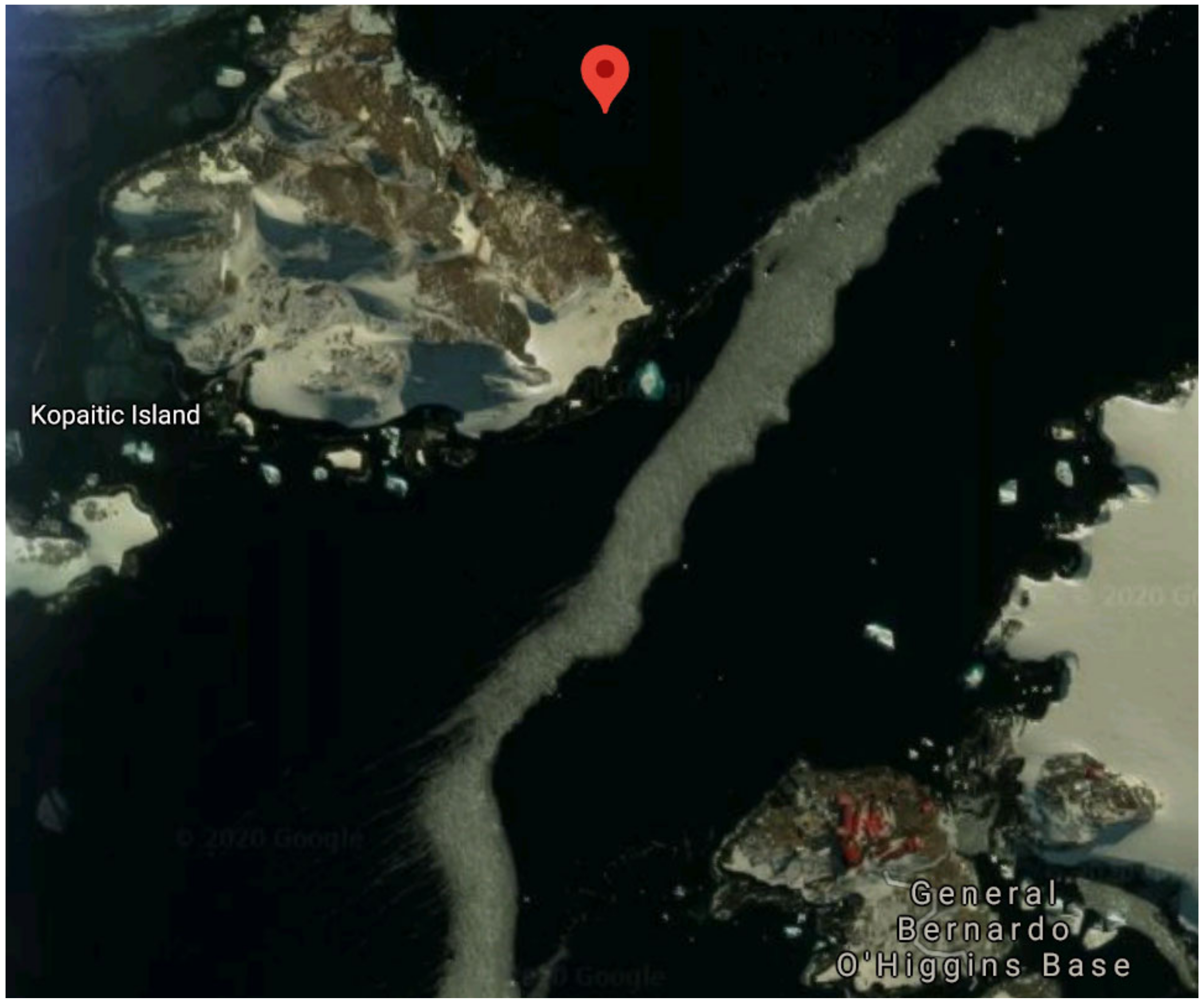
2.1. Sampling and Media
2.2. Screening for Diesel Hydrocarbon Biodegradation
2.3. Optimisation of Diesel Degradation Using One-Factor-at-a-Time (OFAT)
2.4. Quantification of Diesel Degradation
2.5. Response Surface Methodology
2.5.1. Selection of Significant Variables by Plackett-Burman Design
2.5.2. Central Composite Design
3. Results
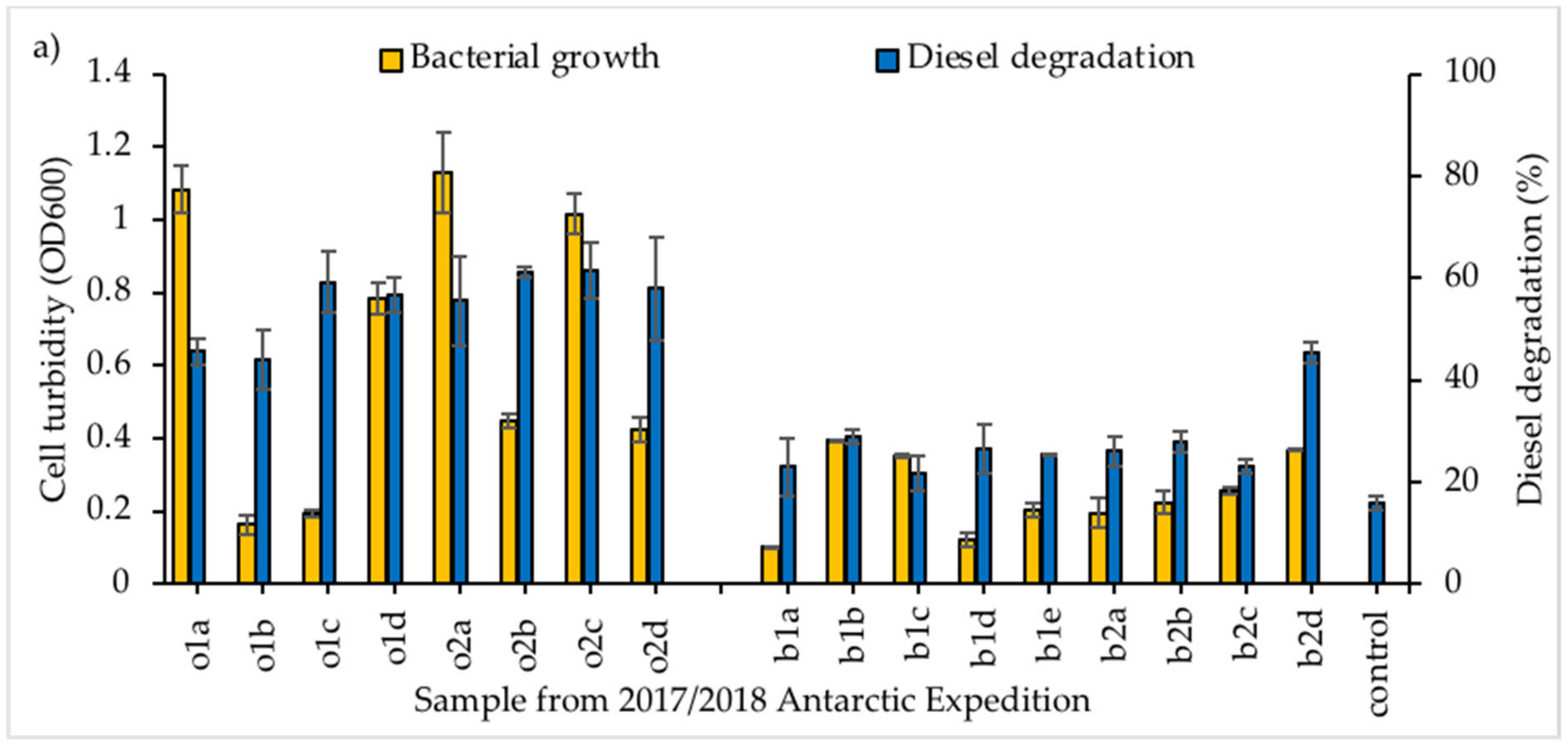
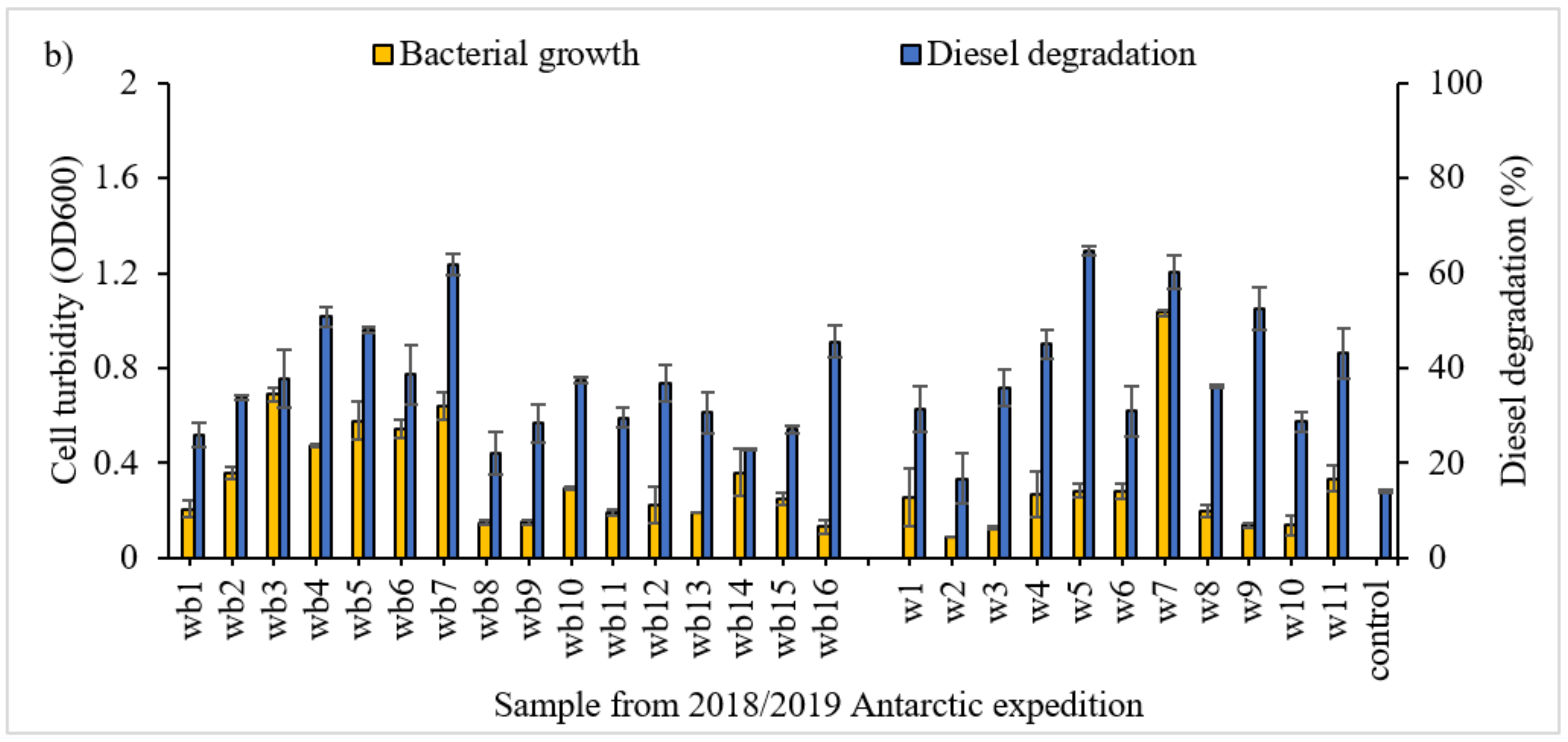
3.1. Screening of Diesel Degrading Consortia
3.2. Optimisation of Growth Conditions Using Conventional One-Factor-at-a-Time
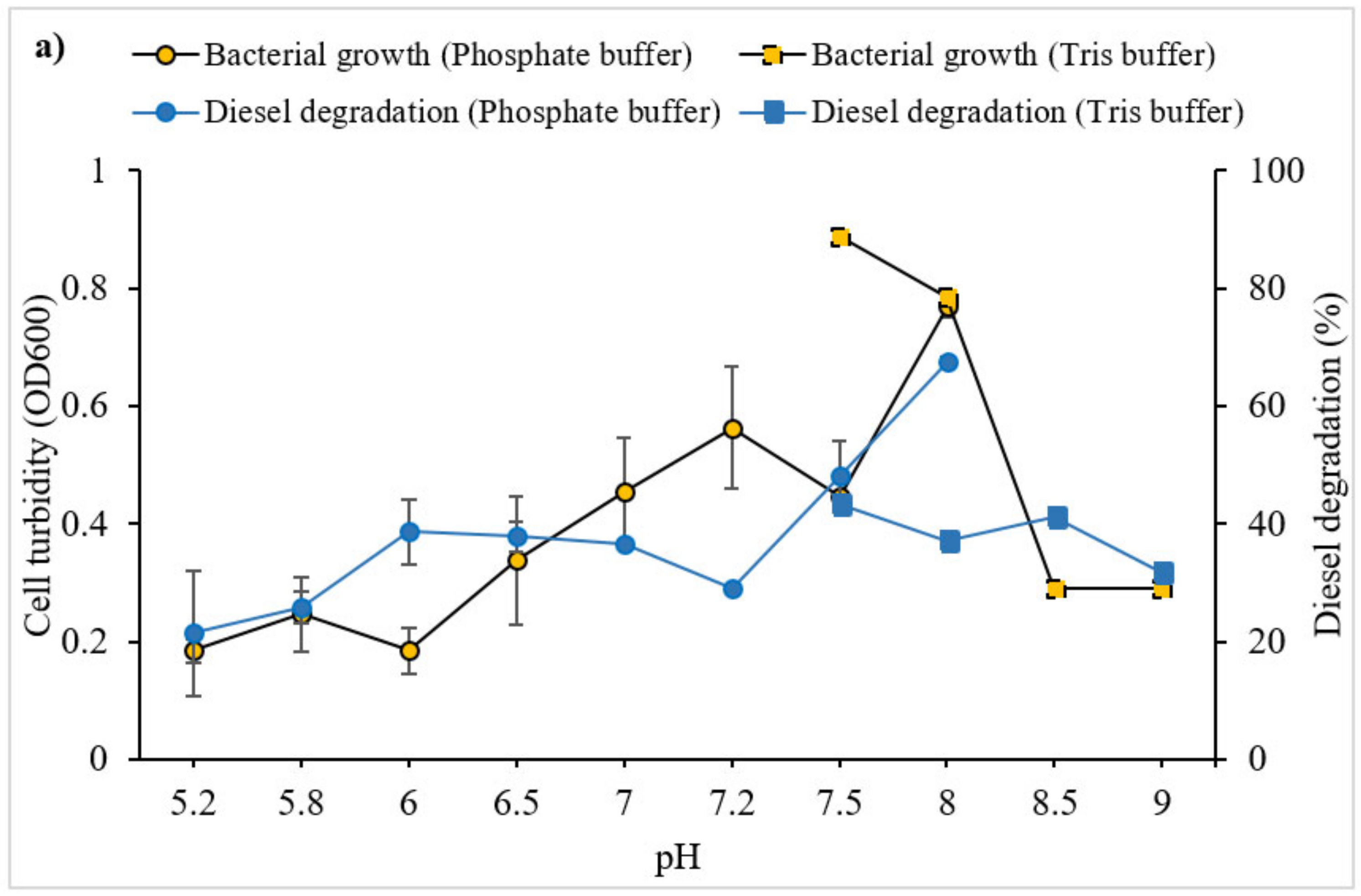
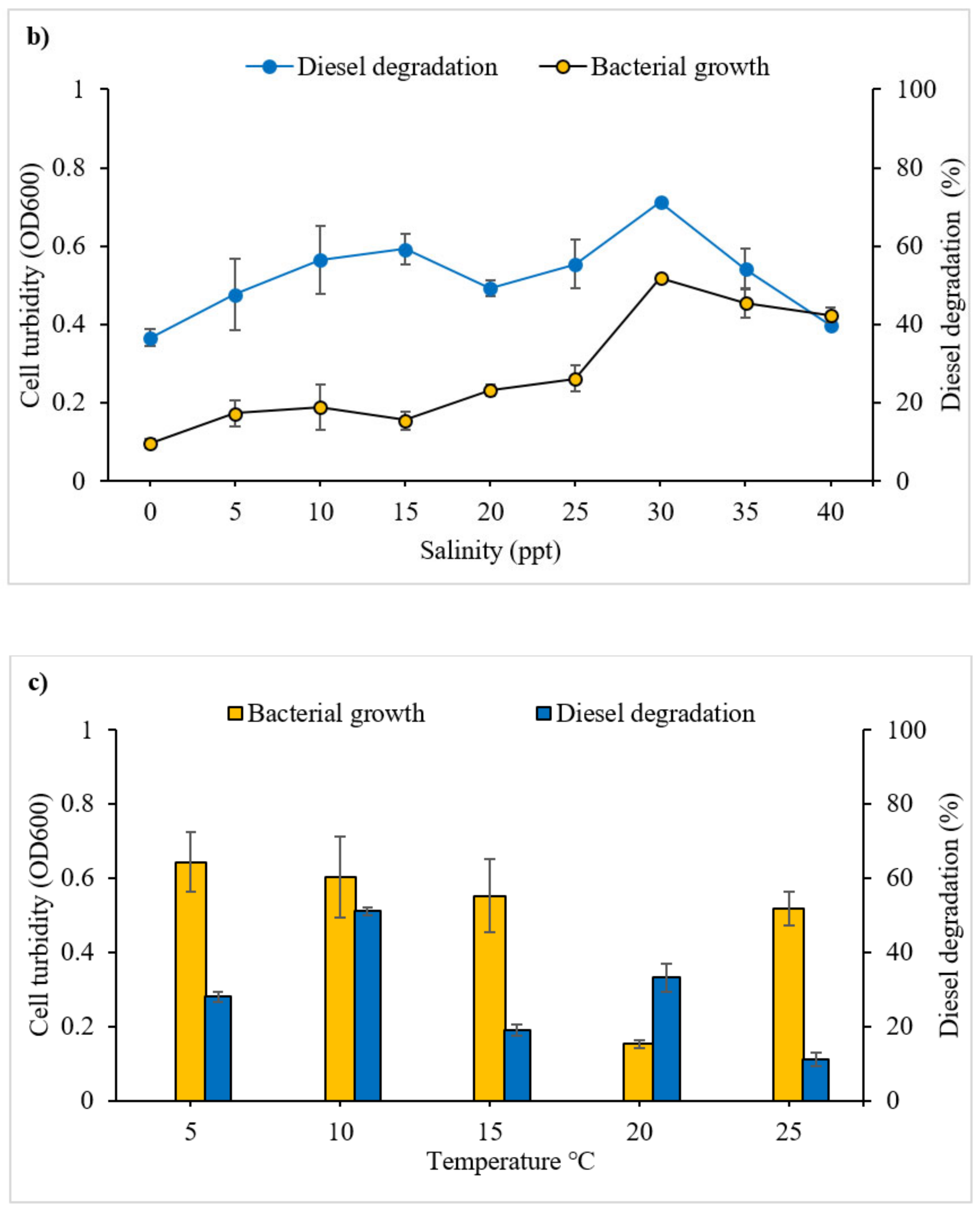
3.2.1. Effect of pH, Salinity and Temperature on Diesel Degradation
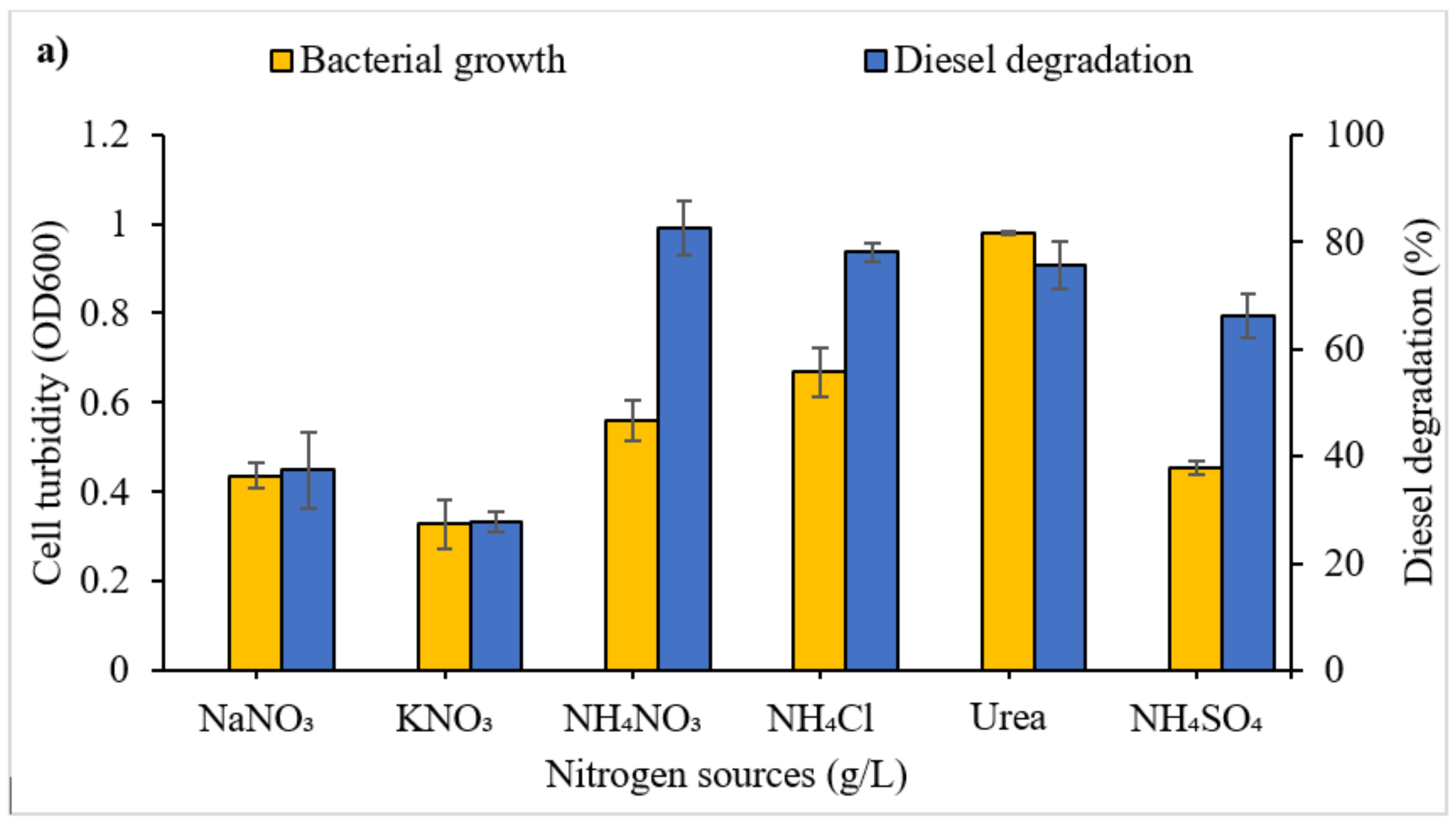
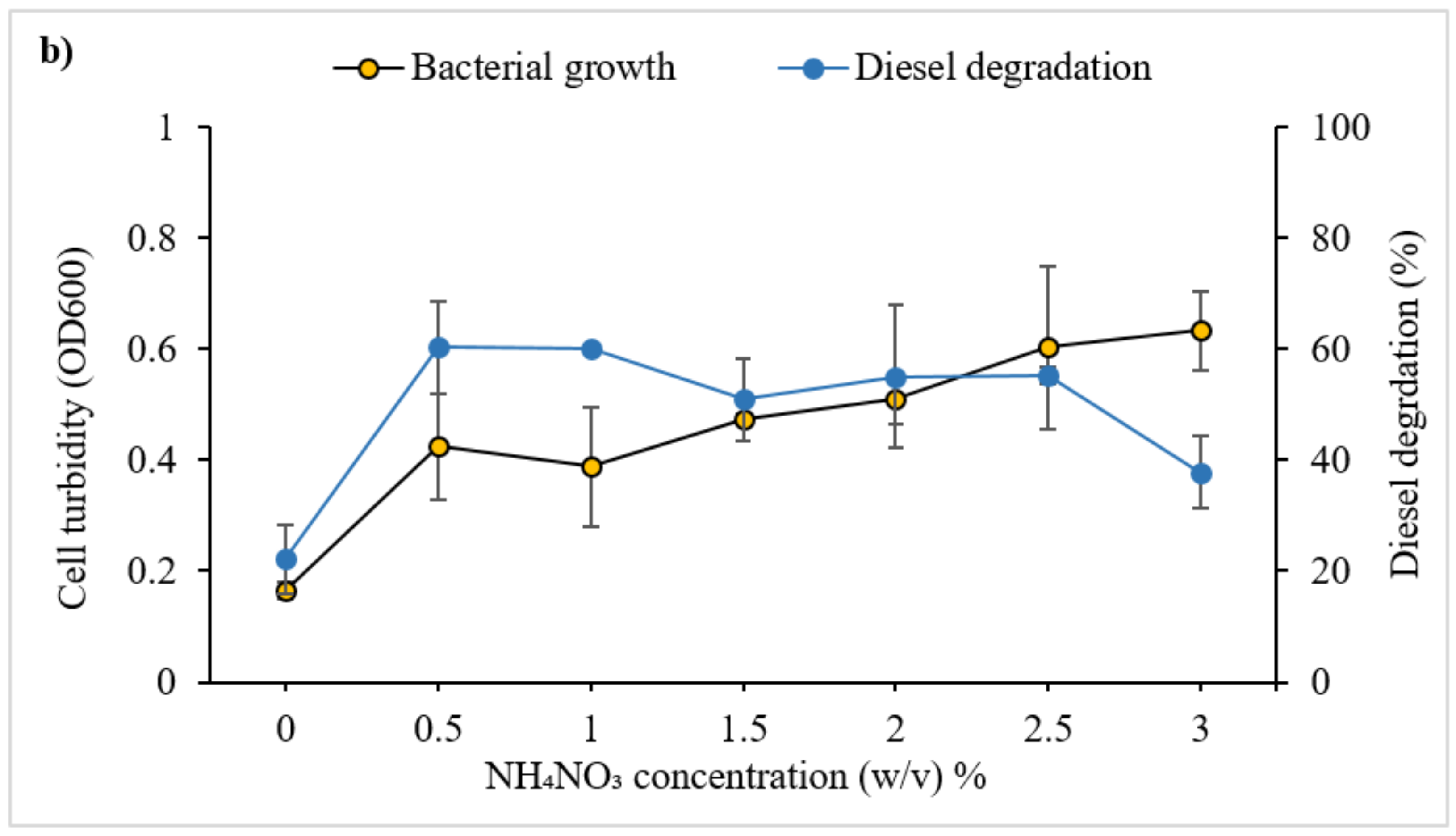
3.2.2. Effect of Nitrogen Source and Concentration on Diesel Degradation
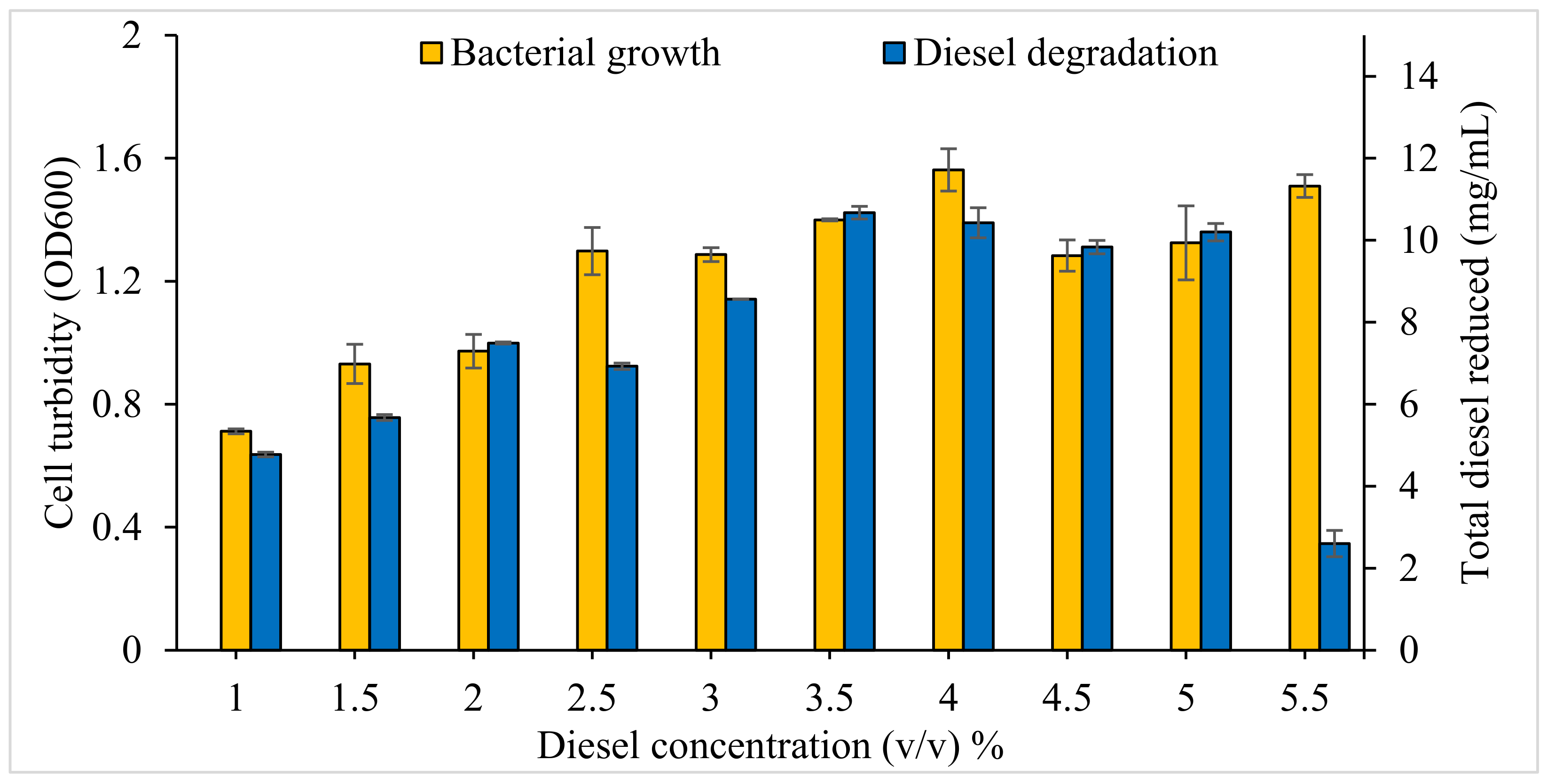
3.2.3. Effect of Initial Diesel Concentration on Bacterial Consortium Growth and Diesel Degradation
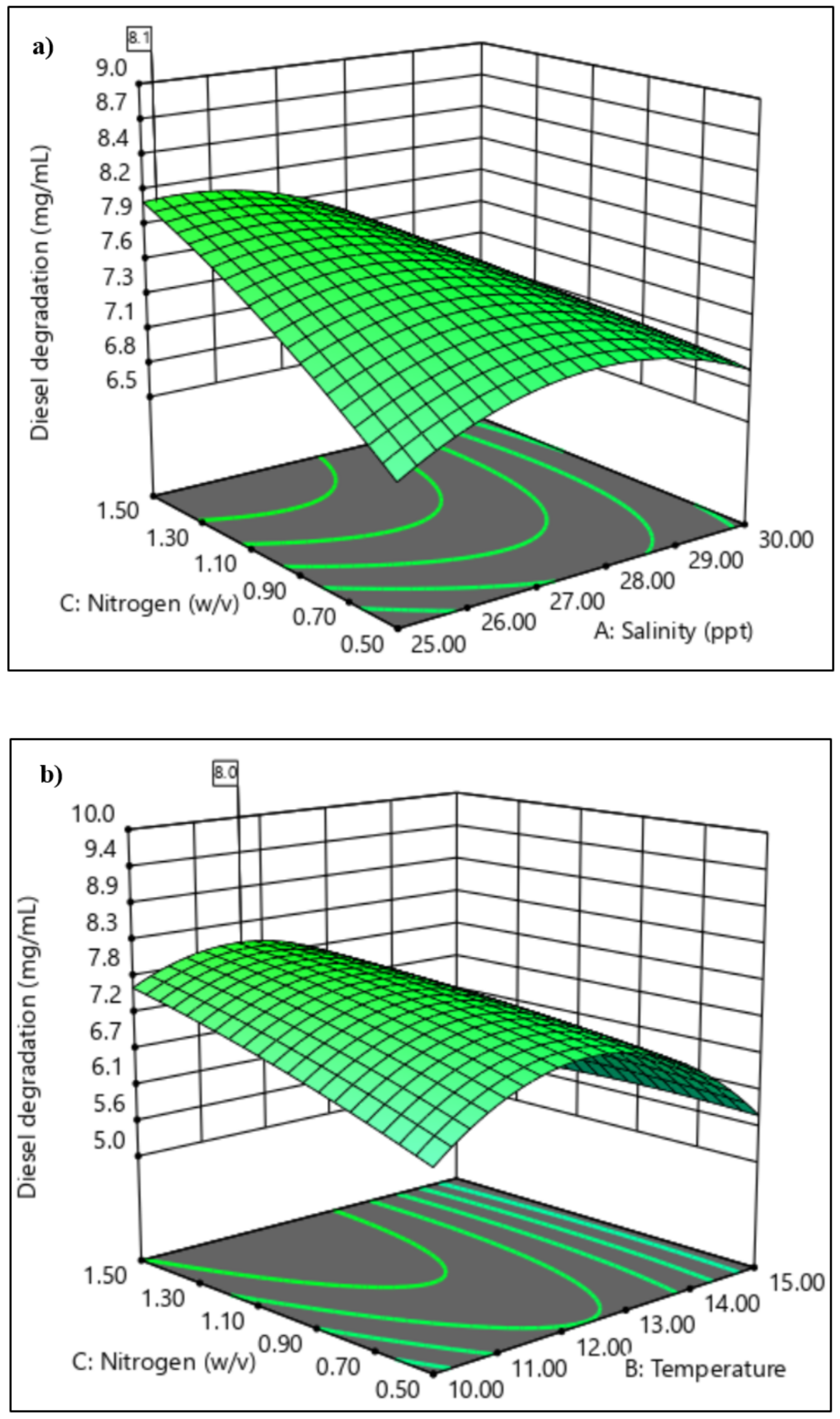
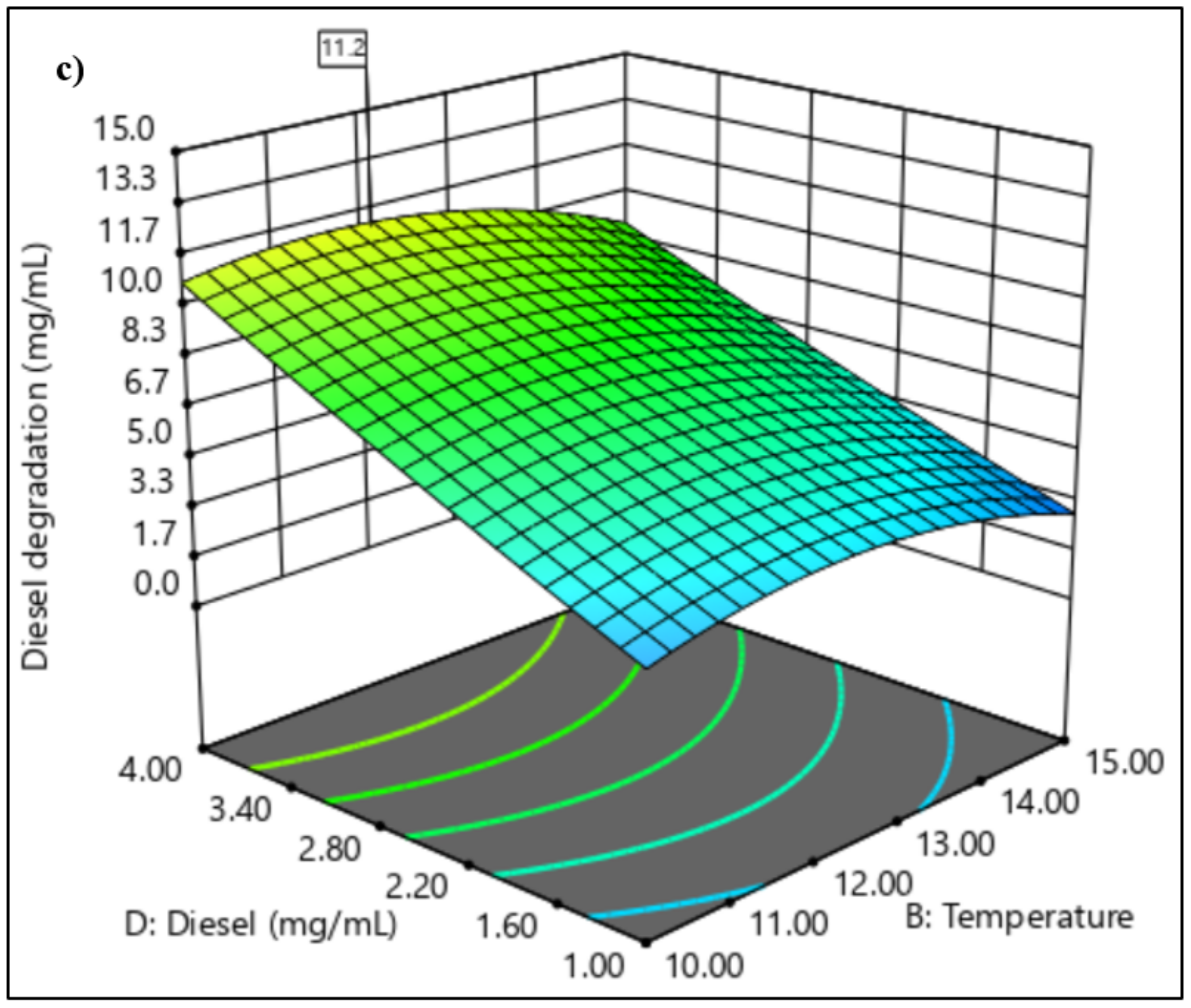
3.3. Application of Response Surface Methodology in the Selection of Significant Variables
3.3.1. Plackett-Burman Design
3.3.2. Central Composite Design
4. Discussion
4.1. Optimisation of Growth Conditions Using Conventional One-Factor-at-a-Time
4.1.1. pH
4.1.2. Salinity
4.1.3. Temperature
4.1.4. Nitrogen: Source and Concentration
4.1.5. Diesel Concentration
4.2. Application of Response Surface Methodology in the Selection of Significant Variables
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martins, C.C.; Bicego, M.C.; Taniguchi, S.; Montone, R.C. Aliphatic and polycyclic aromatic hydrocarbons in surface sediments in Admiralty Bay, King George Island, Antarctica. Antarct. Sci. 2004, 16, 117–122. [Google Scholar] [CrossRef]
- Cripps, G.C.; Shears, J. The fate in the marine environment of a minor diesel fuel spill from an Antarctic research station. Environ. Monit. Assess. 1997, 46, 221–232. [Google Scholar] [CrossRef]
- Stewart, E.J.; Draper, D. The sinking of the MS explorer: Implications for cruise tourism in Arctic Canada. Arctic 2008, 61, 224. [Google Scholar] [CrossRef] [Green Version]
- Stark, J.S.; Mohammad, M.; McMinn, A.; Ingels, J. The effects of hydrocarbons on meiofauna in marine sediments in Antarctica. J. Exp. Mar. Biol. Ecol. 2017, 496, 56–73. [Google Scholar] [CrossRef]
- Khalid, F.E.; Lim, Z.S.; Sabri, S.; Gomez-Fuentes, C.; Zulkharnain, A.; Ahmad, S.A. Bioremediation of diesel contaminated marine water by bacteria: A review and bibliometric analysis. J. Mar. Sci. Eng. 2021, 9, 155. [Google Scholar] [CrossRef]
- Hull, B.; Bergstrom, D. Antarctic terrestrial and limnetic ecosystem conservation and management. In Trends in Antarctic Terrestrial and Limnetic Ecosystems; Bergstrom, D.M., Convey, P., Huiskes, A.H.L., Eds.; Springer: Cham, Switzerland, 2006; pp. 317–340. [Google Scholar] [CrossRef]
- Kennicutt, M.C.; Sweet, S.T.; Fraser, W.R.; Stockton, W.L.; Culver, M. The Fate of Diesel Fuel Spilled by the Bahia Paraiso in Arthur Harbor, Antarctica. In Proceedings of the 1991 International Oil Spill Conference, San Diego, California, USA, 4–7 March 1991; pp. 493–500. [Google Scholar] [CrossRef]
- Wilkness, P. Fuel spill clean up in the Antarctic. Antarct. J. USA 1990, 25, 3–8. [Google Scholar]
- Hamanaka, R.; Obara, S. Study on the Fuel Consumption in the Antarctica Showa Base Microgrid. In Proceedings of the IEEE PES Asia-Pacific Power and Energy Engineering Conference (APPEEC), Xian, China, 25–28 October 2016. [Google Scholar] [CrossRef]
- Brooks, S.T.; Jabour, J.; Sharman, A.J.; Bergstrom, D.M. An analysis of environmental incidents for a national Antarctic program. J. Environ. Manag. 2018, 212, 340–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, Y.; Zuo, G.; Chang, X.; Chen, Y. A study of a standalone renewable energy system of the Chinese Zhongshan Station in Antarctica. Appl. Sci. 2019, 9, 1968. [Google Scholar] [CrossRef] [Green Version]
