Mycobacteriosis and Infections with Non-tuberculous Mycobacteria in Aquatic Organisms: A Review


Abstract
:1. Introduction
2. Classification and History of the Disease
3. Distribution of the Disease
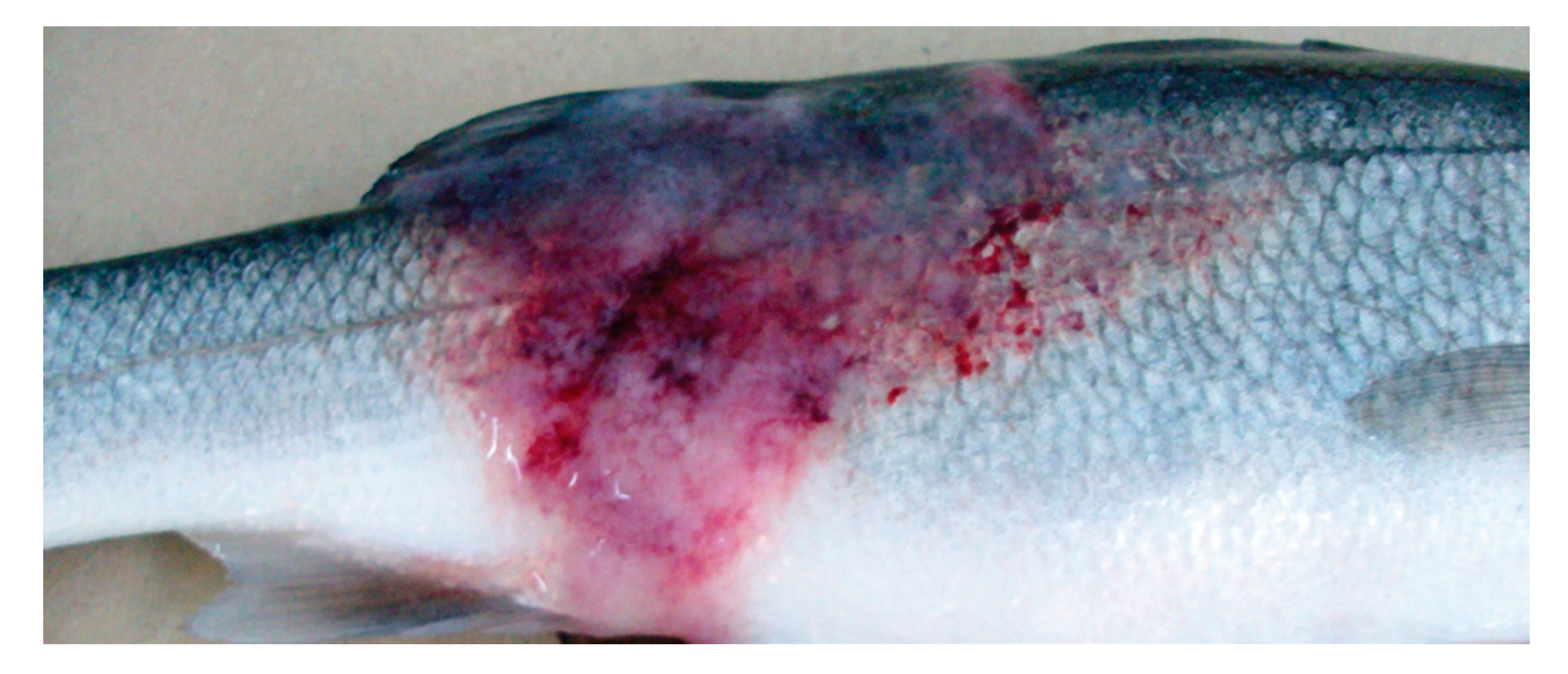
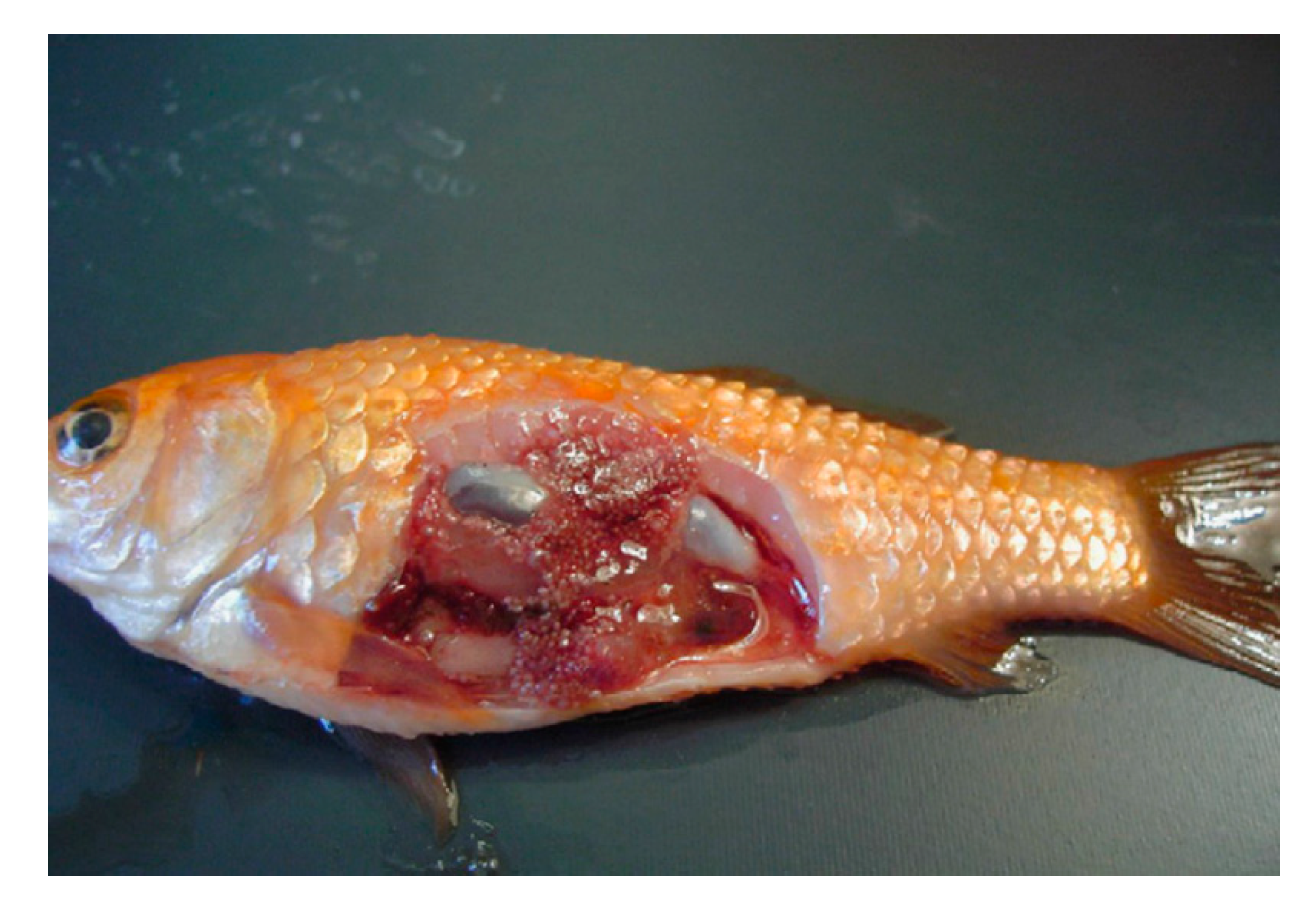
4. Course of the Disease and Clinical Signs
5. Zoonotic Consideration
6. Diagnostics
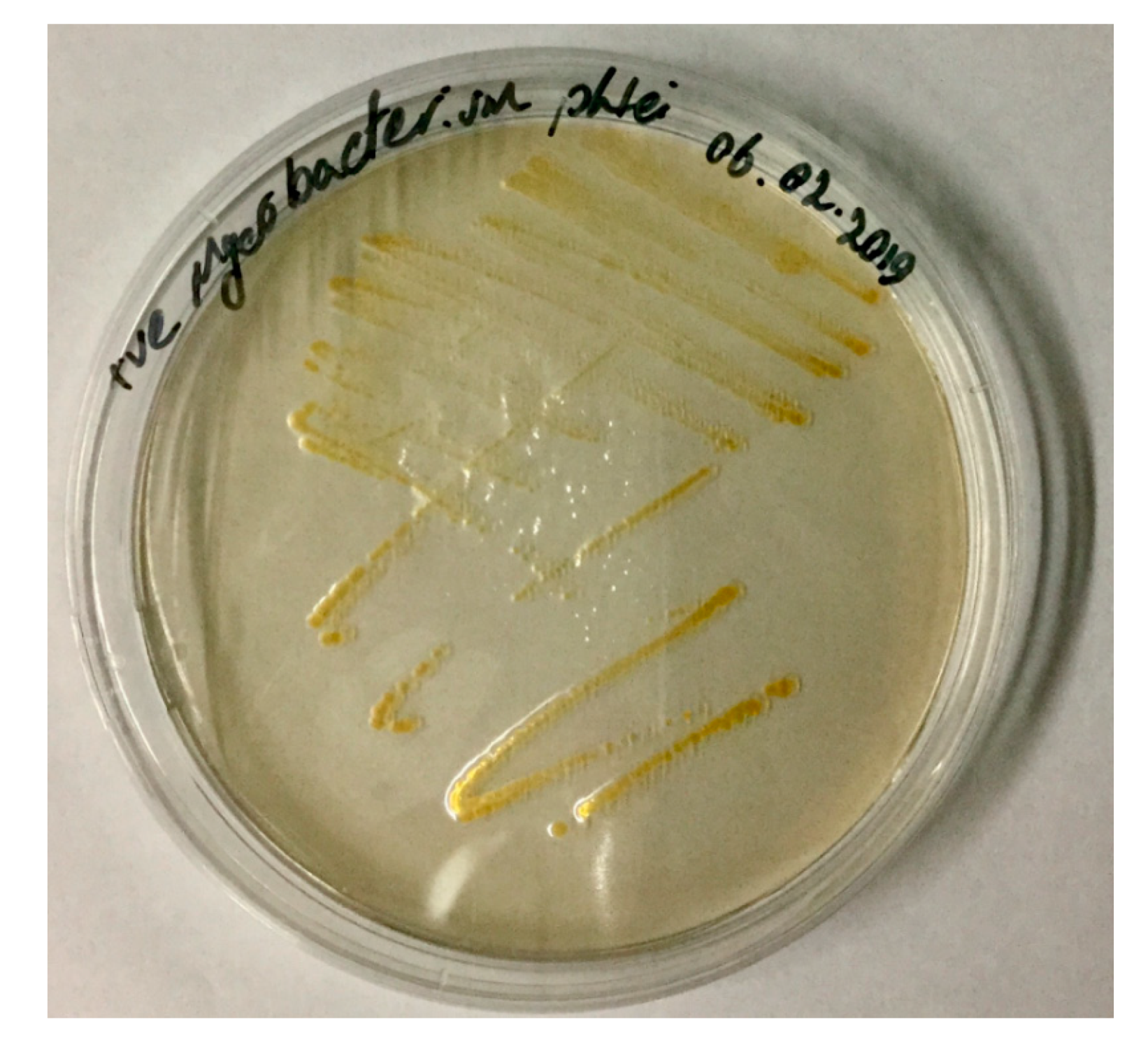
6.1. Isolation and Cultivation
6.2. Serological Diagnostics
6.3. Molecular Diagnostics
7. Virulence Factors
8. Treatment and Control of Mycobacteriosis in Fish
8.1. Vaccination against Mycobacteriosis in Fish
8.2. Antibiotherapy
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hashish, E.; Merwad, A.; Elgaml, S.; Amer, A.; Kamal, H.; Elsadek, A.; Marei, A.; Sitohy, M. Mycobacterium marinum infection in fish and man: Epidemiology, pathophysiology and management; a review. Vet. Q. 2018, 38, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Chinabut, S. Fish Disease and Disorders: Viral, Bacterial, and Fungal Infections; Woo, P.T., Bruno, D.W., Eds.; CAB International: Wallingford, UK, 1999. [Google Scholar]
- Gauthier, D.T.; Rhodes, M. Mycobacteriosis in fishes: A review. Vet. J. 2009, 180, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Aubry, A.; Mougari, F.; Reibel, F.; Cambau, E. Mycobacterium marinum. Tuberculosis and Nontuberculous Mycobacterial Infections, 7th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; Volume 5, pp. 735–752. [Google Scholar]
- Grange, J. Mycobacterium chelonei. Tubercle 1981, 62, 273–276. [Google Scholar] [CrossRef]
- Ross, A.J.; Brancato, F.P. Mycobacterium fortuitum Cruz from the tropical fish Hyphessobrycon innesi. J. Bacteriol. 1959, 78, 392–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescenko, P.; Matlova, L.; Dvorska, L.; Bartos, M.; Vavra, O.; Navratil, S.; Novotny, L.; Pavlik, I. Mycobacterial infection in aquarium fish. Vet. Med. (Praha) 2003, 48, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Janse, M.; Kik, M. Mycobacterium avium granulomas in a captive epaulette shark, Hemiscyllium ocellatum (bonnaterre). J. Fish Dis. 2012, 35, 935–940. [Google Scholar] [CrossRef]
- Gupta, T.; Fine-Coulson, K.; Karls, R.; Gauthier, D.; Quinn, F. Internalization of Mycobacterium shottsii and Mycobacterium pseudoshottsii by Acanthamoeba polyphaga. Can. J. Microbiol. 2013, 59, 570–576. [Google Scholar] [CrossRef]
- Fukano, H.; Wada, S.; Kurata, O.; Katayama, K.; Fujiwara, N.; Hoshino, Y. Mycobacterium stephanolepidis sp. Nov., a rapidly growing species related to Mycobacterium chelonae, isolated from marine teleost fish, Stephanolepis cirrhifer. Int. J. Syst. Evol. Microbiol. 2017, 67, 2811–2817. [Google Scholar] [CrossRef]
- James, A.; Hagan, B.; William, A. Tuberculosis of the Mexican Platyfish (Platypoecilus maculatus). J. Infect. Dis. 1942, 70, 248–252. [Google Scholar]
- Stanford, J.L. Serological and bacteriological investigation of Mycobacterium ranae (fortuitum). J. Bacteriol. 1969, 98, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Kubica, G.P.; Baess, I.; Gordon, R.E.; Jenkins, P.A.; Kwapinski, J.B.; McDurmont, C.; Pattyn, S.R.; Saito, H.; Silcox, V.; Stanford, J.L.; et al. A co-operative numerical analysis of rapidly growing mycobacteria. J. Gen. Microbiol. 1972, 73, 55–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novotny, L.; Halouzka, R.; Matlova, L.; Vavra, O.; Bartosova, L.; Slany, M.; Pavlik, I. Morphology and distribution of granulomatous inflammation in freshwater ornamental fish infected with mycobacteria. J. Fish Dis. 2010, 33, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Han, H.J.; Kim, J.H.; Jeon, C.H.; Kim, W.S.; Kim, D.H.; Jung, S.J.; Oh, M.J. Molecular and histopathological evidence of mycobacteriosis in paradise fish Macropodus opercularis imported into Korea. Fish. Aquat. Sci. 2013, 16, 165–169. [Google Scholar] [CrossRef]
- Puk, K.; Banach, T.; Wawrzyniak, A.; Adaszek, Ł.; Zietek, J.; Winiarczyk, S.; Guz, L. Detection of Mycobacterium marinum, M. peregrinum, M. fortuitum and M. abscessus in aquarium fish. J. Fish Dis. 2018, 41, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Francis-Floyd, R. Mycobacterial Infections of Fish; SRAC Publisher: Stoneville, MS, USA, 2011; pp. 1–12. [Google Scholar]
- Fukano, H.; Wada, S.; Kurata, O.; Mizuno, K.; Nakanaga, K.; Hoshino, Y. Nontuberculous mycobacteriosis in farmed thread-sail filefish Stephanolepis cirrhifer. Fish Pathol. 2015, 50, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Magee, J. Taxonomy of Mycobacteria. In Mycobacteria; Springer: Boston, MA, USA, 1998. [Google Scholar]
- Tortoli, E. Microbiological features and clinical relevance of new species of the genus Mycobacterium. Clin. Microbiol. Rev. 2014, 27, 727–752. [Google Scholar] [CrossRef] [Green Version]
