Lifestyle, Lineage, and Geographical Origin Influence Temperature-Dependent Phenotypic Variation across Yeast Strains during Wine Fermentation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
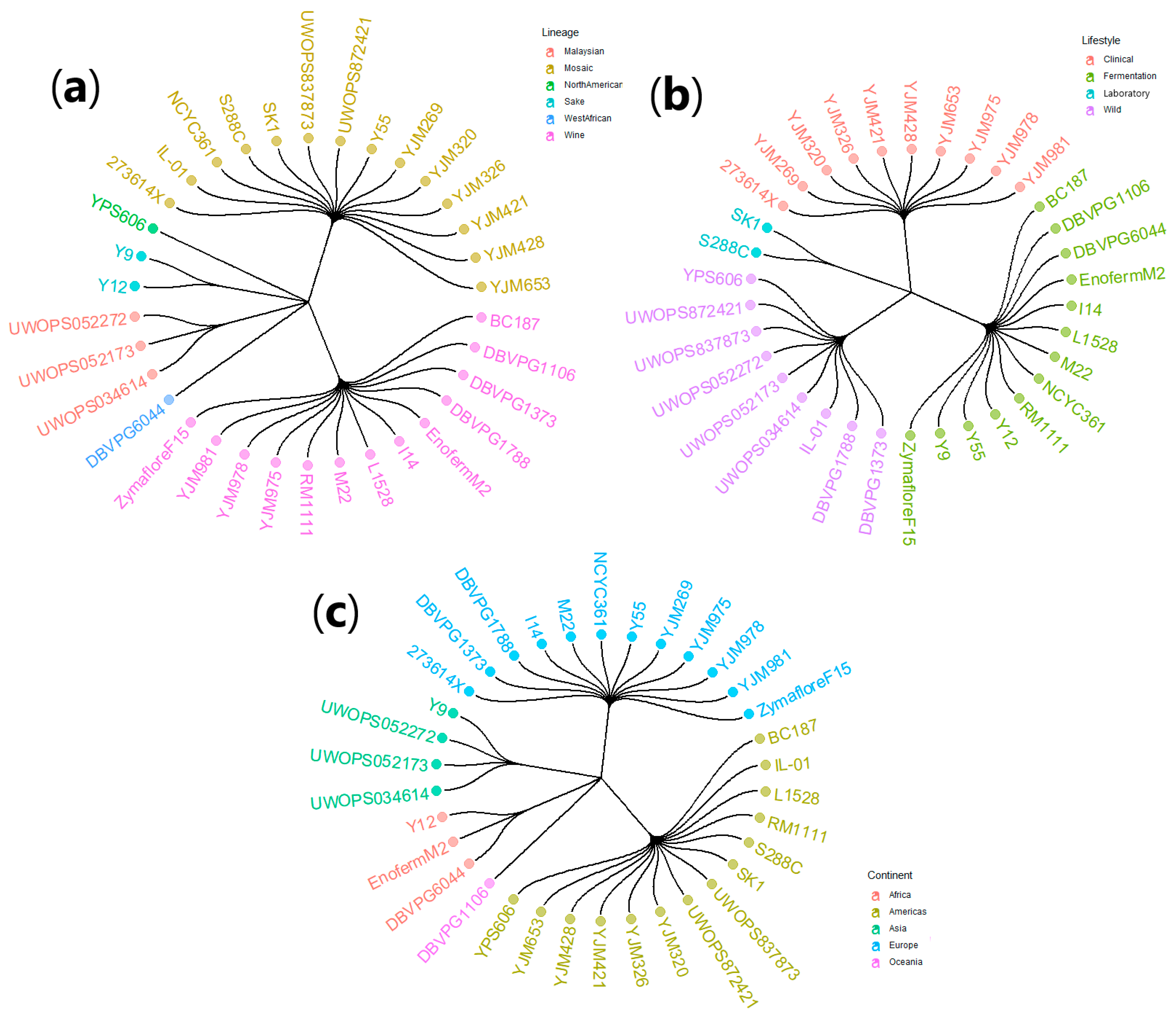
2.1. Yeast Strains
2.2. Growth and Fermentation Conditions
2.3. Calculation of Kinetic Parameters and Statistical Treatment of the Data
3. Results
3.1. S. cerevisiae Strains Show a Range of Fermentation Kinetics in Grape Juice at Different Temperatures
3.2. Statistical Analysis for Each Kinetic Variable
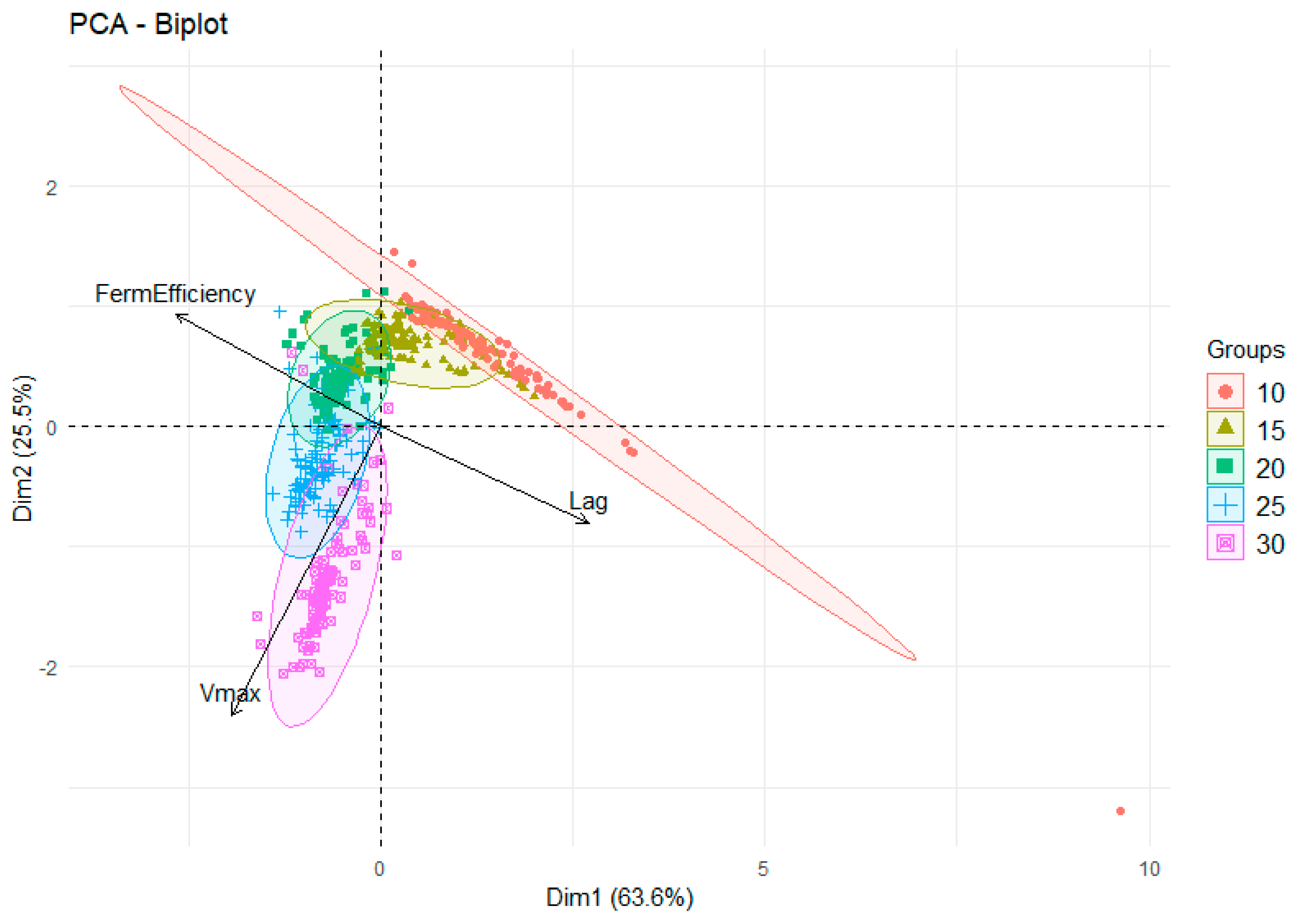
3.3. Multivariate Analysis of Lag Time Using Temperature, Lifestyle, Genetic Lineage, Continent, and Strain as Factors
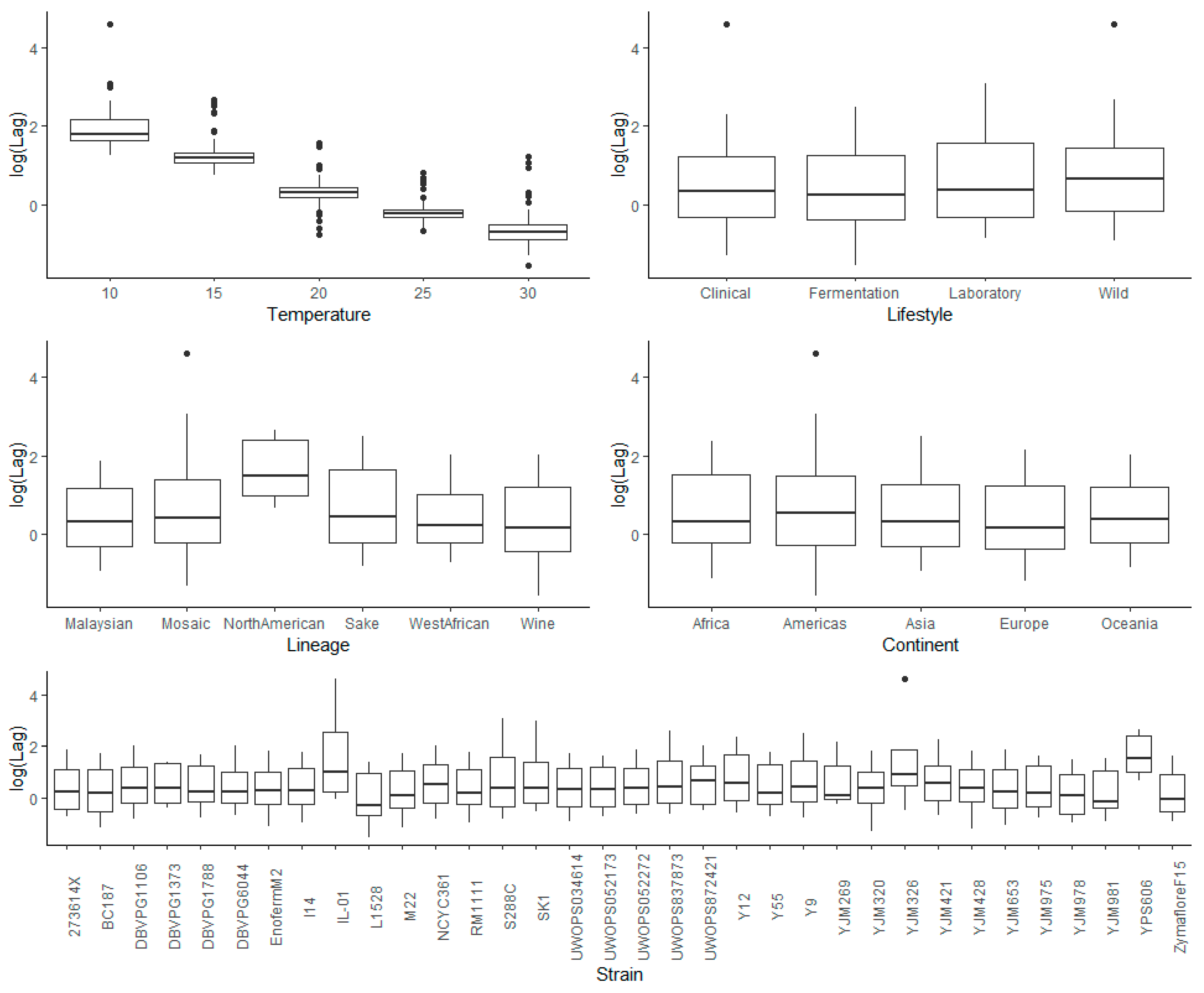
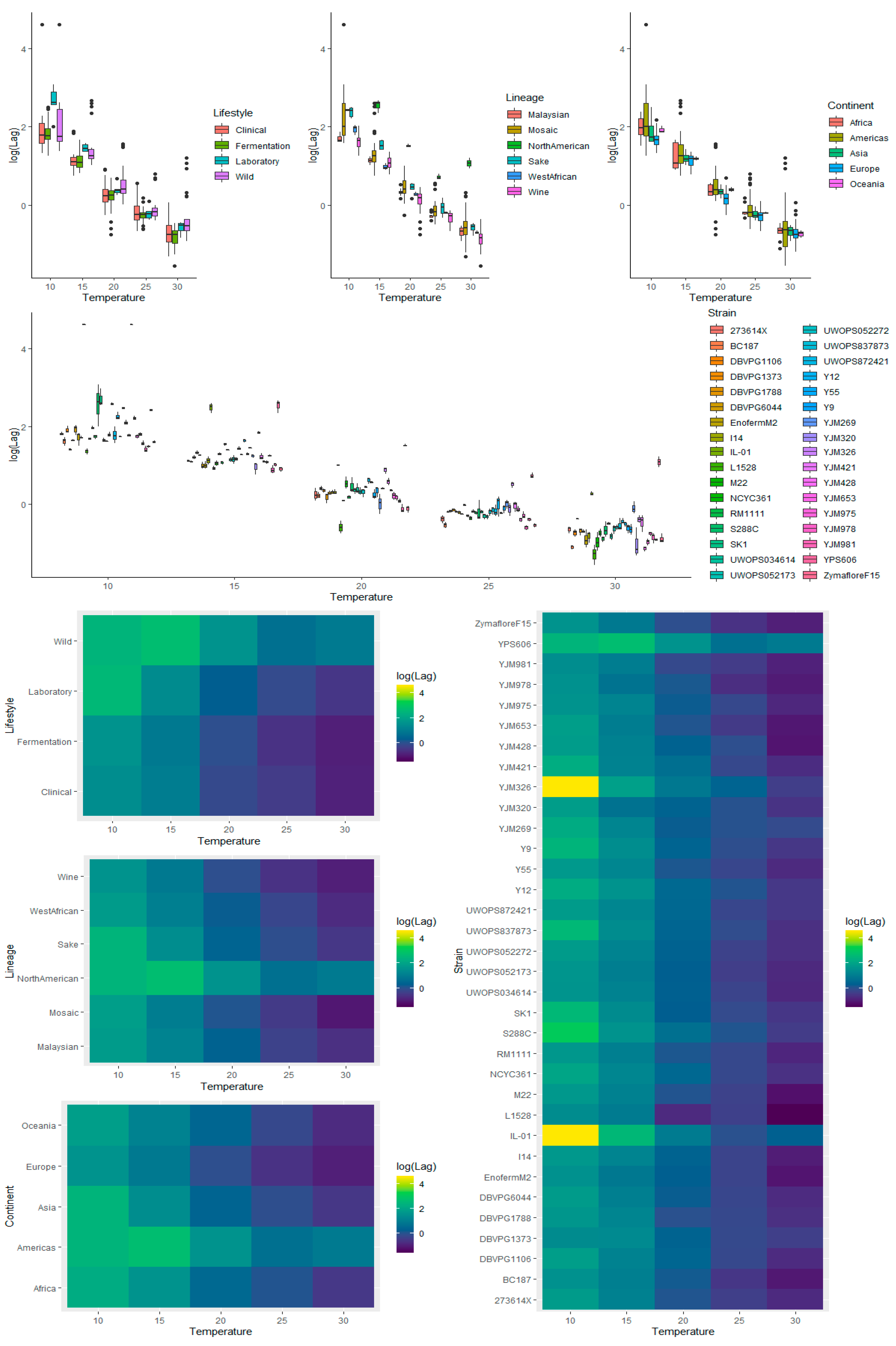
3.3.1. Temperature
3.3.2. Lifestyle
3.3.3. Lineage
3.3.4. Continent
3.3.5. Strain
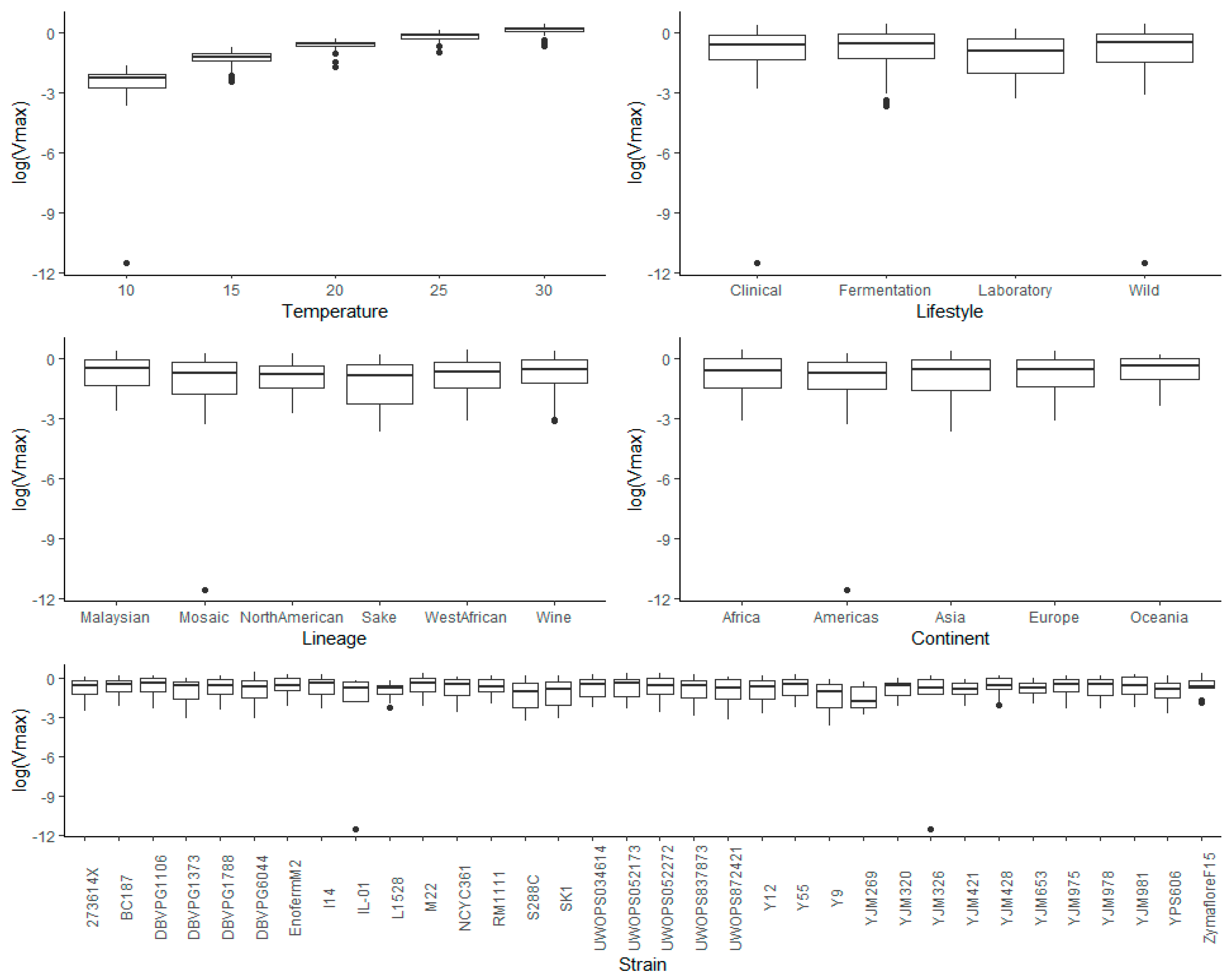
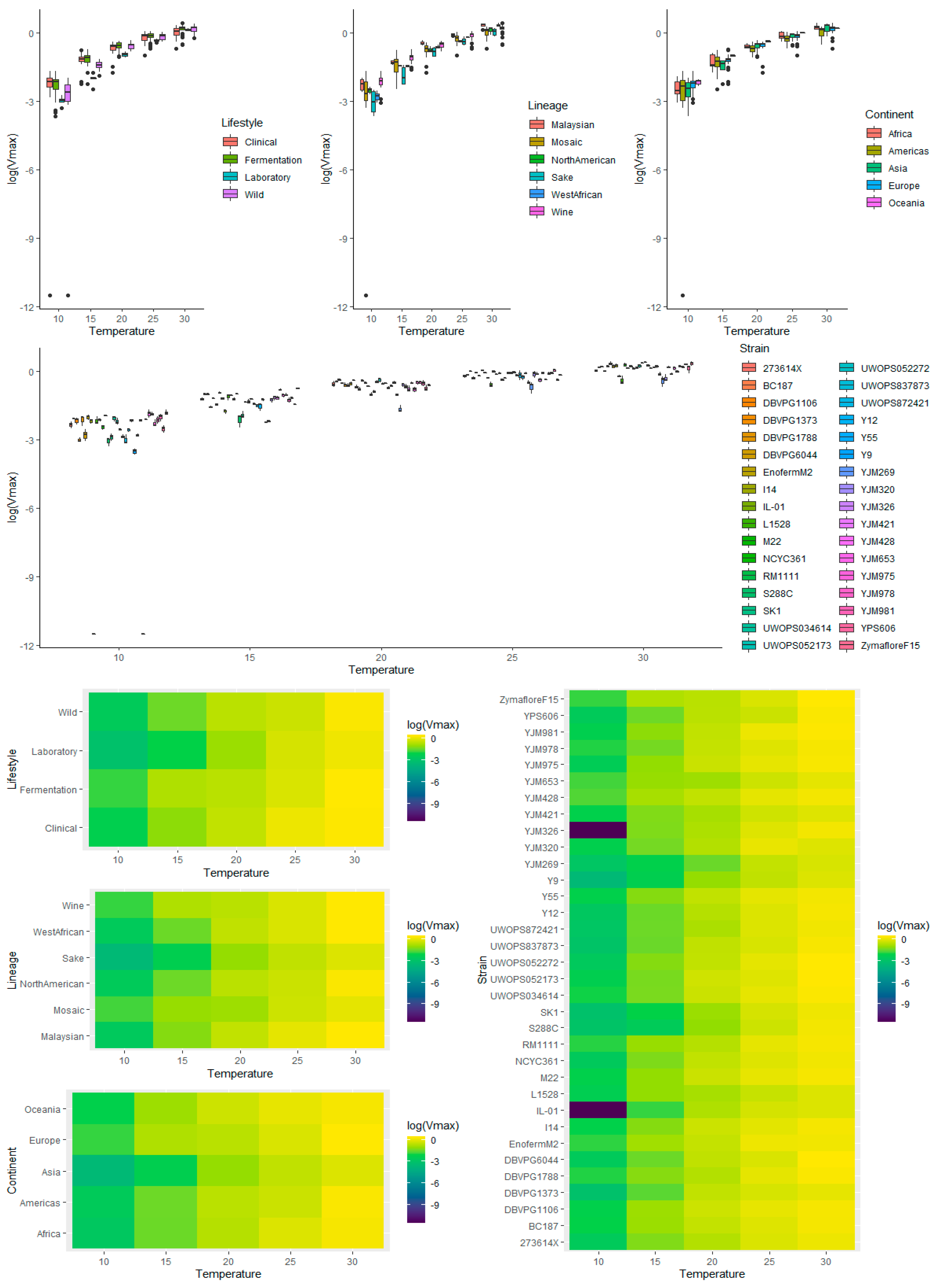
3.4. Multivariate Analysis of Vmax Using Temperature, Lifestyle, Genetic Lineage, Continent, and Strain as Factors
3.4.1. Temperature
3.4.2. Lifestyle
3.4.3. Lineage
3.4.4. Continent
3.4.5. Strain
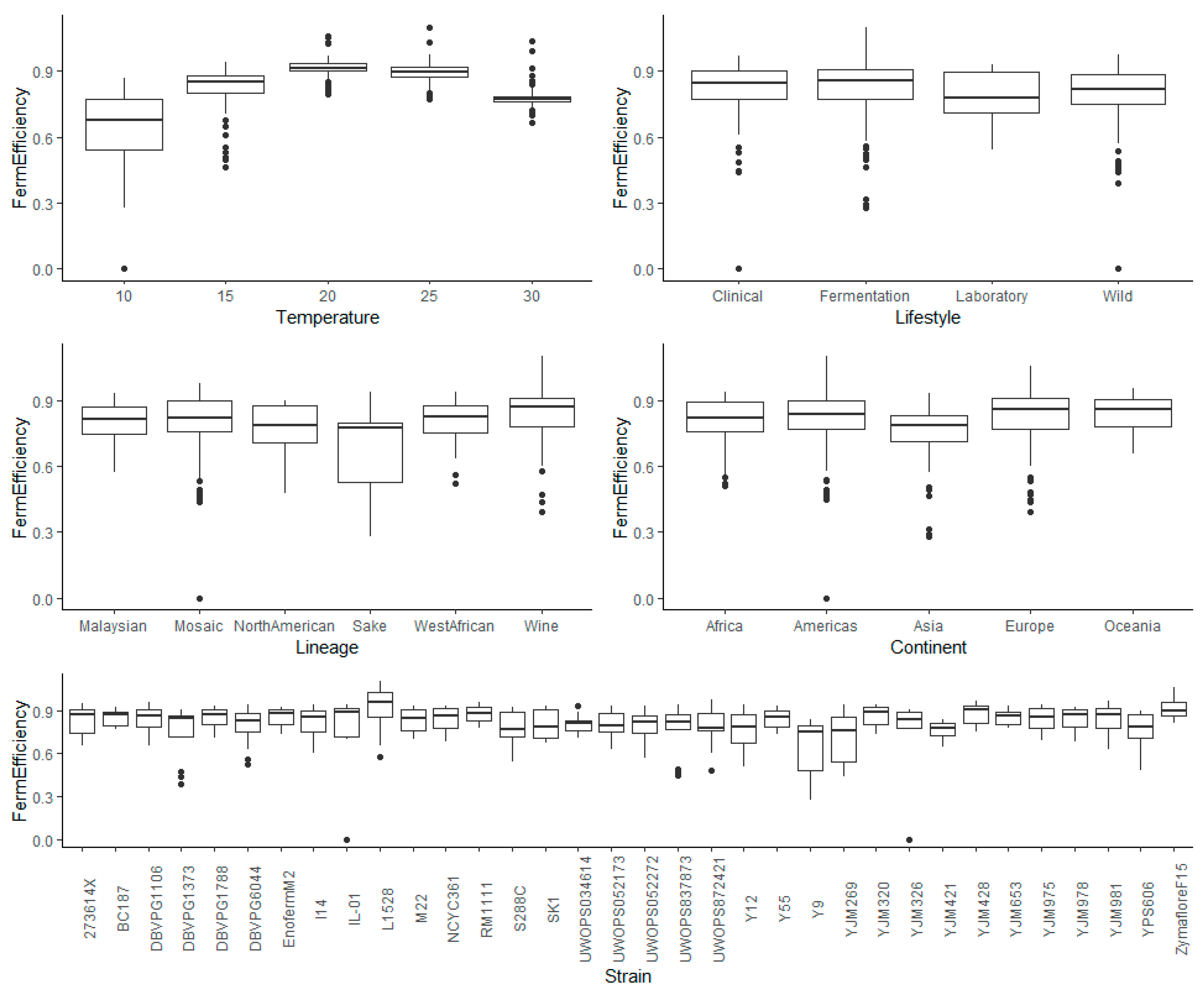
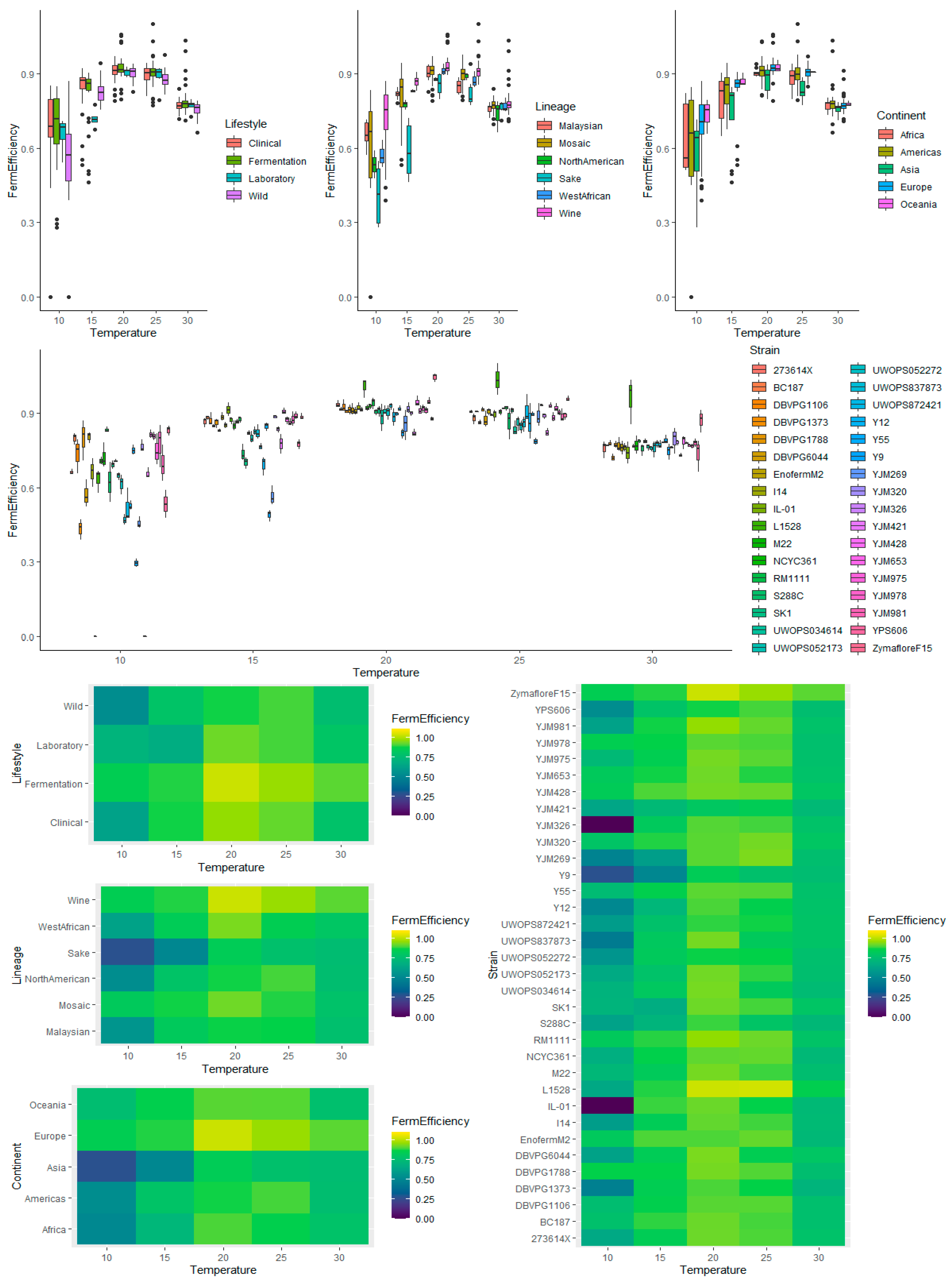
3.5. Multivariate Analysis of Fermentation Efficiency Using Temperature, Lifestyle, Genetic Lineage, Continent, and Strain as Factors
3.5.1. Temperature
3.5.2. Lifestyle
3.5.3. Lineage
3.5.4. Continent
3.5.5. Strain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bisson, L.F. The biotechnology of wine yeast. Food Biotechnol. 2004, 18, 63–96. [Google Scholar] [CrossRef]
- Bauer, F.F.; Pretorius, I.S. Yeast stress response and fermentation efficiency: How to survive the making of a wine—A review. S. Afr. J. Enol. Vitic. 2000, 21, 27–51. [Google Scholar] [CrossRef]
- Marullo, P.; Bely, M.; Masneuf-Pomarède, I.; Pons, M.; Aigle, M.; Dubourdieu, D. Breeding strategies for combining fermentative qualities and reducing off-flavor production in a wine yeast model. FEMS Yeast Res. 2006, 6, 268–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Bely, M.; Sablayrolles, J.M.; Barre, P. Description of alcoholic fermentation kinetics: Its variability and significance. Am. J. Enol. Vitic. 1990, 41, 319–324. [Google Scholar]