- Aislabie, J.M.; Balks, M.R.; Astori, N.; Stevenson, G.; Symons, R. Polycyclic aromatic hydrocarbons in fuel-oil contaminated soils, Antarctica. Chemosphere 1999, 39, 2201–2207. [Google Scholar] [CrossRef]
- Bargagli, R. Environmental contamination in Antarctic ecosystems. Sci. Total Environ. 2008, 400, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Cabrerizo, A.; Tejedo, P.; Dachs, J.; Benayas, J. Anthropogenic and biogenic hydrocarbons in soils and vegetation from the South Shetland Islands (Antarctica). Sci. Total Environ. 2016, 569, 1500–1509. [Google Scholar] [CrossRef]
- Wong, R.R.; Lim, Z.S.; Shaharuddin, N.A.; Zulkharnain, A.; Gomez-Fuentes, C.; Ahmad, S.A. Diesel in Antarctica and a bibliometric study on its indigenous microorganisms as remediation agent. Int. J. Environ. Res. Public Health 2021, 18, 1512. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, N.N.; Convey, P.; Gomez-Fuentes, C.; Zulkharnain, A.; Sabri, S.; Shaharuddin, N.A.; Ahmad, S.A. Oil bioremediation in the marine environment of Antarctica: A review and bibliometric keyword cluster analysis. Microorganisms 2021, 9, 419. [Google Scholar] [CrossRef]
- Tin, T.; Fleming, Z.L.; Hughes, K.A.; Ainley, D.G.; Convey, P.; Moreno, C.A.; Pfeiffer, S.; Scott, J.; Snape, I. Impacts of local human activities on the Antarctic environment. Antarct. Sci. 2009, 21, 3–33. [Google Scholar] [CrossRef] [Green Version]
- Fuoco, R.; Giannarelli, S.; Wei, Y.; Abete, C.; Francesconi, S.; Termine, M. Polychlorobiphenyls and polycyclic aromatic hydrocarbons in the sea-surface micro-layer and the water column at Gerlache Inlet, Antarctica. J. Environ. Monit. 2005, 7, 1313–1319. [Google Scholar] [CrossRef]
- Stortini, A.M.; Martellini, T.; Del Bubba, M.; Lepri, L.; Capodaglio, G.; Cincinelli, A. n-Alkanes, PAHs and surfactants in the sea surface microlayer and sea water samples of the Gerlache Inlet Sea (Antarctica). Microchem. J. 2009, 92, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, T. Occurrence and roles of the obligate hydrocarbonoclastic bacteria in the ocean when there is no obvious hydrocarbon contamination. In Taxonomy, Genomics and Ecophysiology of Hydrocarbon-degrading Microbes (Handbook of Hydrocarbon and Lipid Microbiology); McGenity, T.J., Ed.; Springer: Cham, Switzerland, 2018; pp. 1–17. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Casella, P.; Caruso, C.; Mangano, S.; Bruni, V.; De Domenico, M.; Michaud, L. Occurrence and characterization of psychrotolerant hydrocarbon-oxidizing bacteria from surface seawater along the Victoria Land coast (Antarctica). Polar Biol. 2010, 33, 929–943. [Google Scholar] [CrossRef]
- Gentile, G.; Bonsignore, M.; Santisi, S.; Catalfamo, M.; Giuliano, L.; Genovese, L.; Yakimov, M.M.; Denaro, R.; Genovese, M.; Cappello, S. Biodegradation potentiality of psychrophilic bacterial strain Oleispira antarctica RB-8T. Mar. Pollut. Bull. 2016, 105, 125–130. [Google Scholar] [CrossRef]
- Radwan, S.S.; Khanafer, M.M.; Al-Awadhi, H.A. Ability of the so-called obligate hydrocarbonoclastic bacteria to utilize nonhydrocarbon substrates thus enhancing their activities despite their misleading name. BMC Microbiol. 2019, 19, 41. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M.; Hazen, T.C. Oil biodegradation and bioremediation: A tale of the two worst spills in US history. Environ. Sci. Technol. 2011, 45, 6709–6715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, C.; Iglesias, K.; Bicego, M.; Taniguchi, S.; Sasaki, S.T.; Kandratavicius, N.; Bueno, C.; Brugnoli, E.; Venturini, N. Hydrocarbons in soil and meltwater stream sediments near Artigas Antarctic Research Station: Origin, sources and levels. Antarct. Sci. 2018, 30, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, C.P.D.; Peperzak, L.; Beggah, S.; Wick, L.Y.; Wuerz, B.; Weber, J.; Arey, J.S.; van der Burg, B.; Jonas, A.; Huisman, J.; et al. Immediate ecotoxicological effects of short-lived oil spills on marine biota. Nat. Commun. 2016, 4, 11206. [Google Scholar] [CrossRef] [PubMed]
