Identification Mycobacterium spp. in the Natural Water of Two Austrian Rivers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of the Water Samples
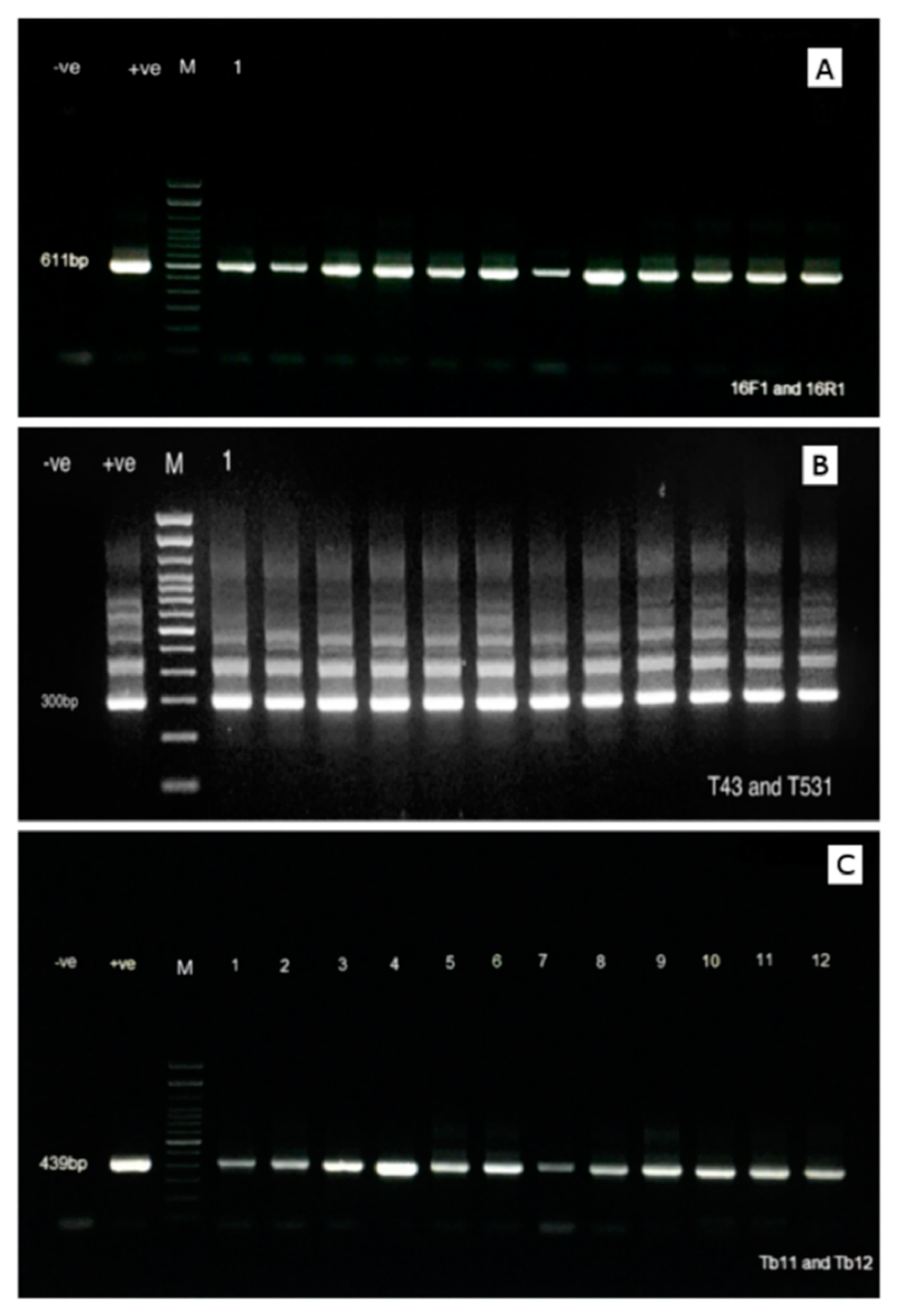
2.2. PCR Assay for the 16S rRNA and hsp65 Genes
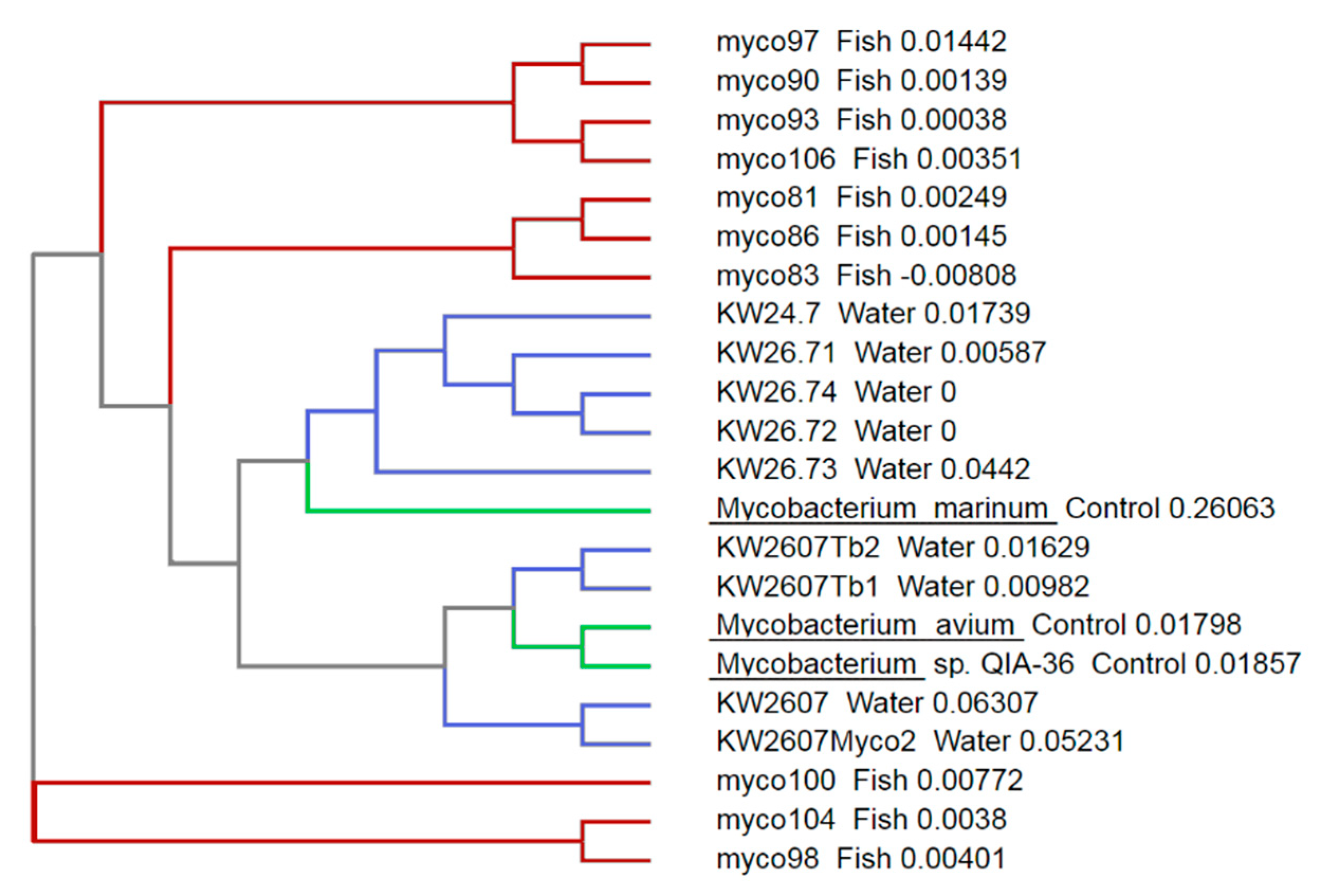
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NTM | Nontuberculous mycobacteria |
| eDNA | Environmental DNA |
| IMS-PCR | Immunomagnetic separation polymerase chain reaction |
| nPCR | Nested PCR |
| BLAST | Basic Local Alignment Sequence Tool |
| HEYM | Herrold’s egg yolk medium |
| IMS-PCR | Immunomagnetic separation polymerase chain reaction |
| PPE proteins | Proline-Proline-Glutamic Acid proteins |
References
- Chinabut, S. Fish Disease and Disorders: Viral, Bacterial, and Fungal Infections; CAB International: Wallingford, UK, 1999; pp. 319–340. [Google Scholar]
- Akbari, S.; Mosavari, N.; Tadayon, K.; Rahmati-Holasoo, H. Isolation of Mycobacterium fortuitum from fish tanks in Alborz, Iran. Iran. J. Microbiol. 2014, 6, 234–239. [Google Scholar] [PubMed]
- Johansen, M.D.; Herrmann, J.-L.; Kremer, L. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat. Rev. Genet. 2020, 18, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Schulze-Robbecke, R.; Hagenau, C.; Behringer, K. Comparison of methods for isolation of mycobacteria from water. Appl. Environ. Microbiol. 1997, 63, 547–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dailloux, M.; Laurain, C.; Weber, M.; Hartemann, P. Water and nontuberculous mycobacteria. Water Res. 1999, 33, 2219–2228. [Google Scholar] [CrossRef]
- Elliot, D.G. Fish Viruses and Bacteria: Pathobiology and Protection; CABI Publishing: Wallingford, UK, 2017; pp. 286–297. [Google Scholar]
- Goodfellow, M.; Magee, J.G. Taxonomy of Mycobacteria. In Mycobacteria; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1998; pp. 1–71. [Google Scholar]