- Coleman, M.C.; Fish, R.; Block, D.E. Temperature-dependent kinetic model for nitrogen-limited wine fermentations. Appl. Environ. Microbiol. 2007, 73, 5875–5884. [Google Scholar] [CrossRef] [Green Version]
- Beltran, G.; Rozes, N.; Mas, A.; Guillamon, J.M. Effect of low-temperature fermentation on yeast nitrogen metabolism. World J. Microbiol. Biotechnol. 2007, 23, 809–815. [Google Scholar] [CrossRef]
- Charoenchai, C.; Fleet, G.H.; Henschke, P.A. Effects of temperature, pH, and sugar concentration on the growth rates and cell biomass of wine yeasts. Am. J. Enol. Vitic. 1998, 49, 283–288. [Google Scholar]
- Torija, M.J.; Rozes, N.; Poblet, M.; Guillamon, J.M.; Mas, A. Effects of fermentation temperature on the strain population of Saccharomyces cerevisiae. Int. J. Food Microbiol. 2003, 80, 47–53. [Google Scholar] [CrossRef]
- Pizarro, F.J.; Jewett, M.C.; Nielsen, J.; Agosin, E. Growth temperature exerts differential physiological and transcriptional responses in laboratory and wine strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2008, 74, 6358–6368. [Google Scholar] [CrossRef] [Green Version]
- Deed, R.C.; Deed, N.K.; Gardner, R.C. Transcriptional response of Saccharomyces cerevisiae to low temperature during wine fermentation. Antonie Leeuwenhoek 2015, 107, 1029–1048. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, F.; Varela, C.; Martabit, C.; Bruno, C.; Perez-Correa, J.R.; Agosin, E. Coupling kinetic expressions and metabolic networks for predicting wine fermentations. Biotechnol. Bioeng. 2007, 98, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Tronchoni, J.; Gamero, A.; Arroyo-López, F.N.; Barrio, E.; Querol, A. Differences in the glucose and fructose consumption profiles in diverse Saccharomyces wine species and their hybrids during grape juice fermentation. Int. J. Food Microbiol. 2009, 134, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.W.; Oliver, S.G.; Harrison, D.E.F.; Righelato, R.C. Ethanol inhibition of yeast growth and fermentation: Differences in the magnitude and complexity of the effect. Eur. J. Appl. Microbiol. Biotechnol. 1981, 11, 151–155. [Google Scholar] [CrossRef]