- Mandri, T.; Lin, J. Isolation and characterization of engine oil degrading indigenous microorganisms in Kwazulu-Natal, South Africa. Afr. J. Biotechnol. 2007, 6, 23–27. [Google Scholar] [CrossRef]
- Prince, R.C. Bioremediation of marine oil spills. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Chapter 16; pp. 2618–2626. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.B.; Oliveira, J.S.; Silva-Portela, R.C.; Araujo, W.; Carlos, A.C.; Vasconcelos, A.T.R.; Freitas, A.T.; Domingos, Y.S.; de Farias, M.F.; Fernandes, G.J.T.; et al. Metagenome enrichment approach used for selection of oil-degrading bacteria consortia for drill cutting residue bioremediation. Environ. Pollut. 2018, 235, 869–880. [Google Scholar] [CrossRef]
- Margesin, R.; Moertelmaier, C.; Mair, J. Low-temperature biodegradation of petroleum hydrocarbons (n-alkanes, phenol, anthracene, pyrene) by four actinobacterial strains. Int. Biodeterior. Biodegrad. 2013, 84, 185–191. [Google Scholar] [CrossRef]
- Kim, S.J.; Kweon, O.; Sutherland, J.B.; Kim, H.L.; Jones, R.C.; Burback, B.L.; Graves, B.L.; Psurny, E.; Cerniglia, C.E. Dynamic response of Mycobacterium vanbaalenii PYR-1 to BP Deepwater Horizon crude oil. Appl. Environ. Microbiol. 2015, 81, 4263–4276. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.H.; Zhang, B.Y.; Chen, B.; Zhu, Z.W.; Lin, W.Y.; Cao, T. Screening of biosurfactant producers from petroleum hydrocarbon contaminated sources in cold marine environments. Mar. Pollut. Bull. 2014, 86, 402–410. [Google Scholar] [CrossRef]
- De Domenico, M.; Lo Giudice, A.; Michaud, L.; Saitta, M.; Bruni, V. Diesel oil and PCB-degrading psychrotrophic bacteria isolated from Antarctic seawaters (Terra Nova Bay, Ross Sea). Polar Res. 2004, 23, 141–146. [Google Scholar] [CrossRef]
- Gurav, R.; Lyu, H.; Ma, J.; Tang, J.; Liu, Q.; Zhang, H. Degradation of n-alkanes and PAHs from the heavy crude oil using salt-tolerant bacterial consortia and analysis of their catabolic genes. Environ. Sci. Pollut. Res. 2017, 24, 11392–11403. [Google Scholar] [CrossRef]
- Crisafi, F.; Giuliano, L.; Yakimov, M.M.; Azzaro, M.; Denaro, R. Isolation and degradation potential of a cold-adapted oil/PAH-degrading marine bacterial consortium from Kongsfjorden (Arctic region). Rend. Lincei 2016, 27 (Suppl. S1), 261–270. [Google Scholar] [CrossRef]
- Powell, S.; Stark, J.; Snape, I.; Woolfenden, E.; Bowman, J.; Riddle, M. Effects of diesel and lubricant oils on Antarctic benthic microbial communities over five years. Aquat. Microb. Ecol. 2010, 59, 119–127. [Google Scholar] [CrossRef]
- Rizzo, C.C.; Malavenda, R.; Gerçe, B.; Papale, M.; Syldatk, C.; Hausmann, R.; Bruni, V.; Michaud, L.; Lo Giudice, A.; Amalfitano, S. Effects of a simulated acute oil spillage on bacterial communities from Arctic and Antarctic marine sediments. Microorganisms 2019, 7, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, K.E.; King, C.K.; Harrison, P.L. Impacts of petroleum fuels on fertilization and development of the Antarctic sea urchin Sterechinus neumayeri. Environ. Toxicol. Chem. 2020, 39, 2527–2539. [Google Scholar] [CrossRef] [PubMed]
- Dvořák, P.; Nikel, P.I.; Damborský, J.; de Lorenzo, V. Bioremediation 3.0: Engineering pollutant-removing bacteria in the times of systemic biology. Biotechnol. Adv. 2017, 35, 845–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jesus, H.E.; Peixoto, R.S.; Rosado, A.S. Bioremediation in Antarctic Soils. J. Pet. Environ. Biotechnol. 2015, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Roslee, A.; Zakaria, N.N.; Convey, P.; Zulkharnain, A.; Lee, G.; Gomez-Fuentes, C.; Ahmad, S.A. Statistical optimisation of growth conditions and diesel degradation by the Antarctic bacterium, Rhodococcus sp. strain AQ5‒07. Extremophiles 2020, 24, 277–291. [Google Scholar] [CrossRef]