- Hruska, K.; Kaevska, M. Mycobacteria in water, soil, plants and air: A review. Veterinární Medicína 2013, 57, 623–679. [Google Scholar] [CrossRef] [Green Version]
- Falkinham, J.O. Impact of human activities on the ecology of nontuberculous mycobacteria. Futur. Microbiol. 2010, 5, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Beran, V.; Matlova, L.; Dvorska, L.; Svastova, P.; Pavlik, I. Distribution of mycobacteria in clinically healthy ornamental fish and their aquarium environment. J. Fish Dis. 2006, 29, 383–393. [Google Scholar] [CrossRef]
- Slany, M.; Makovcova, J.; Jezek, P.; Bodnarova, M.; Pavlik, I. Relative prevalence of Mycobacterium marinum in fish collected from aquaria and natural freshwaters in central Europe. J. Fish Dis. 2014, 37, 527–533. [Google Scholar] [CrossRef]
- Dowdell, K.; Haig, S.J.; Caverly, L.J.; Shen, Y.; Lipuma, J.J.; Raskin, L. Nontuberculous mycobacteria in drinking water systems—The challenges of characterization and risk mitigation. Curr. Opin. Biotechnol. 2019, 57, 127–136. [Google Scholar] [CrossRef]
- Ghazesaeed, K.; Mohammadi, M. Study of different types of mycobacteria in sediments of fish breeding pools of north of Iran. Tehran Univ. Med. J. TUMS Publ. 1997, 55, 45–49. [Google Scholar]
- Francis-Floyd, R. Mycobacterial Infections of Fish; Southern Regional Aquaculture Center: Stoneville, MS, USA, 2011; pp. 1–12. [Google Scholar]
- Novotny, L.; Halouzka, R.; Matlova, L.; Vavra, O.; Bartosova, L.; Slany, M.; Pavlik, I. Morphology and distribution of granulomatous inflammation in freshwater ornamental fish infected with mycobacteria. J. Fish Dis. 2010, 33, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Puk, K.; Banach, T.; Wawrzyniak, A.; Ziętek, J.A.; Winiarczyk, S. Detection of Mycobacterium marinum, M. peregrinum, M. fortuitum and M. abscessus in aquarium fish. J. Fish Dis. 2018, 41, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, D.T.; Rhodes, M.W. Mycobacteriosis in fishes: A review. Veter J. 2009, 180, 33–47. [Google Scholar] [CrossRef]