- Viegas, C.A.; Rosa, M.F.; Sa-Correia, I.; Novais, J.M. Inhibiton of yeast growth by octanoic and decanoic acids produced during ethanolic fermentation. Appl. Environ. Microbiol. 1989, 55, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liti, G. The fascinating and secret wild life of the budding yeast S. cerevisiae. eLife 2015, 4, e05835. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.V.; Petsko, G.A.; Johnston, G.C.; Ringe, D.; Singer, R.A.; Werner-Washburne, M. “Sleeping beauty”: Quiescence in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2004, 68, 187–206. [Google Scholar] [CrossRef] [Green Version]
- Fay, J.C.; Benavides, J.A. Evidence for domesticated and wild populations of Saccharomyces cerevisiae. PLoS Genet. 2005, 1, e5. [Google Scholar] [CrossRef]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Schacherer, J.; Shapiro, J.A.; Ruderfer, D.M.; Kruglyak, L. Comprehensive polymorphism survey elucidates population structure of Saccharomyces cerevisiae. Nature 2009, 458, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Goddard, M.R.; Greig, D. Saccharomyces cerevisiae: A nomadic yeast with no niche? FEMS Yeast Res. 2015, 15, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sardi, M.; Krause, M.; Heilberger, J.; Gasch, A.P. Genotype-by-environment-by-environment interactions in the Saccharomyces cerevisiae transcriptomic response to alcohols and anaerobiosis. G3 (Bethesda) 2018, 8, 3881–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borneman, A.R.; Desany, B.A.; Riches, D.; Affourtit, J.P.; Forgan, A.H.; Pretorius, I.S.; Egholm, M.; Chambers, P.J. Whole-genome comparison reveals novel genetic elements that characterize the genome of industrial strains of Saccharomyces cerevisiae. PLoS Genet. 2011, 7, e1001287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, R.K.; Johnston, J.R. Genealogy of principal strains of the yeast genetic stock center. Genetics 1986, 113, 35–43. [Google Scholar]
- Deed, R.C.; Fedrizzi, B.; Gardner, R.C. Saccharomyces cerevisiae FLO1 gene demonstrates genetic linkage to increased fermentation rate at low temperatures. G3 (Bethesda) 2017, 7, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Baty, F.; Ritz, C.; Charles, S.; Brutsche, M.; Flandrois, J.; Delignette-Muller, M. A Toolbox for Nonlinear Regression in R: The Package nlstools. J. Stat. Softw. 2015, 66, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Chang, W. ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, Package Version 1.0.5; R Development Core Team: Vienna, Austria, 2017. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Camarasa, C.; Sanchez, I.; Brial, P.; Bigey, F.; Dequin, S. Phenotypic landscape of Saccharomyces cerevisiae during wine fermentation: Evidence for origin-dependent metabolic traits. PLoS ONE 2011, 6, e25147. [Google Scholar] [CrossRef] [Green Version]
- Lainioti, G.C.; Kapolos, J.; Koliadima, A.; Karaiskakis, G. The study of the effect of fermentation temperature on the growth kinetics of Saccharomyces cerevisiae yeast strain, in the presence or absence of support, by chromatographic techniques. J. Liq. Chromatogr. Relat. Technol. 2011, 34, 195–208. [Google Scholar] [CrossRef]
- Ferreira, D.; Galeote, V.; Sanchez, I.; Legras, J.L.; Ortiz-Julien, A.; Dequin, S. Yeast multistress resistance and lag-phase characterisation during wine fermentation. FEMS Yeast Res. 2017, 17, fox051. [Google Scholar] [CrossRef] [Green Version]
- Gerke, J.P.; Chen, C.T.L.; Cohen, B.A. Natural isolates of Saccharomyces cerevisiae display complex genetic variation in sporulation efficiency. Genetics 2006, 174, 985–997. [Google Scholar] [CrossRef] [Green Version]
- El Haloui, N.E.; Corrieu, G.; Cleran, Y.; Cheruy, A. Method for on-line prediction of kinetics of alcoholic fermentation in wine making. J. Ferment. Bioeng. 1989, 68, 131–135. [Google Scholar] [CrossRef]
- Deed, R.C.; Fedrizzi, B.; Gardner, R.C. Influence of Fermentation Temperature, Yeast Strain, and Grape Juice on the Aroma Chemistry and Sensory Profile of Sauvignon Blanc Wines. J. Agric. Food Chem. 2017, 65, 8902–8912. [Google Scholar] [CrossRef]
- Ough, C.S. Fermentation rates of grape juice. I. Effects of temperature and composition of white juice fermentation rates. Am. J. Enol. Vitic. 1964, 15, 167–177. [Google Scholar]
- Beltran, G.; Novo, M.; Leberre, V.; Sokol, S.; Labourdette, D.; Guillamon, J.M.; Mas, A.; Francois, J.; Rozes, N. Integration of transcriptomic and metabolic analyses for understanding the global responses of low-temperature winemaking fermentations. FEMS Yeast Res. 2006, 6, 1167–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Ríos, E.; Morard, M.; Parts, L.; Liti, G.; Guillamón, J.M. The genetic architecture of low-temperature adaptation in the wine yeast Saccharomyces cerevisiae. BMC Genom. 2017, 18, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, J.M.; Gaucher, E.A.; Burgan, M.F.; De Kee, D.W.; Li, T.; Aris, J.P.; Benner, S.A. Resurrecting ancestral alcohol dehydrogenases from yeast. Nat. Genet. 2005, 37, 630–635. [Google Scholar] [CrossRef]