- Yang, S.Z.; Wen, X.; Shi, Y.L.; Liebner, S.; Jin, H.J.; Perfumo, A. Hydrocarbon degraders establish at the costs of microbial richness, abundance and keystone taxa after crude oil contamination in permafrost environments. Sci. Rep. 2016, 6, 37473. [Google Scholar] [CrossRef] [PubMed]
- Tengku-Mazuki, T.; Subramaniam, K.; Zakaria, N.; Convey, P.; Abdul Khalil, K.; Lee, G.; Zulkharnain, A.; Shaharuddin, N.A.; Ahmad, S. Optimization of phenol degradation by Antarctic bacterium Rhodococcus sp. Antarct. Sci. 2020, 32, 486–495. [Google Scholar] [CrossRef]
- Bushnell, L.D.; Haas, H.F. The utilization of certain hydrocarbons by microorganisms. J. Bacteriol. 1941, 41, 653–673. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.J.; Correa, P.F.; Almeidaa, D.G.; Lunaa, J.M.; Rufino, R.D.; Sarubbo, L.A. Recovery of contaminated marine environments by biosurfactant-enhanced bioremediation. Colloids Surf. B Biointerfaces 2018, 172, 127–135. [Google Scholar] [CrossRef]
- Chen, Q.; Bao, M.; Fan, X.; Liang, S.; Sun, P. Rhamnolipids enhance marine oil spill bioremediation in laboratory system. Mar. Pollut. Bull. 2013, 71, 269–275. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, K.M.; Perkins, M.J.; Field, J.A.; Leigh, M.B. Biodegradation of crude oil and corexit 9500 in Arctic seawater. Front. Microbiol. 2018, 9, 1788. [Google Scholar] [CrossRef] [Green Version]
- Plackett, R.L.; Burman, J.P. The design of optimum multifactorial experiments. Biometals 1946, 33, 305–325. [Google Scholar] [CrossRef]
- Xiao, Z.J.; Liu, P.H.; Qin, J.Y.; Xu, P. Statistical optimization of medium components for enhanced acetoin production from molasses and soybean meal hydrolyzate. Appl. Microbiol. Biotechnol. 2007, 74, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L.C. Saprophytic, symbiotic and parasitic bacteria: Importance to environment, biotechnological applications and biocontrol. Adv. Res. 2014, 2, 250–265. [Google Scholar] [CrossRef]
- Wright, A.L.; Weaver, R.W.; Webb, J.W. Oil bioremediation in salt marsh mesocosms as influences by N and P fertilisation, flooding and season. Water Air Soil Pollut. 1997, 95, 179–191. [Google Scholar] [CrossRef]
- Delille, D.; Delille, B.; Pelletier, E. Effectiveness of bioremediation of crude oil contaminated subantarctic intertidal sediment: The microbial response. Microb. Ecol. 2002, 44, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Sherry, N.D.; Berges, J.A.; Bishop, J.K.B.; Calvert, S.E.; Charette, M.A.; Giovannoni, S.J.; Goldblatt, R.; Harrison, P.J.; Moran, S.B.; et al. Transformations of biogenic particulates from the pelagic to the deep ocean realm. Deep Sea Res. Part II 1999, 46, 2761–2792. [Google Scholar] [CrossRef]
- Das, S.; Mangwani, N. Ocean acidification and marine microorganisms: Responses and consequences. Oceanologia 2015, 57, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Atlas, R.M. Fate of petroleum pollutants in Arctic ecosystems. Water Sci. Technol. 1986, 18, 59–67. [Google Scholar] [CrossRef]
- Pereira, E.; Napp, A.; Allebrandt, S.; Barbosa, R.; Reuwsaat, J.; Lopes, W.; Kmetzsch, L.; Staats, C.; Schrank, A.; Dallegrave, A.; et al. Biodegradation of aliphatic and polycyclic aromatic hydrocarbons in seawater by autochthonous microorganisms. Int. Biodeterior. Biodegrad. 2019, 145, 104789. [Google Scholar] [CrossRef]
- Rivaro, P.; Messa, R.; Ianni, C.; Magi, E.; Budillon, G. Distribution of total alkalinity and pH in the Ross Sea (Antarctica) waters during austral summer 2008. Polar Res. 2014, 33, 20403. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Sutherland, S.C.; Chipman, D.W.; Goddard, J.G.; Ho, C.; Newberger, T.; Sweeney, C.; Munro, D.R. Climatological distributions of pH, pCO2, total CO2, alkalinity, and CaCO3 saturation in the global surface ocean, and temporal changes at selected locations. Mar. Chem. 2014, 164, 95–125. [Google Scholar] [CrossRef] [Green Version]
- Jury, C.P.; Thomas, F.I.M.; Atkinson, M.J.; Toonen, R.J. Buffer capacity, ecosystem feedbacks, and seawater chemistry under global change. Water 2013, 5, 1303–1325. [Google Scholar] [CrossRef]