- Dousa, K.M.; Babiker, A.; Van Aartsen, D.; Shah, N.; A Bonomo, R.; Johnson, J.L.; Skalweit, M.J. Ibrutinib Therapy and Mycobacterium chelonae Skin and Soft Tissue Infection. Open Forum Infect. Dis. 2018, 5, 168. [Google Scholar] [CrossRef] [Green Version]
- Rudra, P.; Rajesh, S.; Pritam, N. Pulmonary Infection with Mycobacterium Gordonae in an Immunocompetent Patient: A Case Report. Am. J. Infect. Dis. Microbiol. 2013, 2, 1–3. [Google Scholar] [CrossRef]
- Rotman, D.A.; Blauvelt, A.; Kerdel, F.A. Widespread primary cutaneous infection with mycobacterium fortuitum. Int. J. Dermatol. 1993, 32, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.M.; Chng, H.H. Disseminated Mycobacterium flavescens in a probable case of chronic granulomatous disease. J. Infect. 1993, 26, 83–86. [Google Scholar] [CrossRef]
- Aubry, A.; Mougari, F.; Reibel, F.; Cambau, E. Mycobacterium Marinum. Tuberc. Nontuberculous Mycobact. Infect. 2017, 5, 735–752. [Google Scholar] [CrossRef]
- Hashish, E.A.; Merwad, A.-R.M.; Elgaml, S.; Amer, A.; Kamal, H.; Elsadek, A.; Marei, A.; Sitohy, M. Mycobacterium marinum infection in fish and man: Epidemiology, pathophysiology and management; a review. Vet. Q. 2018, 38, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Diaz, E.; Morfin-Otero, R.; Perez-Gomez, H.R.; Esparza-Ahumada, S.; Rodriguez-Noriega, E. Rapidly Growing Mycobacterial Infections of the Skin and Soft Tissues Caused by M. fortuitum and M. chelonae. Curr. Trop. Med. Rep. 2018, 5, 162–169. [Google Scholar] [CrossRef]
- Lescenko, P.; Mátlová, L.; Dvorská, L.; Bartos, M.; Vavra, O.; Navratil, S.; Novotny, L.; Pavlik, I. Mycobacterial infection in aquarium fish. Veterinární Medicína 2012, 48, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Mrlik, V.; Slany, M.; Kubečka, J.; Seda, J.; Necas, A.; Babák, V.; Slana, I.; Kriz, P.; Pavlik, I. A