- Tønjum, T.; Welty, D.B.; Jantzen, E.; Small, P.L. Differentiation of Mycobacterium ulcerans, M. marinum, and M. haemophilum: Mapping of their relationships to M. tuberculosis by fatty acid profile analysis, DNA-DNA hybridization, and 16S rRNA gene sequence analysis. J. Clin. Microbiol. 1998, 36, 918–925. [Google Scholar] [CrossRef] [Green Version]
- Han, X.Y.; Dé, I.; Jacobson, K.L. Rapidly growing mycobacteria: Clinical and microbiologic studies of 115 cases. Am. J. Clin. Pathol. 2007, 128, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Brocklebank, J.; Raverty, S.; Robinson, J. Mycobacteriosis in Atlantic salmon farmed in British Columbia. Can. Vet. J. 2003, 44, 486–489. [Google Scholar]
- Aro, L.; Correa, K.; Martinez, A.; Ildefonso, R.Y.J.M.; Yanez, J.M. Characterization of Mycobacterium salmoniphilum as causal agent of mycobacteriosis in Atlantic salmon, Salmo salar L., from a freshwater recirculation system. J. Fish Dis. 2014, 37, 341–348. [Google Scholar] [CrossRef]
- Zerihun, M.A.; Berg, V.; Lyche, J.L.; Colquhoun, D.J.; Poppe, T.T. Mycobacterium salmoniphilum infection in burbot Lota lota. Dis. Aquat. Organ. 2011, 95, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.T.; Whipps, C. Activity of antibiotics against Mycobacterium species commonly found in laboratory Zebrafish. J. Aquat. Anim. Health 2015, 27, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Prearo, M.; Zanoni, R.G.; Dall’Orto, B.C.; Pavoletti, E.; Florio, D.; Penati, V.; Ghittino, C. Mycobacterioses: Emerging pathologies in aquarium fish. Vet. Res. Commun. 2004, 28, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Slany, M.; Makovcova, J.; Jezek, P.; Bodnarova, M.; Pavlik, I. Relative prevalence of Mycobacterium marinum in fish collected from aquaria and natural freshwaters in central Europe. J. Fish Dis. 2014, 37, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Bartram, J.; Cotruvo, J.A.; Dufour, A.; Rees, G.; Pedley, S.; Water, S.; Organization, W.H. Pathogenic Mycobacteria in Water: A Guide to Public Health Consequences, Monitoring and Management; IWA Publishing: London, UK, 2004. [Google Scholar]
- Beran, V.; Matlova, L.; Dvorska, L.; Svastova, P.; Pavlik, I. Distribution of mycobacteria in clinically healthy ornamental fish and their aquarium environment. J. Fish Dis. 2006, 29, 383–393. [Google Scholar] [CrossRef]
- Mitchell, M.A. Mycobacterial Infections in Reptiles. Vet. Clin. Exot. Anim. Pract. 2012, 15, 101–111. [Google Scholar] [CrossRef]
- Soldati, G.; Lu, Z.H.; Vaughan, L.; Polkinghorne, A.; Zimmermann, D.R.; Huder, J.B.; Pospischil, A. Detection of mycobacteria and chlamydiae in granulomatous inflammation of reptiles: A retrospective study. Vet. Pathol. 2004, 41, 388–397. [Google Scholar] [CrossRef]
- Johansen, M.D.; Herrmann, J.L.; Kremer, L. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef]
- Whipps, C.M.; Lieggi, C.; Wagner, R. Mycobacteriosis in Zebrafish Colonies. ILAR J. 2012, 53, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Conroy, D.A. A report on the problem of bacterial fish diseases in the Argentine Republic. Bull. Off. Int. Epizoot. 1966, 65, 755–768. [Google Scholar]
- Decostere, A.; Hermans, K.; Haesebrouck, F. Piscine mycobacteriosis: A literature review covering the agent and the disease it causes in fish and humans. Vet. Microbiol. 2004, 99, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Whipps, C.M.; Dougan, S.T.; Kent, M.L. Mycobacterium haemophilum infections of zebrafish (Danio rerio) in research facilities. FEMS Microbiol. Lett. 2007, 270, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Harriff, M.J.; Bermudez, L.E.; Kent, M.L. Experimental exposure of zebrafish, Danio rerio (Hamilton), to Mycobacterium marinum and Mycobacterium peregrinum reveals the gastrointestinal tract as the primary route of infection: A potential model for environmental mycobacterial infection. J. Fish Dis. 2007, 30, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.W.; Ordal, E. Tuberculosis in Pacific Salmon and Steelhead Trout. Master’s Thesis, University of Washington, Seattle, WA, USA, 1958. [Google Scholar]
- Hedrick, R.P.; McDowell, T.; Groff, J. Mycobacteriosis in cultured striped bass from California. J. Wildl. Dis. 1987, 23, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Mutoji, K.N. Investigation into Mechanisms of Mycobacterial Transmission between Fish; University of Louisiana at Lafayette: Lafayette, LA, USA, 2011. [Google Scholar]
- Conroy, G.; Conroy, D. Acid-fast bacterial infection and its control in guppies (Lebistes reticulatus) reared on an ornamental fish farm in Venezuela. Vet. Rec. 1999, 144, 177–178. [Google Scholar] [CrossRef]
