Bacillus telluris sp. nov. Isolated from Greenhouse Soil in Beijing, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, and Cultivation
2.2. Phenotypic Characterization
2.3. Chemotaxonomic Analysis
2.4. Phylogenetic 16S rRNA Gene Analysis
2.5. Complete Genome Sequencing and Analysis
2.6. Analysis of Core Orthologous Genes
2.7. Plant Growth-Promoting Characteristics
3. Results and Discussion
3.1. Phenotypic Characterization of 03113T
3.2. Analysis of Isoprenoid Quinones, Polar Lipids, and Cellular Fatty Acids
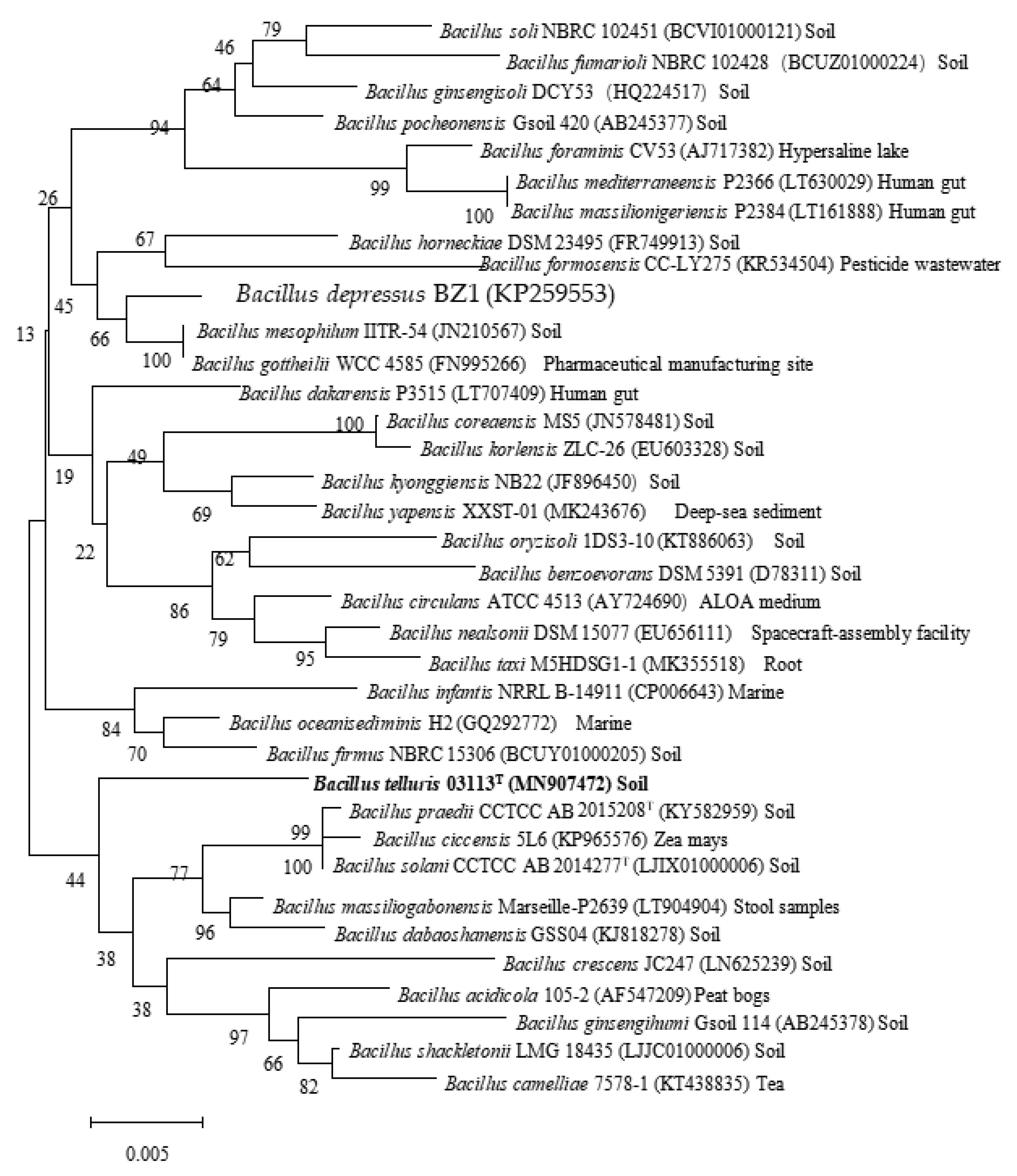
3.3. Phylogenetic Analysis of 16S rRNA
3.4. Whole-Genome Analysis
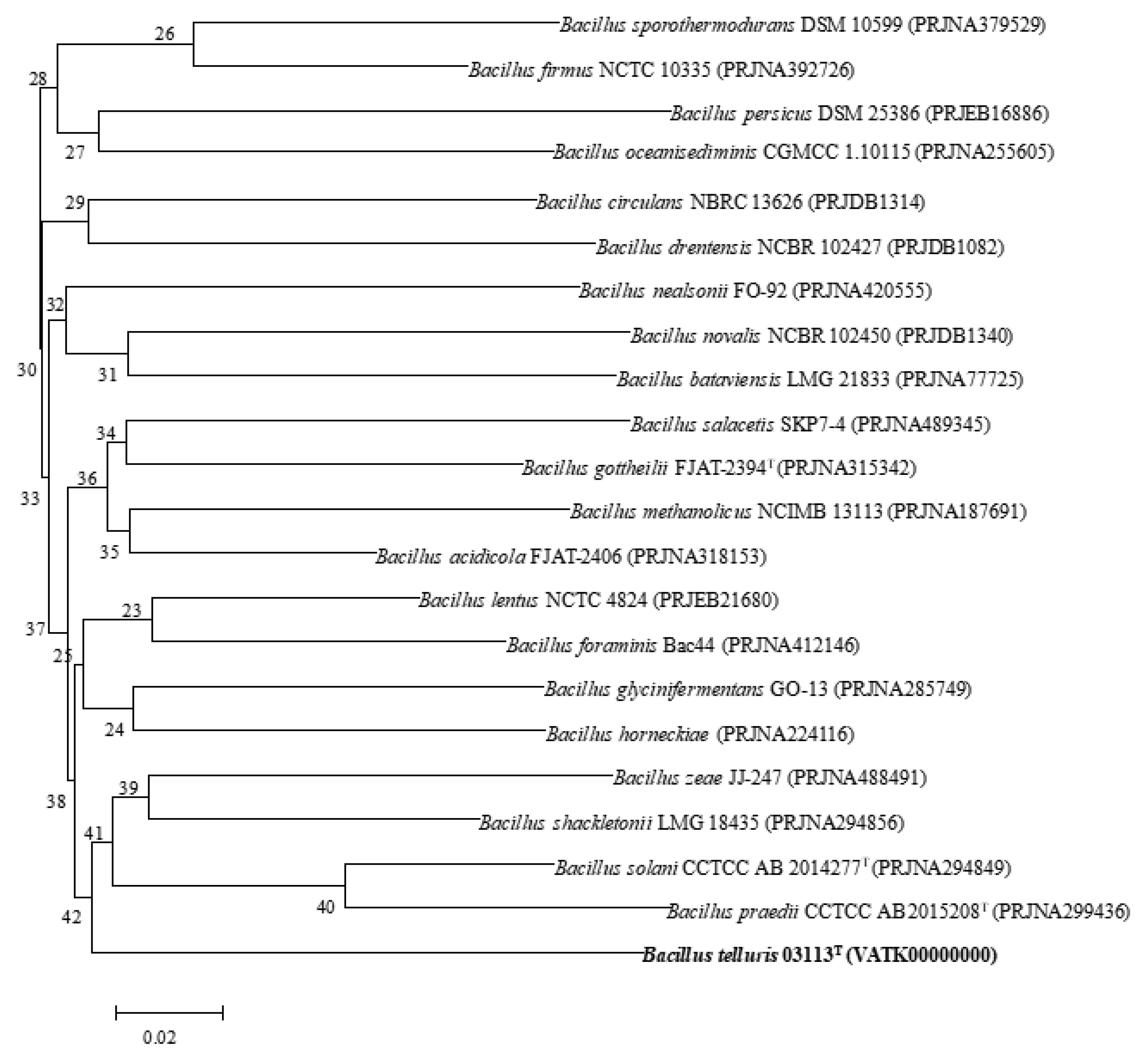
3.5. Phylogenomic Comparative Analysis of Bacillus species
3.6. Plant Growth-Promoting Characteristics of Isolates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A
References
- McKillip, J.L. Prevalence and expression of enterotoxins in Bacillus cereus and other Bacillus spp., a literature review. Antonie Leeuwenhoek 2000, 77, 393–399. [Google Scholar] [CrossRef]
- Claus, D.; Berkeley, R.C. Genus Bacillus. In Bergey’s Manual of Systematic Bacteriology; Wiley: Hoboken, NJ, USA, 1986; pp. 1105–1139. [Google Scholar]
- Alcaraz, L.D.; Moreno-Hagelsieb, G.; Eguiarte, L.E.; Souza, V.; Herrera-Estrella, L.; Olmedo, G. Understanding the evolutionary relationships and major traits of Bacillus through comparative genomics. BMC Genom. 2010, 11, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, N.A.; de Vos, P. Bacillus. In Bergey’s Manual of Systematic Bacteriology: The Firmicutes, 2nd ed.; De Vos, P., Garrity, G., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2009; Volume 3, pp. 21–128. [Google Scholar]
- Read, T.D.; Peterson, S.N.; Tourasse, N.; Baillie, L.W.; Fraser, C.M. The genome sequence of Bacillus anthracis Ames and comparison to closely related bacteria. Nature 2003, 423, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Rosovitz, M.J.; Leppla, S.H. Medicine: Virus deals anthrax a killer blow. Nature 2002, 418, 825–826. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, G.H.; Sengonca, C.; Schumann, P.; Ge, C.B.; Wang, J.P.; Cui, W.D.; Lin, N.Q. Bacillus solani sp. nov. isolated from rhizosphere soil of potato field in Xinjiang of China. Int. J. Syst. Evol. Microbiol. 2015, 65, 4066–4071. [Google Scholar] [CrossRef]