low prevalence of mycobacteria in freshwater fish from water reservoirs, ponds and farms. J. Fish Dis. 2012, 35, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, R.G.; Florio, D.; Fioravanti, M.L.; Rossi, M.; Prearo, M. Occurrence of Mycobacterium spp. in ornamental fish in Italy. J. Fish Dis. 2008, 31, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Griffiths, J.; Mitchell, C.; Wood, B.; Fletcher, Z.; Drobniewski, F.; Hastings, T. Pathology attributed to Mycobacterium chelonae infection among farmed and laboratory-infected Atlantic salmon Salmo salar. Dis. Aquat. Org. 1998, 33, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Strike, T.B.; Feltrer, Y.; Flach, E.; MacGregor, S.K.; Guillaume, S. Investigation and management of an outbreak of multispecies mycobacteriosis in Australian lungfish (Neoceratodus fosteri) including the use of triple antibiotic treatment. J. Fish Dis. 2016, 40, 557–570. [Google Scholar] [CrossRef]
- Pate, M.; Jencic, V.; Zolnir-Dovc, M.; Ocepek, M. Detection of mycobacteria in aquarium fish in Slovenia by culture and molecular methods. Dis. Aquat. Org. 2005, 64, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J.; Stine, C.B.; Baya, A.M.; Kent, M.L. A review of mycobacteriosis in marine fish. J. Fish Dis. 2009, 32, 119–130. [Google Scholar] [CrossRef]
- Pradhan, G.; Shrivastva, R.; Mukhopadhyay, S. Mycobacterial PknG Targets the Rab7l1 Signaling Pathway To Inhibit Phagosome–Lysosome Fusion. J. Immunol. 2018, 201, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- Blaas, K. Aquaculture 2020—Austrian Strategy to Increase the National Fish Production. 2012. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&cad=rja&uact=8&ved=2ahUKEwiujP_TmobfAhVyh4sKHeP6CboQFjAAegQICRAC&url=https%253A%252F%252Fwww.bmnt.gv.at%252Fdam%252Fjcr%253A19848c67-c1f8-406e-a3d5-186a977ece5f%252FAquaculture%2525202020%252520-%252520Austrian%25 (accessed on 26 August 2020).