- Gordon, J.L.; Byrne, K.P.; Wolfe, K.H. Additions, losses, and rearrangements on the evolutionary route from a reconstructed ancestor to the modern Saccharomyces cerevisiae genome. PLoS Genet. 2009, 5, e1000485. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, T.; Postaire, O.; Storai, J.; Blondin, B. Analysis of the genomic response of a wine yeast to rehydration and inoculation. Appl. Microbiol. Biotechnol. 2006, 71, 699–712. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Beltran, G.; Warringer, J.; Guillamón, J.M. Genetic basis of variations in nitrogen source utilization in four wine commercial yeast strains. PLoS ONE 2013, 8, e67166. [Google Scholar] [CrossRef] [Green Version]
- Treu, L.; Campanaro, S.; Nadai, C.; Toniolo, C.; Nardi, T.; Giacomini, A.; Valle, G.; Blondin, B.; Corich, V. Oxidative stress response and nitrogen utilization are strongly variable in Saccharomyces cerevisiae wine strains with different fermentation performances. Appl. Microbiol. Biotechnol. 2014, 98, 4119–4135. [Google Scholar] [CrossRef]
- Groat, M.; Ough, C.S. Effects of Insoluble Solids Added to Clarified Musts on Fermentation Rate, Wine Composition, and Wine Quality. Am. J. Enol. Vitic. 1978, 29, 112–119. [Google Scholar]
- Marks, V.D.; Ho Sui, S.J.; Erasmus, D.; Van Der Merwe, G.K.; Brumm, J.; Wasserman, W.W.; Bryan, J.; Van Vuuren, H.J.J. Dynamics of the yeast transcriptome during wine fermentation reveals a novel fermentation stress response. FEMS Yeast Res. 2008, 8, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Gasch, A.P.; Werner-Washburne, M. The genomics of yeast responses to environmental stress and starvation. Funct. Integr. Genom. 2002, 2, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Cliff, M.; Girard, B.; Kopp, T.G. Influence of fermentation temperature on composition and sensory properties of Sémillon and Shiraz wines. Am. J. Enol. Vitic. 2001, 52, 235–240. [Google Scholar]