- Liu, B.; Liu, G.H.; Sengonca, C.; Schumann, P.; Wang, M.K. Bacillus praedii sp. nov. isolated from purplish paddy soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2823–2826. [Google Scholar] [CrossRef]
- Meierkolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Markus, G. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar]
- Murray, R.G.; Doetsch, R.N.; Robinow, C.F. Determinative and cytological light microscopy. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology (ASM): Washington, DC, USA, 1994; pp. 21–41. [Google Scholar]
- Chen, Y.G.; Cui, X.L.; Pukall, R.; Li, H.M.; Yang, Y.L.; Xu, L.H.; Wen, M.L.; Peng, Q.; Jiang, C.L. Salinicoccus kunmingensis sp. nov., a moderately halophilic bacterium isolated from a salt mine in Yunnan, south-west China. Int. J. Syst. Evol. Microbiol. 2007, 57, 2327–2332. [Google Scholar] [CrossRef]
- Dong, X.Z.; Cai, M.Y. Determination of Biochemical Properties. In Manual for the Systematic Identification of General Bacteria; Science Press: Beijing, China, 2001; pp. 370–398. [Google Scholar]
- Collins, M.D.; Pirouz, T.; Goodfellow, M.; Minnikin, D.E. Distribution of Menaquinones in Actinomycetes and Corynebacteria. J Gen Microbiol. 1977, 100, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Groth, I.; Schumann, P.; Weiss, N.; Martin, K.; Rainey, F.A. Agrococcus jenensis gen. nov., sp. nov., a new genus of actinomycetes with diaminobutyric acid in the cell wall. Int. J. Syst. Bacteriol. 1996, 46, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Minnikin, D.E.; Collins, M.D.; Goodfellow, M. Fatty acid and polar lipid composition in the classification of Cellulomonas, Oerskovia and related taxa. J. Appl. Bacteriol. 1979, 47, 87–95. [Google Scholar] [CrossRef]
- Sasser, M. Identification of bacteria by gas chromatography of cellular fatty acids. USFCC News. 1990, 20, 16. [Google Scholar]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Rzhetsky, A.; Nei, M. A simple method for estimating and testing minimum evolution trees. Mol. Biol. Evol. 1992, 9, 945–967. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA 4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes. 2016, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2014, 31, 587–589. [Google Scholar] [CrossRef]