- Delghandi, M.R.; Menanteau-Ledouble, S.; Waldner, K.; El-Matbouli, M. Renibacterium salmoninarum and Mycobacterium spp.: Two bacterial pathogens present at low levels in wild brown trout (Salmo trutta fario) populations in Austrian rivers. BMC Vet. Res. 2020, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Waldner, K.; Bechter, T.; Auer, S.; Borgwardt, F.; El-Matbouli, M.; Unfer, G. A brown trout (Salmo trutta) population faces devastating consequences due to proliferative kidney disease and temperature increase: A case study from Austria. Ecol. Freshw. Fish 2019, 29, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, P.R.; Sepulveda, A.J.; Martin, R.M.; Hopper, L.R. A probe-based quantitative PCR assay for detecting Tetracapsuloides bryosalmonae in fish tissue and environmental DNA water samples. Conserv. Genet. Resour. 2017, 10, 317–319. [Google Scholar] [CrossRef]
- Talaat, A.M.; Reimschuessel, R.; Trucksis, M. Identification of mycobacteria infecting fish to the species level using polymerase chain reaction and restriction enzyme analysis. Vet. Microbiol. 1997, 58, 229–237. [Google Scholar] [CrossRef]
- Telenti, A.; Marchesi, F.; Balz, M.; Bally, F.; Böttger, E.C.; Bodmer, T. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol. 1993, 31, 175–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickup, R.W.; Rhodes, G.; Arnott, S.; Sidi-Boumedine, K.; Bull, T.J.; Weightman, A.J.; Hurley, M.; Hermon-Taylor, J. Mycobacterium avium subsp. paratuberculosis in the Catchment Area and Water of the River Taff in South Wales, United Kingdom, and Its Potential Relationship to Clustering of Crohn’s Disease Cases in the City of Cardiff. Appl. Environ. Microbiol. 2005, 71, 2130–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartram, J.; Dufour, A. Pathogenic Mycobacteria in Water: A Guide to Public Health Consequences, Monitoring and Management; World Health Organization: Geneva, Switzerland; IWA Publishing: London, UK, 2004. [Google Scholar]
- Peters, M.; Muller, C.; Rüsch-Gerdes, S.; Seidel, C.; Gobel, U.; Pohle, H.; Ruf, B. Isolation of atypical mycobacteria from tap water in hospitals and homes: Is this a possible source of disseminated MAC infection in AIDS patients? J. Infect. 1995, 31, 39–44. [Google Scholar] [CrossRef]
- Le Dantec, C.; Duguet, J.-P.; Montiel, A.; Dumoutier, N.; Dubrou, S.; Vincent, V. Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems. Appl. Environ. Microbiol. 2002, 68, 5318–5325. [Google Scholar] [CrossRef] [Green Version]
- Chilima, B.Z.; Clark, I.M.; Floyd, S.; Fine, P.E.M.; Hirsch, P. Distribution of Environmental Mycobacteria in Karonga District, Northern Malawi. Appl. Environ. Microbiol. 2006, 72, 2343–2350. [Google Scholar] [CrossRef] [Green Version]
- Salogni, C.; Zanoni, M.; Covi, M.; Pacciarini, M.L.; Alborali, G.L. Infezione da Mycobacterium marinum: Descrizione di un focolaio di malattia in trota iridea (Oncorhynchus mykiss) e trota fario (Salmo trutta) d’allevamento. Ittiopatologia 2007, 4, 227–237. [Google Scholar]
- Kušar, D.; Zajc, U.; Jenčič, V.; Ocepek, M.; Higgins, J.; Žolnir-Dovč, M.; Pate, M. Mycobacteria in aquarium fish: Results of a 3-year survey indicate caution required in handling pet-shop fish. J. Fish Dis. 2016, 40, 773–784. [Google Scholar] [CrossRef]