- Cai, W.-J.; Huang, W.-J.; Luther, G.W.; Pierrot, D.; Li, M.; Testa, J.; Xue, M.; Joesoef, A.; Mann, R.; Brodeur, J.; et al. Redox reactions and weak buffering capacity lead to acidification in the Chesapeake Bay. Nat. Commun. 2017, 369, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Koolivand, A.; Rajaei, M.S.; Ghanadzadeh, M.J.; Saeedi, R.; Abtahi, H.; Godini, K. Bioremediation of storage tank bottom sludge by using a two-stage composting system: Effect of mixing ratio and nutrients addition. Bioresour. Technol. 2017, 235, 240–249. [Google Scholar] [CrossRef]
- Harvey, H.W. The Chemistry and Fertility of Sea Waters; Cambridge University Press: London, UK, 1957; p. 248. [Google Scholar]
- Rozen, Y.; Belkin, S. Survival of enteric bacteria in seawater. FEMS Microbiol. Rev. 2001, 25, 513–529. [Google Scholar] [CrossRef]
- Booth, I.R. Regulation of cytoplasmic pH in bacteria. Microbiol. Rev. 1985, 49, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T.A. Alkaline pH homeostasis in bacteria: New insights. Biochim. Biophys. Acta Biomembr. 2005, 1717, 67–88. [Google Scholar] [CrossRef] [Green Version]
- Borah, D.; Yadav, R.N.S. Optimization of BH medium for efficient biodegradation of diesel, crude oil and used engine oil by a newly isolates B. cereus strain DRDU1 from an automobile engine. Biotechnology 2014, 13, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Strynar, M.; Sen, D.; Weaver, R.W. Nitrogen and phosphorus for growth of oil-degrading microorganisms in seawater. Bioremediat. J. 1999, 3, 85–91. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Giuliano, L.; Gentile, G.; Crisafi, E.; Chernikova, T.N.; Abraham, W.R.; Lünsdorf, H.; Timmis, K.N.; Golyshin, P.N. Oleispira antarctica gen. nov., sp. nov., a novel hydrocarbonoclastic marine bacterium isolated from Antarctic coastal sea water. Int. J. Syst. Evol. Microbiol. 2003, 53, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Bers, A.V.; Momo, F.; Schloss, I.R.; Abele, D. Analysis of trends and sudden changes in long-term environmental data from King George Island (Antarctica): Relationships between global climatic oscillations and local system response. Clim. Chang. 2013, 116, 789–803. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, D.; Marshall, G.; Connolley, W.; Parkinson, C.; Mulvaney, R.; Hodgson, D.; King, J.; Pudsey, C.; Turner, J. Recent rapid regional climate warming on the Antarctic Peninsula. Clim. Chang. 2003, 60, 243–274. [Google Scholar] [CrossRef]
- Turner, J.; Overland, J. Contrasting climate change in the two polar regions. Polar Res. 2009, 28, 146–164. [Google Scholar] [CrossRef]
- Choudhry, I. Antarctic Treaty Consultative Meeting (ATCM) XLII/Committee for Environmental Protection (CEP) XXII. Polar J. 2019, 9, 476–477. [Google Scholar] [CrossRef]
- Turner, J.; Barrand, N.E.; Bracegirdle, T.J.; Convey, P.; Hodgson, D.A.; Jarvis, M.; Jenkins, A.; Marshall, G.; Meredith, M.P.; Roscoe, H.; et al. Antarctic climate change and the environment: An update. Polar Rec. 2014, 50, 237–259. [Google Scholar] [CrossRef] [Green Version]
- Convey, P.; Peck, L.S. Antarctic environmental change and biological responses. Sci. Adv. 2019, 5, eaaz0888. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.; Murphy, E.J.; Meredith, M.P.; King, J.C.; Peck, L.S.; Barnes, D.K.; Smith, R.C. Climate change and the marine ecosystem of the western Antarctic Peninsula. Philos. Trans. R. Soc. B 2007, 362, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Blanco, A.; Duval, A.; Pelletier, E.; Delille, D.; Ghiglione, J.F. Effects of temperature and fertilization on the structure of total versus active bacterial communities from sub-Antarctic seawater exposed to crude oil and diesel fuel. Polar Res. 2013, 32, 18521. [Google Scholar] [CrossRef] [Green Version]