- John, B.E.; Bass, L. Usability and software architecture. Behav. Inform. Technol. 2001, 20, 329–338. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Auch, A.F.; von Jan, M.; Klenk, H.P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA—An ultra-fast pan-genome analysis pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Glickmann, E.; Dessaux, Y. Critical Examination of the Specificity of the Salkowski Reagent for Indolic Compounds Produced by Phytopathogenic Bacteria. Appl. Environ. Microbiol. 1995, 61, 793. [Google Scholar] [CrossRef] [Green Version]
- Machuca, A.; Milagres, A.M.F. Use of CAS-agar plate modified to study the effect of different variables on the siderophore production by Aspergillus. Lett. Appl. Microbiol. 2003, 36, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Fankem, H.; Nwaga, D.; Deubel, A.; Dieng, L.; Merbach, W.; Etoa, F.X. Occurrence and functioning of phosphate solubilizing microorganisms from oil palm tree (Elaeis guineensis) rhizosphere in Cameroon. Afr. J. Biotechnol. 2007, 5, 2450–2460. [Google Scholar]
- Kampfer, P. Limits and Possibilities of Total Fatty Acid Analysis for Classification and Identification of Bacillus Species. Syst. Appl. Microbiol. 1994, 17, 86–98. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Y.; Liu, J.; Chang, M.; Zhao, Y.; Zhou, S.; Zhuang, L. Bacillus dabaoshanensis sp. nov., a Cr(VI)-tolerant bacterium isolated from heavy-metal-contaminated soil. Arch Microbiol. 2015, 197, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Hu, G.P.; Tang, W.Q. Characteristic of average nucleotide identity (ANI) based on the whole genomes from Bacillus species in Bacillus-like genus. Fujian J. Agric. Sci. 2013, 28, 833–843. [Google Scholar]
- Li, J.; Gao, R.; Chen, Y.; Xue, D.; Han, J.; Wang, J.; Dai, Q.; Lin, M.; Ke, X.; Zhang, W. Isolation and Identification of Microvirga thermotolerans HR1, a Novel Thermo-Tolerant Bacterium, and Comparative Genomics among Microvirga Species. Microorganisms 2020, 8, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Characteristic | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Optimal growth conditions | ||||
| Temperature for growth (°C) | 37 | 30–37 | 35 | 30 |