- Niva, M.; Hernesmaa, A.; Haahtela, K.S.S.M.; Sivonen, K.H.K. Actinobacterial communities of boreal forest soil and lake water are rich in mycobacteria. Boreal. Environ. Res. 2006, 11, 45–53. [Google Scholar]
- Gauthier, D.T.; Reece, K.S.; Xiao, J.; Rhodes, M.W.; Kator, H.I.; Latour, R.J.; Bonzek, C.F.; Hoenig, J.M.; Vogelbein, W.K. Quantitative PCR Assay for Mycobacterium pseudoshottsii and Mycobacterium shottsii and Application to Environmental Samples and Fishes from the Chesapeake Bay. Appl. Environ. Microbiol. 2010, 76, 6171–6179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahbar, M.; Lamei, A.; Babazadeh, H.; Yavari, S.A. Isolation of rapid growing mycobacteria from soil and water in Iran. Afr. J. Biotechnol. 2010, 9, 3618–3621. [Google Scholar]
- Slany, M.; Jezek, P.; Fiserova, V.; Bodnarova, M.; Štork, J.; Havelkova, M.; Kalat, F.; Pavlik, I. Mycobacterium marinum infections in humans and tracing of its possible environmental sources. Can. J. Microbiol. 2012, 58, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, J.L. Mycobacteriosis in Striped Bass, Morone Saxatilis, from Virginia Waters of Chesapeake Bay. Master’s Thesis, The Collage of William and Mary, Williamsburg, VA, USA, 2001. [Google Scholar]
- Josephson, K.L.; Gerba, C.P.; Pepper, I.L. Polymerase chain reaction detection of nonviable bacterial pathogens. Appl. Environ. Microbiol. 1993, 59, 3513–3515. [Google Scholar] [CrossRef] [Green Version]
- Whan, L.; Ball, H.J.; Grant, I.R.; Rowe, M.T. Development of an IMS–PCR assay for the detection of Mycobacterium avium subsp. paratuberculosis in water. Lett. Appl. Microbiol. 2005, 40, 269–273. [Google Scholar] [CrossRef]
- Whan, L.; Ball, H.J.; Grant, I.R.; Rowe, M.T. Occurrence of Mycobacterium avium subsp. paratuberculosis in Untreated Water in Northern Ireland. Appl. Environ. Microbiol. 2005, 71, 7107–7112. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| River Sites | Sampling | Number of Positive/Prevalence Rate of Mycobacterium sp. | |
|---|---|---|---|
| Date | Number | ||
| Kamp | June 2018 | 25 | 25/25 (100%) |
| January 2019 | 20 | 20/20 (100%) | |
| Wulka | July 2018 | 10 | 10/10 (100%) |
| January 2019 | 15 | 11/15 (73.33%) | |
| Totals | Total for 2018 | 35 | 35/35 (100%) |
| Total for 2019 | 35 | 31/35 (88.57%) | |
| Both years | 70 | 66/70 (94.28%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delghandi, M.R.; Waldner, K.; El-Matbouli, M.; Menanteau-Ledouble, S. Identification Mycobacterium spp. in the Natural Water of Two Austrian Rivers. Microorganisms 2020, 8, 1305. https://doi.org/10.3390/microorganisms8091305
Delghandi MR, Waldner K, El-Matbouli M, Menanteau-Ledouble S. Identification Mycobacterium spp. in the Natural Water of Two Austrian Rivers. Microorganisms. 2020; 8(9):1305. https://doi.org/10.3390/microorganisms8091305
Chicago/Turabian StyleDelghandi, Mohammad Reza, Karoline Waldner, Mansour El-Matbouli, and Simon Menanteau-Ledouble. 2020. "Identification Mycobacterium spp. in the Natural Water of Two Austrian Rivers" Microorganisms 8, no. 9: 1305. https://doi.org/10.3390/microorganisms8091305