- Nedwell, D.B. Effect of low temperature on microbial growth: Lowered affinity for substrates limits growth at low temperature. FEMS Microbiol. Ecol. 1999, 30, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Leahy, J.G.; Colwell, R.R. Microbial degradation of hydrocarbons in the environment. Microbiol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Speight, J.G. Industrial inorganic chemistry. In Environmental Inorganic Chemistry for Engineers; Speight, J.G., Ed.; CD&W Inc.: Laramie, WY, USA, 2017; Chapter 3; pp. 111–169. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Morán, X.A.; Ducklow, H. Microbial growth in the polar oceans—Role of temperature and potential impact of climate change. Nat. Rev. Microbiol. 2009, 7, 451–459. [Google Scholar] [CrossRef]
- Head, I.M.; Jones, D.M.; Röling, W.F. Marine microorganisms make a meal of oil. Nat. Rev. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef]
- Marietou, A.; Chastain, R.; Beulig, F.; Scoma, A.; Hazen, T.; Bartlett, D. The effect of hydrostatic pressure on enrichments of hydrocarbon degrading microbes from the Gulf of Mexico following the Deepwater Horizon oil spill. Front. Microbiol. 2018, 9, 808. [Google Scholar] [CrossRef]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L.; et al. Deep-sea oil plume enriches indigenous oil-degrading bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Valentine, D.L.; Kessler, J.D.; Redmond, M.C.; Mendes, S.D.; Heintz, M.B.; Farwell, C.; Hu, L.; Kinnaman, F.S.; Yvon-Lewis, S.; Du, M.; et al. Propane respiration jump-starts microbial response to a deep oil spill. Science 2010, 330, 208–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubinsky, E.A.; Conrad, M.E.; Chakraborty, R.; Bill, M.; Borglin, S.E.; Hollibaugh, J.T.; Mason, O.U.; Piceno, Y.M.; Reid, F.C.; Stringfellow, W.T.; et al. Succession of hydrocarbon-degrading bacteria in the aftermath of the Deepwater Horizon oil spill in the Gulf of Mexico. Environ. Sci. Technol. 2013, 47, 10860–10867. [Google Scholar] [CrossRef]
- Kwapisz, E.; Wszelaka, J.; Marchut-Mikolajczyk, O.; Bielecki, S. The effect of nitrate and ammonium ions on kinetics of diesel oil degradation by Gordonia alkanivorans S7. Int. Biodeterior. Biodegrad. 2008, 61, 214–222. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Bordoloi, N.K. Biodegradation of benzene, toluene and xylene (BTX) in liquid culture and in soil by Bacillus subtilis and Pseudomonas aeruginosa strains and a formulated bacterial consortium. Environ. Sci. Pollut. Res. Int. 2012, 19, 3380–3388. [Google Scholar] [CrossRef]
- Ferguson, S. Nitrogen cycle enzymology. Curr. Opin. Chem. Biol. 1998, 2, 182–193. [Google Scholar] [CrossRef]
- Carswell, A.; Shaw, R.; Hunt, J.; Sánchez-Rodríguez, A.R.; Saunders, K.; Cotton, J.; Hill, P.; Chadwick, D.R.; Jones, D.L.; Misselbrook, T.H. Assessing the benefits and wider costs of different N fertilisers for grassland agriculture. Arch. Agron. Soil Sci. 2019, 65, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Urakawa, H.; Rajan, S.; Feeney, M.E.; Sobecky, P.A.; Mortazavi, B. Ecological response of nitrification to oil spills and its impact on the nitrogen cycle. Environ. Microbiol. 2019, 21, 18–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, R. Nitrogen cycling in coastal marine ecosystems. FEMS Microbiol. Rev. 1999, 23, 563–590. [Google Scholar] [CrossRef] [PubMed]
- Isnansetyo, A.; Getsu, S.; Seguchi, M.; Koriyama, M. Independent effects of temperature, salinity, ammonium concentration and pH on nitrification rate of the Ariake seawater above mud sediment. HAYATI J. Biosci. 2014, 21, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Vyas, T.K.; Dave, B.P. Effect of nitrogen, phosphorous and potassium fertiliser on biodegradation of crude oil by marine bacteria. Indian J. Mar. Sci. 2010, 39, 143–150. [Google Scholar]
- Chen, Y.; Yu, B.; Lin, J.; Naidu, R.; Chen, Z. Simultaneous adsorption and biodegradation (SAB) of diesel oil using immobilized Acinetobacter venetianus on porous material. Chem. Eng. J. 2016, 289, 463–470. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Wiebe, W. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol. 2001, 23, 187–204. [Google Scholar] [CrossRef] [Green Version]