| pH for growth | 8.0 | 7.0 | 9.0 | 9.0 |
| NaCl concentration for growth (%, w/v) | 5 | 1 | 0 | 4 |
| The Acid produced from (API 50CH) | ||||
| L-arabinose | + | + | + | − |
| Esculin | + | + | − | − |
| API 20NE | ||||
| β-galactosidase | + | + | − | − |
| Lysine | + | − | − | − |
| Lohn gelatin | + | + | + | − |
| Utilization (Biolog GEN III) | ||||
| dextrin | + | + | − | + |
| d-maltos, d-trehalose, sucrose, d-turanose, d-raffinose | + | − | − | − |
| N-acetyl-d-glucosamine | − | + | + | − |
| N-acetyl-β-d-mannosamine, acetic acid | − | + | − | − |
| Stachyose, d-mannose | w | − | − | − |
| N-acetyl-d-galactosamine | − | w | − | − |
| α-d-glucose | + | − | w | − |
| d-fructose | + | + | w | − |
| Inosine, d-serine, glycerol, d-glucose-6-PO4, d-fructose-6-PO4, nalidixic acid, lithium chloride, aztreonam, lincomycin | − | + | + | + |
| Troleandomycin | − | − | + | − |
| L-aspartic acid, L-glutamic acid, L-histidine, L-pyroglutamic acid, L-serine, L-lactic acid, sodium butyrate | - | + | + | − |
| Fatty acid | 1 | 2a | 3 | 4 |
|---|---|---|---|---|
| C14:0 | 1.96 | 1.1 | 1.77 | 1.68 |
| C16:0 | 7.50 | 2.60 | 1.98 | 2.20 |
| iso-C14:0 | 7.13 | ND | 6.61 | 5.13 |
| iso-C15:0 | 11.06 | 42.9 | 45.43 | 54.12 |
| iso-C16:0 | 8.73 | 6.7 | 6.07 | 5.61 |
| anteiso-C15:0 | 51.35 | 24.1 | 27.16 | 20.15 |
| anteiso-C17:0 | 6.71 | 6.2 | 3.88 | 3.13 |
| C16:1ω7c alcohol | ND | ND | 2.56 | 2.36 |
| C16:1ωw11c | ND | ND | 1.21 | 1.12 |
| Summed Feature 3 * | <1 | 2.5 | ND | ND |
| Summed Feature 8 † | <1 | 1.5 | ND | ND |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.-B.; He, S.-W.; Wang, X.; Thin, K.-K.; Wei, H.-L.; Zhang, X.-X. Bacillus telluris sp. nov. Isolated from Greenhouse Soil in Beijing, China. Microorganisms 2020, 8, 702. https://doi.org/10.3390/microorganisms8050702
Guo H-B, He S-W, Wang X, Thin K-K, Wei H-L, Zhang X-X. Bacillus telluris sp. nov. Isolated from Greenhouse Soil in Beijing, China. Microorganisms. 2020; 8(5):702. https://doi.org/10.3390/microorganisms8050702
Chicago/Turabian StyleGuo, He-Bao, Shan-Wen He, Xing Wang, Kyu-Kyu Thin, Hai-Lei Wei, and Xiao-Xia Zhang. 2020. "Bacillus telluris sp. nov. Isolated from Greenhouse Soil in Beijing, China" Microorganisms 8, no. 5: 702. https://doi.org/10.3390/microorganisms8050702