- Somsiri, T.; Puttinaowarat, S.; Soontornwit, S.; Lacharoje, S. Contamination of Mycobacterium spp. in Live Feeds. In Disease in Asian in Aquaculture V.; Fish Health Section, Asian Fisheries SocietyAsian Fisheries Society: Manila, The Philippines, 2005; pp. 227–235. [Google Scholar]
- Nenoff, P.; Uhlemann, R. Mycobacteriosis in mangrove killifish (Rivulus magdalenae) caused by living fish food (Tubifex tubifex) infected with Mycobacterium marinum. Dtsch. Tierarztl. Wochenschr. 2006, 113, 230–232. [Google Scholar] [PubMed]
- Collins, C.H.; Grange, J.M.; Noble, W.C.; Yates, M.D. Mycobacterium marinum infections in man. Epidemiol. Infect. 1985, 94, 135–149. [Google Scholar] [CrossRef] [Green Version]
- Wiens, G. Fish Diseases and Disorders: Viral, Bacterial and Fungal Infections, 2nd ed.; CAB International: Wallingford, UK, 2011; pp. 338–374. [Google Scholar]
- Swaim, L.E.; Connolly, L.E.; Volkman, H.E.; Humbert, O.; Born, D.E.; Ramakrishnan, L. Mycobacterium marinum infection of adult zebrafish causes caseating granulomatous tuberculosis and is moderated by adaptive immunity. Infect. Immun. 2006, 74, 6108–6117. [Google Scholar] [CrossRef] [Green Version]
- Evely, M.M.; Donahue, J.M.; Sells, S.F.; Loynachan, A.T. Ocular mycobacteriosis in a red-bellied piranha, Pygocentrus nattereri Kner. J. Fish Dis. 2011, 34, 323–326. [Google Scholar] [CrossRef]
- Keller, C.; Wenker, C.; Jermann, T.; Hirschi, R.; Schildger, B.; Meier, R.; Schmidt-Posthaus, H. Piscine mycobacteriosis—Involvement of bacterial species and reflection in pathology. Schweiz Arch Tierheilkd 2018, 160, 385–393. [Google Scholar] [CrossRef]
- Marzouk, M.S.M.; Essa, M.A.; El-Seedy, F.R.; Kenawy, A.M.; Abd El-Gawad, D.M. Epizootiological and histopathological studies on mycobacteriosis in some ornamental fishes. Glob. Vet. 2009, 3, 137–143. [Google Scholar]
- Avsever, M.L.; Çavuçoǧlu, C.; Eskiizmirliler, S.; Türe, M.; Korun, J.; Çamkerten, I. First isolation of Mycobacterium marinum from sea bass (Dicentrarchus labrax) and gilthead sea bream (Spams auratus) cultured in Turkey. Bull. Eur. Assoc. Fish Pathol. 2016, 36, 193–200. [Google Scholar]
- Van Der Sar, A.; Abdallah, A.; Sparrius, M.; Reinders, E.; Vandenbroucke-Grauls, C.E.; Bitter, W. Mycobacterium marinum strains can be divided into two distinct types based on genetic diversity and virulence. Infect. Immun. 2004, 72, 6306–6312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehane, L.; Rawlin, G.T. Topically acquired bacterial zoonoses from fish: A review. Med. J. Aust. 2000, 173, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Passantino, A.; Macrì, D.; Coluccio, P.; Foti, F.; Marino, F. Importation of mycobacteriosis with ornamental fish: Medico-legal implications. Travel Med. Infect. Dis. 2008, 6, 240–244. [Google Scholar] [CrossRef]
- Bhambri, S.; Bhambri, A.; Del Rosso, J.Q. Atypical mycobacterial cutaneous infections. Dermatol. Clin. 2009, 27, 63–73. [Google Scholar] [CrossRef]
- Hernandez-Divers, S.J.; Shearer, D. Pulmonary mycobacteriosis caused by Mycobacterium haemophilum and M. marinum in a royal python. J. Am. Vet. Med. Assoc. 2002, 220, 1661–1663. [Google Scholar] [CrossRef]
- Bouricha, M.; Castan, B.; Duchene-Parisi, E.; Drancourt, M. Mycobacterium marinum infection following contact with reptiles: Vivarium granuloma. Int. J. Infect. Dis. 2014, 21, 17–18. [Google Scholar] [CrossRef] [Green Version]
- Streit, M.; Böhlen, L.M.; Hunziker, T.; Zimmerli, S.; Tscharner, G.G.; Nievergelt, H.; Bodmer, T.; Braathen, L.R. Disseminated Mycobacterium marinum infection with extensive cutaneous eruption and bacteremia in an immunocompromised patient. Eur. J. Dermatol. 2006, 16, 79–83. [Google Scholar]
- Afzal, A.; Nadeem, M.; Aman, S.; Kazmi, A.H. Mycobacterium marinum infection: A case report. J. Pak. Assoc. Dermatol. 2009, 19, 48–51. [Google Scholar]
- Chung, J.; Ince, D.; Ford, B.A.; Wanat, K.A. Cutaneous Infections Due to Nontuberculosis Mycobacterium: Recognition and Management. Am. J. Clin. Dermatol. 2018, 19, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-S.; Chiu, C.-H.; Su, L.-H.; Chia, J.-H.; Lee, M.-H.; Chiang, P.-C.; Kuo, A.-J.; Wu, T.-L.; Leu, H.-S. Mycobacterium marinum infection in Taiwan. J. Microbiol. Immunol. Infect. 2002, 35, 42–46. [Google Scholar] [PubMed]
- O’Brien, D.P.; Jeanne, I.; Blasdell, K.; Avumegah, M.; Athan, E. The changing epidemiology worldwide of Mycobacterium ulcerans. Epidemiol. Infect. 2019, 147, e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.H.; Fadul, N.; Ashraf, M.S.; Siraj, D.S. Osteomyelitis Infection of Mycobacterium marinum: A Case Report and Literature Review. Case Rep. Infect. Dis. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Babamahmoodi, F.; Babamahmoodi, A.; Nikkhahan, B. Review of Mycobacterium marinum Infection Reported from Iran and Report of Three New Cases with Sporotrichoid Presentation. Iran. Red Crescent Med. J. 2014, 16, e10120. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Diaz, E.; Morfin-Otero, R.; Perez-Gomez, H.R.; Esparza-Ahumada, S.; Rodriguez-Noriega, E. Rapidly growing mycobacterial infections of the skin and soft tissues caused by M. fortuitum and M. chelonae. Curr. Trop. Med. Rep. 2018, 5, 162–169. [Google Scholar] [CrossRef]