- Goud, B.S.; Prathibha, D. Evaluating the effect of substrate type and concentration in the biodegradation of petroleum hydrocarbons. Int. J. Agric. Environ. Biotechnol. 2017, 10, 669–674. [Google Scholar] [CrossRef]
- Vieira, G.; Magrini, M.J.; Bonugli-Santos, R.C.; Rodrigues, M.; Sette, L.D. Polycyclic aromatic hydrocarbons degradation by marine-derived basidiomycetes: Optimization of the degradation process. Braz. J. Microbiol. 2018, 49, 749–756. [Google Scholar] [CrossRef]
- Kanaly, R.A.; Harayama, S.; Watanabe, K. Rhodanobacter sp. strain BPC1 in a benzo[a]pyrene-mineralizing bacterial consortium. Appl. Environ. Microbiol. 2002, 68, 5826–5833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhu, Q.; Zeng, J.; Ding, Q.; Gong, Y.; Xing, P.; Lin, X. Effects of pH and polycyclic aromatic hydrocarbon pollution on thaumarchaeotal community in agricultural soils. J. Soil Sediment 2016, 16, 1960–1969. [Google Scholar] [CrossRef]
- Radniecki, T.S.; Dolan, M.E.; Semprini, L. Physiological and transcriptional responses of Nitrosomonas europaea to toluene and benzene inhibition. Environ. Sci. Technol. 2008, 42, 4093–4098. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Range |
|---|---|
| pH (potassium phosphate and Tris buffer) | 5.2–9.0 |
| Salinity, NaCl (ppt, w/v) | 0–50 |
| Temperature (°C) | 10–25 |
| Nitrogen source | urea, NH4NO3, NaNO3, KNO3, NH4SO4 |
| Nitrogen concentration, g/L (w/v) | 0–3.5 |
| Initial substrate concentration, diesel (%, v/v) | 0.5–5.0 |
| Variables | Code | Unit | Experimental Range | |
|---|---|---|---|---|
| Low (‒1) | High (+1) | |||
| pH | A | - | 7.5 | 8.0 |
| Salinity | B | ppt | 25.0 | 30.0 |
| Temperature | C | °C | 10.0 | 15.0 |
| Nitrogen concentration | D | g/L | 0.5 | 2.5 |
| Initial diesel concentration | E | % (v/v) | 1.0 | 4.0 |
| Symbol | Unit | Experimental Values | |||||
|---|---|---|---|---|---|---|---|
| ‒2 | ‒1 | 0 | +1 | +2 | |||
| Salinity | A | ppt | 22.5 | 25.0 | 27.5 | 30.0 | 32.5 |
| Temperature | B | °C | 7.5 | 10.0 | 12.5 | 15.0 | 17.5 |
| Nitrogen concentration | C | (g/L) | 0.0 | 0.5 | 1.0 | 1.5 | 2.0 |
| Initial diesel concentration | D | % (v/v) | 0 | 1.0 | 2.5 | 4.0 | 5.5 |
| Run | A | B | C | D | E | Degradation (mg/mL) |
|---|---|---|---|---|---|---|
| 1 | 8.0 | 30.0 | 15.0 | 0.5 | 30.51 | 9.62 ± 1.54 |
| 2 | 7.5 | 25.0 | 15.0 | 2.5 | 30.51 | 5.76 ± 2.51 |
| 3 | 8.0 | 30.0 | 10.0 | 2.5 | 30.51 | 5.17 ± 0.37 |
| 4 | 7.5 | 25.0 | 10.0 | 2.5 | 30.51 | 5.06 ± 1.06 |
| 5 | 7.5 | 30.0 | 15.0 | 2.5 | 7.69 | 2.34 ± 2.24 |
| 6 | 8.0 | 30.0 | 10.0 | 2.5 | 7.69 | 2.05 ± 1.01 |
| 7 | 8.0 | 25.0 | 15.0 | 2.5 | 7.69 | 2.10 ± 1.99 |
| 8 | 8.0 | 25.0 | 10.0 | 0.5 | 30.51 | 8.03 ± 1.98 |
| 9 | 7.5 | 30.0 | 10.0 | 0.5 | 7.69 | 2.85 ± 8.17 |
| 10 | 8.0 | 25.0 | 15.0 | 0.5 | 7.69 | 3.13 ± 1.37 |
| 11 | 7.5 | 25.0 | 10.0 | 0.5 | 7.69 | 2.89 ± 2.51 |
| 12 | 7.5 | 30.0 | 15.0 | 0.5 | 30.51 | 9.71 ± 1.02 |
| Source | Sum of Squares | DF | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Model | 629.47 | 5 | 125.89 | 164.52 | <0.0001 *** |
| A | 0.5663 | 1 | 0.56 | 0.74 | 0.4227 |
| B | 4.62 | 1 | 4.62 | 6.04 | 0.0493 * |
| C | 32.12 | 1 | 32.12 | 41.99 | 0.0006 *** |
| D | 376.17 | 1 | 376.17 | 491.59 | <0.0001 *** |
| E | 215.98 | 1 | 215.98 | 282.24 | <0.0001 *** |
| Residual | 4.59 | 6 | 0.7652 | ||
| Cor Total | 634.06 | 11 | |||
| Std. Dev. | 0.87.48 | R2 | 0.9928 | ||
| Mean | 27.92 | Adjusted R2 | 0.9867 | ||
| C.V. | 3.13 | Predicted R2 | 0.9710 | ||
| Adequate Precision | 36.4091 | ||||
| Run Order | A | B | C | D | Diesel Reduction (mg/mL) | |
|---|---|---|---|---|---|---|
| Experimental Value | Predicted Value | |||||
| 1 | 27.5 | 12.5 | 1.0 | 44.37 | 15.01 ± 3.54 | 14.84 |
| 2 | 25.0 | 15.0 | 0.5 | 7.96 | 3.15 ± 8.67 | 2.71 |
| 3 | 30.0 | 10.0 | 0.5 | 7.96 | 2.61 ± 3.10 | 2.27 |
| 4 | 25.0 | 15.0 | 1.5 | 30.51 | 8.31 ± 4.10 | 8.78 |
| 5 | 27.5 | 12.5 | 1.0 | 18.45 | 7.74 ± 17.04 | 7.63 |
| 6 | 27.5 | 17.5 | 1.0 | 18.45 | 1.63 ± 1.52 | 1.41 |
| 7 | 25.0 | 10.0 | 1.5 | 30.51 | 12.39 ± 2.37 | 11.67 |