- Salvadó, Z.; Arroyo-López, F.N.; Guillamón, J.M.; Salazar, G.; Quero, A.; Barrio, E. Temperature adaptation markedly determines evolution within the genus Saccharomyces. Appl. Environ. Microbiol. 2011, 77, 2292–2302. [Google Scholar] [CrossRef] [Green Version]
- Cromie, G.A.; Hyma, K.E.; Ludlow, C.L.; Garmendia-Torres, C.; Gilbert, T.L.; May, P.; Huang, A.A.; Dudley, A.M.; Fay, J.C. Genomic sequence diversity and population structure of Saccharomyces cerevisiae assessed by RAD-seq. G3 (Bethesda) 2013, 3, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Borneman, A.R.; Forgan, A.H.; Pretorius, I.S.; Chambers, P.J. Comparative genome analysis of a Saccharomyces cerevisiae wine strain. FEMS Yeast Res. 2008, 8, 1185–1195. [Google Scholar] [CrossRef] [Green Version]
- Ivorra, C.; Perez-Ortin, J.E.; Del Olmo, M. An inverse correlation between stress resistance and stuck fermentations in wine yeasts. A molecular study. Biotechnol. Bioeng. 1999, 64, 698–708. [Google Scholar] [CrossRef]
- Berthels, N.J.; Cordero Otero, R.R.; Bauer, F.F.; Pretorius, I.S.; Thevelein, J.M. Correlation between glucose/fructose discrepancy and hexokinase kinetic properties in different Saccharomyces cerevisiae wine yeast strains. Appl. Microbiol. Biotechnol. 2007, 77, 1083–1091. [Google Scholar] [CrossRef]
- Santos, J.; Sousa, M.J.; Cardoso, H.; Inacio, J.; Silva, S.; Spencer-Martins, I.; Leao, C. Ethanol tolerance of sugar transport, and the rectification of stuck wine fermentations. Microbiology 2008, 154, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Harsch, M.J.; Lee, S.A.; Goddard, M.R.; Gardner, R.C. Optimized fermentation of grape juice by laboratory strains of Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro, F.; Vargas, F.A.; Agosin, E. A systems biology perspective of wine fermentations. Yeast 2007, 24, 977–991. [Google Scholar] [CrossRef] [PubMed]
- García-Ríos, E.; Ramos-Alonso, L.; Guillamón, J.M. Correlation between low temperature adaptation and oxidative stress in Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strope, P.K.; Skelly, D.A.; Kozmin, S.G.; Mahadevan, G.; Stone, E.A.; Magwene, P.M.; Dietrich, F.S.; McCusker, J.H. The 100-genomes strains, an S. cerevisiae resource that illuminates its natural phenotypic and genotypic variation and emergence as an opportunistic pathogen. Genome Res. 2015, 125, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Kong, I.I.; Turner, T.L.; Kim, H.; Kim, S.R.; Jin, Y.S. Phenotypic evaluation and characterization of 21 industrial Saccharomyces cerevisiae yeast strains. FEMS Yeast Res. 2018, 18, foy001. [Google Scholar] [CrossRef]