- Otsuka, Y.; Fujino, T.; Mori, N.; Sekiguchi, J.I.; Toyota, E.; Saruta, K.; Kikuchi, Y.; Sasaki, Y.; Ajisawa, A.; Otsuka, Y.; et al. Survey of human immunodeficiency virus (HIV)-seropositive patients with mycobacterial infection in Japan. J. Infect. 2005, 51, 364–374. [Google Scholar] [CrossRef]
- Lapierre, S.; Toro, A.; Drancourt, M. Mycobacterium iranicum bacteremia and hemophagocytic lymphohistiocytosis: A case report. BMC Res. Notes 2017, 10, 327. [Google Scholar]
- Bhatty, M.A.; Turner, D.P.; Chamberlain, S.T. Mycobacterium marinum hand infection: Case reports and review of literature. Br. J. Plast. Surg. 2000, 53, 161–165. [Google Scholar] [CrossRef]
- Bartralot, R.; García-Patos, V.; Sitjas, D.; Rodríguez-Cano, L.; Mollet, J.; Martín-Casabona, N.; Coll, P.; Castells, A.; Pujol, R.M. Clinical patterns of cutaneous nontuberculous mycobacterial infections. Br. J. Dermatol. 2005, 152, 727–734. [Google Scholar] [CrossRef]
- Slany, M.; Jezek, P.; Bodnarova, M. Fish tank granuloma caused by Mycobacterium marinum in two aquarists: Two case reports. BioMed. Res. Int. 2013, 2013, 161329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucko, M.; Colorni, A. Mycobacterium marinum infections in fish and humans in Israel. J. Clin. Microbiol. 2005, 43, 892–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosty, T.S.; McDurmont, C. Isolation of acid-fast organisms from milk and oysters. Health Lab. Sci. 1975, 12, 16–19. [Google Scholar] [PubMed]
- Rhodes, M.W.; Kator, H.; Kaattari, I.; Gauthier, D.; Vogelbein, W.; Ottinger, C.A. Isolation and characterization of mycobacteria from striped bass Morone saxatilis from the Chesapeake Bay. Dis. Aquat. Organ. 2004, 61, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Röbbecke, R.; Janning, B.; Fischeder, R. Occurrence of mycobacteria in biofilm samples. Tuber. Lung Dis. 1992, 73, 141–144. [Google Scholar] [CrossRef]
- Bonamonte, D.; De Vito, D.; Vestita, M.; Delvecchio, S.; Ranieri, L.D.; Santantonio, M.; Angelini, G. Aquarium-borne Mycobacterium marinum skin infection. Report of 15 cases and review of the literature. Eur. J. Dermatol. 2013, 23, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Belić, M.; Miljković, J.; Marko, P.B. Sporotrichoid presentation of Mycobacterium marinum infection of the upper extremity. A case report. Acta Dermatovenerol. Alp. Panon. Adriat. 2006, 15, 135–139. [Google Scholar]
- Rhodes, M.W.; Kator, H.; McNabb, A.; Deshayes, C.; Reyrat, J.M.; Brown-Elliott, B.A.; Wallace, R.; Trott, K.A.; Parker, J.M.; Lifland, B.; et al. Mycobacterium pseudoshottsii sp. nov., a slowly growing chromogenic species isolated from Chesapeake Bay striped bass (Morone saxatilis). Int. J. Syst. Evol. Microbiol. 2005, 55, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Whipps, C.; Butler, W.; Pourahmad, F.; Watral, V.; Kent, M. Molecular systematics support the revival of Mycobacterium salmoniphilum (ex Ross 1960) sp. nov., nom. rev., a species closely related to Mycobacterium chelonae. Int. J. Syst. Evol. Microbiol. 2007, 57, 2525–2531. [Google Scholar] [CrossRef] [Green Version]
- Bhalla, G.S.; Sarao, M.S.; Kalra, D.; Bandyopadhyay, K.; John, A.R. Methods of phenotypic identification of non-tuberculous mycobacteria. Pract. Lab. Med. 2018, 12, e00107. [Google Scholar] [CrossRef]
- Sarli, G.; Morandi, F.; Zanoni, R.; Brunetti, B.; Mandrioli, L.; Florio, D.; Prearo, M. Comparison of immunohistochemistry and Ziehl-Neelsen for the detection of Mycobacterium infection in sea bass (Dicentrarchus labrax). Bull. Eur. Assoc. Fish Pathol. 2005, 25, 182–185. [Google Scholar]
- Cribier, B.; Aubry, A.; Caumes, E.; Cambau, E.; Jarlier, V.; Chosidow, O. Aspects histopathologiques de l’infection à Mycobacterium marinum. Ann. Dermatol. Venereol. 2011, 138, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Stavri, H.; Brânaru-Gheorghiu, M.; Moldovan, O.; Raileanu, M.; Popa, M.I.; Popa, L.; Ene, L. Rapid immunochromatographic serum assay of nontuberculous mycobacterial infections. Roum. Arch. Microbiol. Immunol. 2005, 9, 42. [Google Scholar]
- Correa, A.G.; Starke, J. Nontuberculous mycobacterial disease in children. Semin. Respir. Infect. 1996, 11, 262–271. [Google Scholar]