| 8 | 25.0 | 15.0 | 0.5 | 30.51 | 7.66 ± 1.77 | 7.91 |
| 9 | 30.0 | 10.0 | 0.5 | 30.51 | 9.49 ± 5.67 | 9.67 |
| 10 | 27.5 | 12.5 | 1.0 | 0 | 0.00 | 0.00 |
| 11 | 32.5 | 12.5 | 1.0 | 18.45 | 5.83 ± 0.21 | 5.57 |
| 12 | 30.0 | 15.0 | 0.5 | 7.96 | 1.89 ± 1.28 | 2.63 |
| 13 | 30.0 | 10.0 | 1.5 | 7.96 | 2.88 ± 1.57 | 2.64 |
| 14 | 27.5 | 12.5 | 0.0 | 18.45 | 6.43 ± 9.94 | 6.59 |
| 15 | 27.5 | 12.5 | 1.0 | 18.45 | 7.88 ± 0.96 | 7.63 |
| 16 | 25.0 | 10.0 | 1.5 | 7.96 | 3.87 ± 4.40 | 4.51 |
| 17 | 25.0 | 15.0 | 1.5 | 7.96 | 3.11 ± 2.14 | 2.95 |
| 18 | 30.0 | 15.0 | 0.5 | 30.51 | 9.2 ± 2.06 | 8.72 |
| 19 | 27.5 | 12.5 | 1.0 | 18.45 | 7.72 ± 0.43 | 7.63 |
| 20 | 25.0 | 10.0 | 0.5 | 30.51 | 9.16 ± 9.50 | 9.23 |
| 21 | 30.0 | 15.0 | 1.5 | 30.51 | 8.08 ± 3.67 | 8.21 |
| 22 | 27.5 | 12.5 | 1.0 | 18.45 | 7.85 ± 0.04 | 7.63 |
| 23 | 27.5 | 12.5 | 1.0 | 18.45 | 7.09 ± 0.99 | 7.63 |
| 24 | 30.0 | 10.0 | 1.5 | 30.51 | 10.09 ± 3.33 | 10.69 |
| 25 | 27.5 | 12.5 | 2.0 | 18.45 | 8.18 ± 2.75 | 7.86 |
| 26 | 27.5 | 7.5 | 1.0 | 18.45 | 3.86 ± 1.02 | 3.91 |
| 27 | 27.5 | 12.5 | 1.0 | 18.45 | 7.48 ± 13.19 | 7.63 |
| 28 | 30.0 | 15.0 | 1.5 | 7.96 | 1.37 ± 17.36 | 1.46 |
| 29 | 22.5 | 12.5 | 1.0 | 18.45 | 6.54 ± 2.76 | 6.62 |
| 30 | 25.0 | 10.0 | 0.5 | 7.96 | 2.84 ± 6.79 | 2.73 |
| Source | Sum of | Df | Mean | F-Value | p Value | |
|---|---|---|---|---|---|---|
| Model | 307.675 | 14 | 21.977 | 85.488 | <0.0001 | *** |
| A | 1.649 | 1 | 1.649 | 6.415 | 0.0239 | * |
| B | 9.397 | 1 | 9.397 | 36.553 | <0.0001 | *** |
| C | 2.411 | 1 | 2.411 | 9.377 | 0.0084 | ** |
| D | 188.075 | 1 | 188.075 | 731.598 | <0.0001 | *** |
| AB | 0.142 | 1 | 0.142 | 0.552 | 0.469 | |
| AC | 1.987 | 1 | 1.987 | 7.728 | 0.014 | * |
| AD | 0.791 | 1 | 0.791 | 3.078 | 0.101 | |
| BC | 2.369 | 1 | 2.369 | 9.213 | 0.009 | ** |
| BD | 1.707 | 1 | 1.707 | 6.642 | 0.022 | ** |
| CD | 0.432 | 1 | 0.432 | 1.681 | 0.216 | |
| A2 | 3.907 | 1 | 3.907 | 15.197 | 0.002 | ** |
| B2 | 41.091 | 1 | 41.091 | 159.839 | <0.0001 | *** |
| C2 | 0.273 | 1 | 0.273 | 1.060 | 0.321 | |
| D2 | 0.347 | 1 | 0.347 | 1.351 | 0.265 | |
| Residual | 3.599 | 14 | 0.257 | |||
| Lack of Fit | 3.149 | 9 | 0.350 | 3.883 | 0.075 | Not significant |
| Pure Error | 0.450 | 5 | 0.090 | |||
| Cor Total | 311.274 | 28 | ||||
| R2 | 0.9894 | |||||
| Std. Dev. | 0.507 | Adjusted R2 | 0.9769 | |||
| Mean | 6.53 | Predicted R2 | 0.9326 | |||
| C.V. % | 7.76 | Adeq Precision | 38.8315 | |||
| Optimised Parameters | Value | Predicted Value | Experimental Value |
|---|---|---|---|
| pH | 8.0 | 11.66 mg/mL | 12.23 mg/mL ± 1.46 |
| Salinity (NaCl) | 25.0 ppt | ||
| Temperature | 10 °C | ||
| NH3NO4 concentration | 1.5 g/L | ||
| Initial diesel concentration | 4.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakaria, N.N.; Gomez-Fuentes, C.; Abdul Khalil, K.; Convey, P.; Roslee, A.F.A.; Zulkharnain, A.; Sabri, S.; Shaharuddin, N.A.; Cárdenas, L.; Ahmad, S.A. Statistical Optimisation of Diesel Biodegradation at Low Temperatures by an Antarctic Marine Bacterial Consortium Isolated from Non-Contaminated Seawater. Microorganisms 2021, 9, 1213. https://doi.org/10.3390/microorganisms9061213
Zakaria NN, Gomez-Fuentes C, Abdul Khalil K, Convey P, Roslee AFA, Zulkharnain A, Sabri S, Shaharuddin NA, Cárdenas L, Ahmad SA. Statistical Optimisation of Diesel Biodegradation at Low Temperatures by an Antarctic Marine Bacterial Consortium Isolated from Non-Contaminated Seawater. Microorganisms. 2021; 9(6):1213. https://doi.org/10.3390/microorganisms9061213
Chicago/Turabian StyleZakaria, Nur Nadhirah, Claudio Gomez-Fuentes, Khalilah Abdul Khalil, Peter Convey, Ahmad Fareez Ahmad Roslee, Azham Zulkharnain, Suriana Sabri, Noor Azmi Shaharuddin, Leyla Cárdenas, and Siti Aqlima Ahmad. 2021. "Statistical Optimisation of Diesel Biodegradation at Low Temperatures by an Antarctic Marine Bacterial Consortium Isolated from Non-Contaminated Seawater" Microorganisms 9, no. 6: 1213. https://doi.org/10.3390/microorganisms9061213