- Perez-Ortin, J.E.; Querol, A.; Puig, S.; Barrio, E. Molecular characterization of a chromosomal rearrangement involved in the adaptive evolution of yeast strains. Genome Res. 2002, 12, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Will, J.L.; Kim, H.S.; Clarke, J.; Painter, J.C.; Fay, J.C.; Gasch, A.P. Incipient balancing selection through adaptive loss of aquaporins in natural Saccharomyces cerevisiae populations. PLoS Genet. 2010, 6, e1000893. [Google Scholar] [CrossRef] [Green Version]
- MacLean, R.C.; Gudelj, I. Resource competition and social conflict in experimental populations of yeast. Nature 2006, 441, 498–501. [Google Scholar] [CrossRef]
- Albertin, W.; Marullo, P.; Aigle, M.; Dillmann, C.; De Vienne, D.; Bely, M.; Sicard, D. Population size drives industrial Saccharomyces cerevisiae alcoholic fermentation and is under genetic control. Appl. Environ. Microbiol. 2011, 77, 2772–2784. [Google Scholar] [CrossRef] [Green Version]
- Brice, C.; Cubillos, F.A.; Dequin, S.; Camarasa, C.; Martínez, C. Adaptability of the Saccharomyces cerevisiae yeasts to wine fermentation conditions relies on their strong ability to consume nitrogen. PLoS ONE 2018, 13, e0192383. [Google Scholar] [CrossRef] [Green Version]
- Almeida, P.; Barbosa, R.; Zalar, P.; Imanishi, Y.; Shimizu, K.; Turchetti, B.; Legras, J.L.; Serra, M.; Dequin, S.; Couloux, A.; et al. A population genomics insight into the Mediterranean origins of wine yeast domestication. Mol. Ecol. 2015, 24, 5412–5427. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; Forgan, A.H.; Kolouchova, R.; Fraser, J.A.; Schmidt, S.A. Whole genome comparison reveals high levels of inbreeding and strain redundancy across the spectrum of commercial wine strains of Saccharomyces cerevisiae. G3 (Bethesda) 2016, 6, 957–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldarov, M.A.; Kishkovskaia, S.A.; Tanaschuk, T.N.; Mardanov, A.V. Genomics and biochemistry of Saccharomyces cerevisiae wine yeast strains. Biochemistry (Moscow) 2016, 81, 1650–1668. [Google Scholar] [CrossRef]