- Xaio, H.; Gillespie, S. Using RT qPCR for quantifying Mycobacteria marinum from in-vitro and in-vivo samples. In Antibiotic Resistance Protocols; Human Press: New York, NY, USA, 2018; pp. 137–145. [Google Scholar]
- Delghandi, M.; Menanteau-Ledouble, S.; Waldner, K.; El-Matbouli, M. Renibacterium salmoninarum and Mycobacterium spp.: Two bacterial pathogens present at low levels in wild brown trout (Salmo trutta fario) populations in Austrian rivers. BMC Vet. Res. 2020, 16, 40. [Google Scholar] [CrossRef] [Green Version]
- Seyfahmadi, M.; Moaddab, S.R.; Sabokbar, A. Identification of mycobacteria from unhealthy and apparently healthy aquarium fish using both conventional and PCR analyses of hsp65 gene. Thai J. Vet. Med. 2017, 47, 571–578. [Google Scholar]
- Ponpornpisit, A.; Areechon, N.; Kono, T.; Kitao, Y.; Sakai, M.; Katagiri, T.; Endo, M. Detection of Mycobacteriosis in guppy, Poecilia reticulata, by loop-mediated isothermal amplification method. Bull. Eur. Ass. Fish Pathol. 2009, 29, 3–9. [Google Scholar]
- Tsai, M.; Wang, P.; Yoshida, S.; Aono, A.; Mitarai, S. Establishment of loop-mediated isothermal ampli fi cation for rapid and convenient detection of Mycobacterium marinum complex. J. Microbiol. Methods 2019, 164, 105671. [Google Scholar] [CrossRef]
- Phung, T.; Caruso, D.; Godreuil, S.; Keck, N.; Vallaeys, T.; Avarre, J. Rapid detection and identification of nontuberculous mycobacterial pathogens in fish by using high-resolution melting analysis. Appl. Environ. Microbiol. 2013, 79, 7837–7845. [Google Scholar] [CrossRef] [Green Version]
- Salati, F.; Meloni, M.; Fenza, A.; Angelucci, G.; Colorni, A.; Orrù, G. A sensitive FRET probe assay for the selective detection of Mycobacterium marinum in fish. J. Fish Dis. 2010, 33, 47–56. [Google Scholar] [CrossRef]
- Kothavade, R.J.; Dhurat, R.S.; Mishra, S.N.; Kothavade, U.R. Clinical and laboratory aspects of the diagnosis and management of cutaneous and subcutaneous infections caused by rapidly growing mycobacteria. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 161–188. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, S.; Kabayama, J.; Fukuyasu, T.; Castillo, C.S.; Hikima, J.; Jung, T. Bacterial Classification of Fish-Pathogenic Mycobacterium Species by Multigene Phylogenetic Analyses and MALDI Biotyper Identification System. Mar. Biotechnol. 2013, 15, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Sturgill-koszycki, A.S.; Schlesinger, P.H.; Chakraborty, P.; Haddix, P.L.; Collins, H.L.; Fok, A.K.; Allen, R.D.; Gluck, S.L.; Heuser, J.; Russell, D.G. Lack of acidification in Mycobacterium phagosomes produced by exclusion of the vesicular proton-ATPase. Science 1994, 263, 678–681. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, M.I.; Pehau-Arnaudet, G.; Fretz, M.M.; Romain, F.; Bottai, D.; Brodin, P.; Honoré, N.; Marchal, G.; Jiskoot, W.; England, P.; et al. ESAT-6 from Mycobacterium tuberculosis dissociates from its putative chaperone CFP-10 under acidic conditions and exhibits membrane-lysing activity. J. Bacteriol. 2007, 189, 6028–6034. [Google Scholar] [CrossRef] [Green Version]
- Ruley, K.M.; Ansede, J.H.; Pritchett, C.L.; Talaat, A.M.; Reimschuessel, R.; Trucksis, M. Identification of Mycobacterium marinum virulence genes using signature-tagged mutagenesis and the goldfish model of mycobacterial pathogenesis. FEMS Microbiol. Lett. 2004, 232, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Weerdenburg, E.M.; Abdallah, A.M.; Rangkuti, F.; Abd El Ghany, M.; Otto, T.D.; Adroub, S.A.; Molenaar, D.; Ummels, R.; Ter Veen, K.; van Stempvoort, G.; et al. Genome-wide transposon mutagenesis indicates that Mycobacterium marinum customizes its virulence mechanisms for survival and replication in different hosts. Infect. Immun. 2015, 83, 1778–1788. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wang, H.; Meng, L.; Wang, Q.; Yu, J.; Gao, Q.; Wang, D. Role of eosinophils and apoptosis in PDIMs/PGLs deficient mycobacterium elimination in adult zebrafish. Dev. Comp. Immunol. 2016, 59, 199–206. [Google Scholar] [CrossRef]
- Sassetti, C.M.; Rubin, E.J. Genetic requirements for mycobacterial survival during infection. Proc. Natl. Acad. Sci. USA 2003, 100, 12989–12994. [Google Scholar] [CrossRef] [Green Version]
- Quigley, J.; Hughitt, V.K.; Velikovsky, C.A.; Mariuzza, R.A.; El-Sayed, N.M.; Briken, V. The cell wall lipid PDIM contributes to phagosomal escape and host cell exit of Mycobacterium tuberculosis. mBio 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Johansen, M.D.; Hortle, E.; Kasparian, J.A.; Romero, A.; Novoa, B.; Figueras, A.; Britton, W.J.; de Silva, K.; Purdie, A.C.; Oehlers, S.H. Analysis of mycobacterial infection-induced changes to host lipid metabolism in a zebrafish infection model reveals a conserved role for LDLR in infection susceptibility. Fish Shellfish Immunol. 2018, 83, 238–242. [Google Scholar] [CrossRef]
- Laencina, L.; Dubois, V.; Le Moigne, V.; Viljoen, A.; Majlessi, L.; Pritchard, J.; Bernut, A.; Piel, L.; Roux, A.L.; Gaillard, J.L.; et al. Identification of genes required for Mycobacterium abscessus growth in vivo with a prominent role of the ESX-4 locus. Proc. Natl. Acad. Sci. USA 2018, 115, E1002–E1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weerdenburg, E.M.; Abdallah, A.M.; Mitra, S.; de Punder, K.; van der Wel, N.N.; Bird, S.; Appelmelk, B.J.; Bitter, W.; van der Sar, A.M. ESX-5-deficient Mycobacterium marinum is hypervirulent in adult zebrafish. Cell. Microbiol. 2012, 14, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Ates, L.S.; Ummels, R.; Commandeur, S.; van der Weerd, R.; Sparrius, M.; Weerdenburg, E.; Alber, M.; Kalscheuer, R.; Piersma, S.R.; Abdallah, A.M.; et al. Essential Role of the ESX-5 Secretion System in Outer Membrane Permeability of Pathogenic Mycobacteria. PLoS Genet. 2015, 11, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Daleke, M.H.; Cascioferro, A.; De Punder, K.; Ummels, R.; Abdallah, A.M.; Van Der Wel, N.; Peters, P.J.; Luirink, J.; Manganelli, R.; Bitter, W. Conserved Pro-Glu (PE) and Pro-Pro-Glu (PPE) protein domains target LipY lipases of pathogenic mycobacteria to the cell surface via the ESX-5 pathway. J. Biol. Chem. 2011, 286, 19024–19034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensing, B.A.; Seepersaud, R.; Yen, Y.T.; Sullam, P.M. Selective transport by SecA2: An expanding family of customized motor proteins. Biochim. Biophys. Acta—Mol. Cell Res. 2014, 1843, 1674–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulauf, K.E.; Sullivan, J.T.; Braunstein, M. The SecA2 pathway of Mycobacterium tuberculosis exports effectors that work in concert to arrest phagosome and autophagosome maturation. PLoS Pathog. 2018, 14, 1–29. [Google Scholar] [CrossRef]
- Sullivan, J.T.; Young, E.F.; Mccann, J.R.; Braunstein, M. The Mycobacterium tuberculosis SecA2 system subverts phagosome maturation to promote growth in macrophages. Infect. Immun. 2012, 80, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, G.; Shrivastva, R.; Mukhopadhyay, S. Mycobacterial PknG Targets the Rab7l1 Signaling Pathway to Inhibit Phagosome–Lysosome Fusion. J. Immunol. 2018, 201, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- van der Woude, A.D.; Stoop, E.J.M.; Stiess, M.; Wang, S.; Ummels, R.; van Stempvoort, G.; Piersma, S.R.; Cascioferro, A.; Jiménez, C.R.; Houben, E.N.G.; et al. Analysis of secA2-dependent substrates in Mycobacterium marinum identifies protein kinase G (PknG) as a virulence effector. Cell. Microbiol. 2014, 16, 280–295. [Google Scholar] [CrossRef]
- Watkins, B.Y.; Joshi, S.A.; Ball, D.A.; Leggett, H.; Park, S.; Kim, J.; Austin, C.D.; Paler-Martinez, A.; Xu, M.; Downing, K.H.; et al. Mycobacterium marinum SecA2 promotes stable granulomas and induces tumor necrosis factor alpha in Vivo. Infect. Immun. 2012, 80, 3512–3520. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ru, H.; Xiang, Z.; Jiang, J.; Wang, Y.; Zhang, L.; Liu, J. WhiB4 Regulates the PE/PPE Gene Family and is Essential for Virulence of Mycobacterium marinum. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kato, G.; Kondo, H.; Aoki, T.; Hirono, I. BCG vaccine confers adaptive immunity against Mycobacterium sp. infection in fish. Dev. Comp. Immunol. 2010, 34, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Ziklo, N.; Colorni, A.; Gao, L.Y.; Du, S.J.; Ucko, M. Humoral and Cellular Immune Response of European Seabass Dicentrarchus labrax Vaccinated with Heat-Killed Mycobacterium marinum (iipA::kan Mutant). J. Aquat. Anim. Health 2018, 30, 312–324. [Google Scholar] [CrossRef] [PubMed]
- López, V.; Risalde, M.A.; Contreras, M.; Mateos-Hernández, L.; Vicente, J.; Gortázar, C.; de la Fuente, J. Heat–inactivated Mycobacterium bovis protects zebrafish against mycobacteriosis. J. Fish Dis. 2018, 41, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Risalde, M.A.; López, V.; Contreras, M.; Mateos-Hernández, L.; Gortázar, C.; de la Fuente, J. Control of mycobacteriosis in zebrafish (Danio rerio) mucosally vaccinated with heat-inactivated Mycobacterium bovis. Vaccine 2018, 36, 4447–4453. [Google Scholar] [CrossRef]
- Chen, S.C.; Yoshida, T.; Adams, A.; Thompson, K.D.; Richards, R.H. Immune response of rainbow trout to extracellular products of Mycobacterium spp. J. Aquat. Anim. Health 1996, 8, 216–222. [Google Scholar] [CrossRef]
- Myllymäki, H.; Niskanen, M.; Oksanen, K.E.; Sherwood, E.; Ahava, M.; Parikka, M.; Rämet, M. Identification of novel antigen candidates for a tuberculosis vaccine in the adult zebrafish (Danio rerio). PLoS ONE 2017, 12, 1–21. [Google Scholar] [CrossRef]