- Fleet, G.H.; Heard, G.M. Yeasts—Growth During Fermentation; Harwood Academic Publishers: Singapore, 1993. [Google Scholar]
- Pretorius, I.S.; Du Toit, M.; Van Rensburg, P. Designer yeasts for the fermentation industry of the 21st century. Food Technol. Biotechnol. 2003, 41, 3–10. [Google Scholar]
- Akao, T. Progress in the genomics and genome-wide study of sake yeast. Biosci. Biotechnol. Biochem. 2019, 83, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Salinas, F.; Cubillos, F.A.; Soto, D.; Garcia, V.; Bergström, A.; Warringer, J.; Ganga, M.A.; Louis, E.J.; Liti, G.; Martinez, C. The genetic basis of natural variation in oenological traits in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e49640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazir, M.H.M.; Kadir, R.A.; Gloor, E.; Galbraith, D. Effect of agroclimatic variability on land suitability for cultivating rubber (Hevea brasiliensis) and growth performance assessment in the tropical rainforest climate of Peninsular Malaysia. Clim. Risk Manag. 2020, 27, 100203. [Google Scholar] [CrossRef]
- Sampaio, J.P.; Gonçalves, P. Biogeography and ecology of the genus Saccharomyces. In Yeasts in Natural Ecosystems: Ecology; Springer: Cham, Switzerland, 2017; pp. 131–153. [Google Scholar]
- Spor, A.; Wang, S.; Dillmann, C.; De Vienne, D.; Sicard, D. “Ant” and “grasshopper” life-history strategies in Saccharomyces cerevisiae. PLoS ONE 2008, 3, e1579. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deed, R.C.; Pilkington, L.I. Lifestyle, Lineage, and Geographical Origin Influence Temperature-Dependent Phenotypic Variation across Yeast Strains during Wine Fermentation. Microorganisms 2020, 8, 1367. https://doi.org/10.3390/microorganisms8091367
Deed RC, Pilkington LI. Lifestyle, Lineage, and Geographical Origin Influence Temperature-Dependent Phenotypic Variation across Yeast Strains during Wine Fermentation. Microorganisms. 2020; 8(9):1367. https://doi.org/10.3390/microorganisms8091367
Chicago/Turabian StyleDeed, Rebecca C., and Lisa I. Pilkington. 2020. "Lifestyle, Lineage, and Geographical Origin Influence Temperature-Dependent Phenotypic Variation across Yeast Strains during Wine Fermentation" Microorganisms 8, no. 9: 1367. https://doi.org/10.3390/microorganisms8091367