- Stinear, T.; Jenkin, G.; Johnson, P.; Davies, J. Comparative genetic analysis of Mycobacterium ulcerans and Mycobacterium marinum reveals evidence of recent divergence. J. Bacteriol. 2000, 182, 6322–6330. [Google Scholar] [CrossRef] [Green Version]
- Tanghe, A.; Denis, O.; Lambrecht, B.; Motte, V.; Van Den Berg, T.; Huygen, K. Tuberculosis DNA vaccine encoding Ag85A is immunogenic and protective when administered by intramuscular needle injection but not by epidermal gene gun bombardment. Infect. Immun. 2000, 68, 3854–3860. [Google Scholar] [CrossRef] [Green Version]
- Pasnik, D.J.; Smith, S. Immunogenic and protective effects of a DNA vaccine for Mycobacterium marinum in fish. Vet. Immunol. Immunopathol. 2005, 103, 195–206. [Google Scholar] [CrossRef]
- Cui, Z.; Samuel-Shaker, D.; Watral, V.; Kent, M.L. Attenuated Mycobacterium marinum protects zebrafish against mycobacteriosis. J. Fish Dis. 2010, 33, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toranzo, A.E.; Romalde, J.L.; Magariños, B.; Barja, J.L. Present and future of aquaculture vaccines against fish bacterial diseases. Options Mediterr. 2009, 86, 155–176. [Google Scholar]
- Muktar, Y.; Tesfaye, S. Present Status and Future Prospects of Fish Vaccination: A Review. J. Vet. Sci. Technol. 2016, 7, 299. [Google Scholar] [CrossRef]
- Adams, A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol. 2019, 90, 210–214. [Google Scholar] [CrossRef]
- Ramírez-Paredes, J.G.; Mendoza-Roldan, M.A.; Lopez-Jimena, B.; Shahin, K.; Metselaar, M.; Thompson, K.D.; Adams, A. Whole cell inactivated autogenous vaccine effectively protects red Nile tilapia (Oreochromis niloticus) against francisellosis via intraperitoneal injection. J. Fish Dis. 2019, 42, 1191–1200. [Google Scholar] [CrossRef]
- Kawakami, K.; Kusuda, R. Efficacy of rifampicin, streptomycin and erythromycin against experimental Mycobacterium infection in cultured yellowtail. Nippon Suisan Gakkaishi Bull. Jpn. Soc. Sci. Fish. 1990, 56, 51–53. [Google Scholar] [CrossRef]
- Bragg, R.R.; Huchzermeyer, H.F.; Hanisch, M.A. Mycobacterium fortuitum isolated from three species of fish in South Africa. Onderstepoort J. Vet. Res. 1990, 57, 101–102. [Google Scholar]
- Astrofsky, K.M.; Schrenzel, M.D.; Bullis, R.A.; Smolowitz, R.M.; Fox, J.G. Diagnosis and Management of Atypical Mycobacterium spp. Infections in (Brachydanio rerio) Facilities. Comp. Med. 2000, 50, 666–672. [Google Scholar]
- Chang, C.T.; Doerr, K.M.; Whipps, C.M. Antibiotic treatment of zebrafish mycobacteriosis: Tolerance and efficacy of treatments with tigecycline and clarithromycin. J. Fish Biol. 2017, 40, 1473–1485. [Google Scholar] [CrossRef]
- Menanteau-ledouble, S.; Krauss, I.; Alexandre, R.; Weber, B.; Abreu, G.; El-matbouli, M. Research in Veterinary Science Antimicrobial effect of the Biotronic® Top3 supplement and efficacy in protecting rainbow trout (Oncorhynchus mykiss) from infection by Aeromonas salmonicida subsp. salmonicida. Res. Vet. Sci. 2017, 114, 95–100. [Google Scholar] [CrossRef]
- Ghanei-Motlagh, R.; Mohammadian, T.; Gharibi, D.; Menanteau-Ledouble, S.; Mahmoudi, E.; Khosravi, M.; Zarea, M.; El-Matbouli, M. Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer. Mar. Drugs 2020, 18, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljubobratovic, U.; Kosanovic, D.; Vukotic, G.; Molnar, Z.; Stanisavljevic, N.; Ristovic, T.; Peter, G.; Lukic, J.; Jeney, G. Supplementation of lactobacilli improves growth, regulates microbiota composition and suppresses skeletal anomalies in juvenile pike-perch (Sander lucioperca) reared in recirculating aquaculture system (RAS): A pilot study. Res. Vet. Sci. 2017, 115, 451–462. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Aquatic Host | Environment | Zoonotic Potential |
|---|---|---|---|
| M. szulgai | Crocodile | Fresh water | Yes |
| African clawed frogs (Xenopus tropica) | |||
| M. septicum/ | Zebrafish (Danio rerio), Cichlid (Pseudotopheus lombardoi), Koi fish | Fresh water | Yes |
| M. peregrinum | Labidochromis caeruleus, Black mollies (Poecilia sphenops), guppies (Poecilia reticulata), green swordtails (Xiphophorus hellerii), catfish (Pangasius hypophthalmus) | ||
| M. chelonae | Multiple | Fresh and Marine water | Yes |
| M. avium | Dwarf Cichlid (Apistogramma cacatuodes) | Fresh water | Yes |
| M. abscessus | Zebrafish (Danio rerio) | Fresh water | Yes |
| Medaka (Oryzias latipes) | |||
| Milkfish (Chanos chanos) | |||
| German blue ram (Mikrogeophagus ramirezi) | |||
| M. haemophilum | Zebrafish (Danio rerio) | Fresh water | Yes |
| M. lentiflavum | Swordtail (Xiphophorus hellerii) | Fresh water | Yes |
| M. gordonae | Goldfish (Carassius auratus) | Marine water | Yes |
| Guppy (Poecilia reticulate) | |||
| Angel fish (Pterophyllum scalare) | |||
| M. chesapeaki | Striped bass (Morone saxatilis) | Marine water | Unknown |
| M. marinum | Multiple | Fresh and marine water | Yes |
| M. montefiorence | Moray eel (Gymnothorax funebris) | Marine water | Unknown |
| M. neoaurum | Chinook salmon (Oncorhynchus tschawytscha) | Marine water | Yes |
| M. shottsii | Striped bass (Morone saxatilis) | Freshwater | Unknown |
| M. pseudoshottsii | Yellow tail (Seriola quinqueradiata), Greater amberjack (Seriola dumerili), Striped jack (Pseudocaranx dentex) | Marine water | Unknown |
| M. fortuitum | Neon tetra (Paracheirodon innesi) | Freshwater | Yes |
| M. syngnathidarum | syngnathid fish | Marine water | Unknown |
| M. flavescens | Tiger Oscar Astronatus ocellatus | Fresh water | Yes |
| M. smegmatis | Poecilia sphenop, Poecilia reticulata, Gymnocorymbus ternetzi | Fresh water | Yes |
| M. nonchromogenicum | Betta splendens, Carassius auratus, Poecilia reticulata, Pterophyllum scalare | Fresh water | No |
| M. stephanolepidis | Filefish (Stephanolepis cirrhifer) | Fresh water | Unknown |
| M. holsaticum | Silver moony fish (Monodactylus argenteus) | Marine water | Yes |
| M. salmoniphilum | Burbot (Lota lota) | Fresh water | No |
| Biochemical Tests | Results |
|---|---|
| Growth in NaCl 5% | Negative (M. gordonae, M. ulcerans, M. kansasii, M. marinum, M. simiae, M. szulgai, M. scrofulaceum, M. xenopi, M. avium, M. intracellulare, M. chelonae, M. diernhoferi, M. celatum, M. terrae) |
| Arylsulfatase | Negative (M. avium, M. intracellulare, M. smegmatis, M. kansasii, M. simiae, M. szulgai, M. scrofulaceum, M. asiaticum, M terrae, M. gordonae) (weakly positive after 3 day) |
| Catalase | Positive (M. fortuitum, M. chelonae, M. abscessus, M. smegmatis, M. kansasii, M. marinum, M. ulcerans, M. simiae, M. szulgai, M. scrofulaceum, M. gordonae) |
| Nitrate reduction | Negative (M. avium, M. intracellulare, M. chelonae, M. abscessus, M. ulcerans, M. simiae, M. scrofulaceum, M. gordonae, M. xenopi, M. celatum) |
| Urease activity | positive (M. marinum, M. fortuitum, M. chelonae, M. abscessus, M. kansasii, M. simiae, M. szulgai, M. scrofulaceum, M. flavescens) |
| Pirazinamidase | Positive |
| Thiophene-2-carboxylic hydrazide | Positive |
| Target Gene | Primer Name | Sequence (5′ to 3′) |
|---|---|---|
| dnaJ1 | J10F | CGIGARTGGGTYGARAARG |
| J335R | ARICCICCGAAIARRTCICC | |
| secA1 | MtuF1 | GACAGYGAGTGGATGGGYCGSGTGCACCG |
| MtuR3 | ACCACGCCCAGCTTGTAGATCTCGTGCAGCTC | |
| 32-kDa protein genes | MV1 | GGCCAGTCAAGCTTCTACTCCGACTGG |
| MV2 | GCCGTTGCCGCAGTACACCCAGACGCG | |
| hsp65 (heat-shock protein 65) | 21M13F | ACCAACGATGGTGTG TCCAT |
| 21M13R | CTTGTCGAACCGCATACCCT | |
| Tb11 | ACCAACGATGGTGTGTCCAT | |
| Tb12 | CTTGTCGAACCGCATACCCT | |
| erp (exported repeated protein) | erp-C1 | GCTCTA GACGAGCGGTCATCGGTTGCATAGGG |
| erp-C2 | GCTCTAGATTAGGCGACCGGCACGGTGATTGG | |
| erp-C3 | CGGAATTCATGGTGCTCGGGCCGCTC | |
| erp-C4 | CGGAATTCACCCAGG CCGCGCTGGTCACC | |
| erp-C6 | GCTCTAGATCAGGCAGGCGGCGGCACGGGTGC | |
| erp-C5 | CGGAATTCAAAC AAGCAGCATCGATAGCC | |
| erp-C7 | GCTCTAGACTAC GTGACAGGAATCAGTGATAT | |
| erp-8 | GTGCCGAACCGACGCCGACG | |
| erp-9 | GGCACCGGCGGCAGGTTGATCCCG | |
| rpoB (RNA polymerase B subunit) | MycoF | GGCAAGGTCACCCCGAAGGG |
| MycoR | AGCGGCTGCTGGGTGATC ATC | |
| RPO5′ | TCAAGGAGAAGCGATACGA | |
| RPO3′ | GGATGTTGATCAGGGTCTGC | |
| rec A | recF1 | GGTGGTCGNCTANTGTGGTG |
| recR1 | AGCTGGTTGATGAAGATYGC | |
| recF2 | GYGTCACSGCCAACCGAY | |
| recR2 | TTGATCTTCTTCTCGATCTC | |
| recF3 | GGCAARGGYTCGGTSATG | |
| sodA | sodlgF | GAAGGAATCTCGTGGCTGAATAC |
| sodlgR | AGTCGGCCTTGACGTTCTTGTAC | |
| erm (Erythromycin ribosome methyltransferase gene) | ermF | GACCGGGGCCTTCTTCGTGAT |
| ermR1 | GACTTCCCCGCACCGATTCC | |
| 16S-23S internal transcribed spacer (ITS) | ITS-F | CCTTTCTAAGGAGCACC |
| ITS-R | GATGCTCGCAACCACTATCC | |
| 16S rRNA | T39 | GCGAACGGTGAGTAACACG |
| T13 | TGCACACAGGCCACAAGGGA | |
| T43 | AATGGGCGCAAGCCTGATG | |
| T531 | ACCGCTACACCAGGAAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delghandi, M.R.; El-Matbouli, M.; Menanteau-Ledouble, S. Mycobacteriosis and Infections with Non-tuberculous Mycobacteria in Aquatic Organisms: A Review. Microorganisms 2020, 8, 1368. https://doi.org/10.3390/microorganisms8091368
Delghandi MR, El-Matbouli M, Menanteau-Ledouble S. Mycobacteriosis and Infections with Non-tuberculous Mycobacteria in Aquatic Organisms: A Review. Microorganisms. 2020; 8(9):1368. https://doi.org/10.3390/microorganisms8091368
Chicago/Turabian StyleDelghandi, Mohammad Reza, Mansour El-Matbouli, and Simon Menanteau-Ledouble. 2020. "Mycobacteriosis and Infections with Non-tuberculous Mycobacteria in Aquatic Organisms: A Review" Microorganisms 8, no. 9: 1368. https://doi.org/10.3390/microorganisms8091368