Differential Analysis of Stress Tolerance and Transcriptome of Probiotic Lacticaseibacillus casei Zhang Produced from Solid-State (SSF-SW) and Liquid-State (LSF-MRS) Fermentations

 ,
, 


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Media, and Growth Conditions
2.2. Preparation of L. casei Zhang Bacterial Suspensions after Solid-State and Liquid-State Fermentation
2.3. Sensitivity of L. casei Zhang to NaCl, H2O2, and Ethanol after Solid-State and Liquid-State Fermentations
2.4. Bacterial Agent Preparation by Cold-Air Drying and Spray Drying
2.5. RNA Extraction, RNA-Seq, Transcriptomic Data Processing, and RNA-Seq Data Accession Number
2.6. Determination of Organic Acids
2.7. Statistical Analysis
3. Results and Discussions
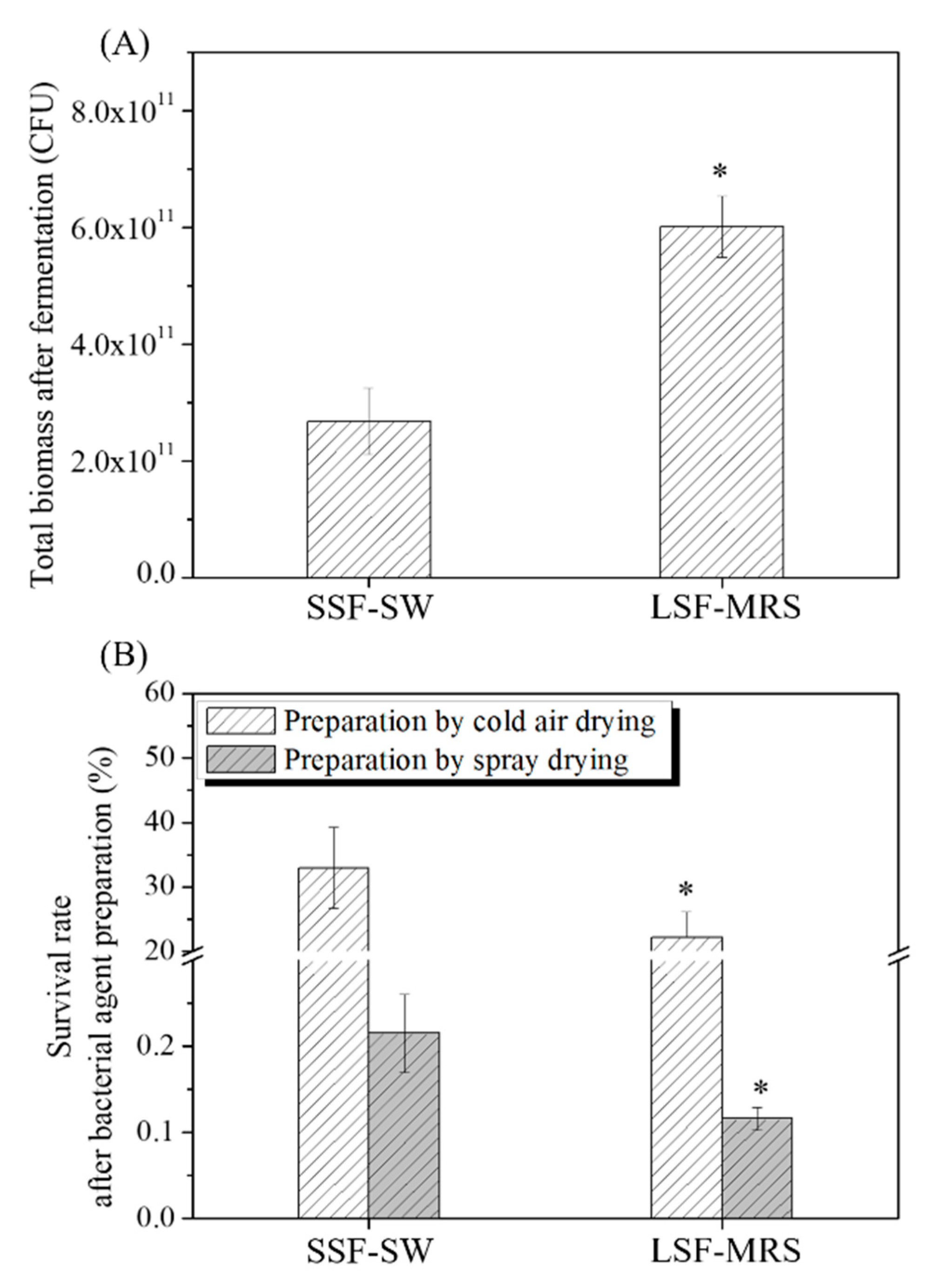
3.1. Total Biomasses of L. casei Zhang after Fermentation with SSF-SW and LSF-MRS Media and Survival Rates of L. casei Zhang after Cold-Air Drying and Spray Drying
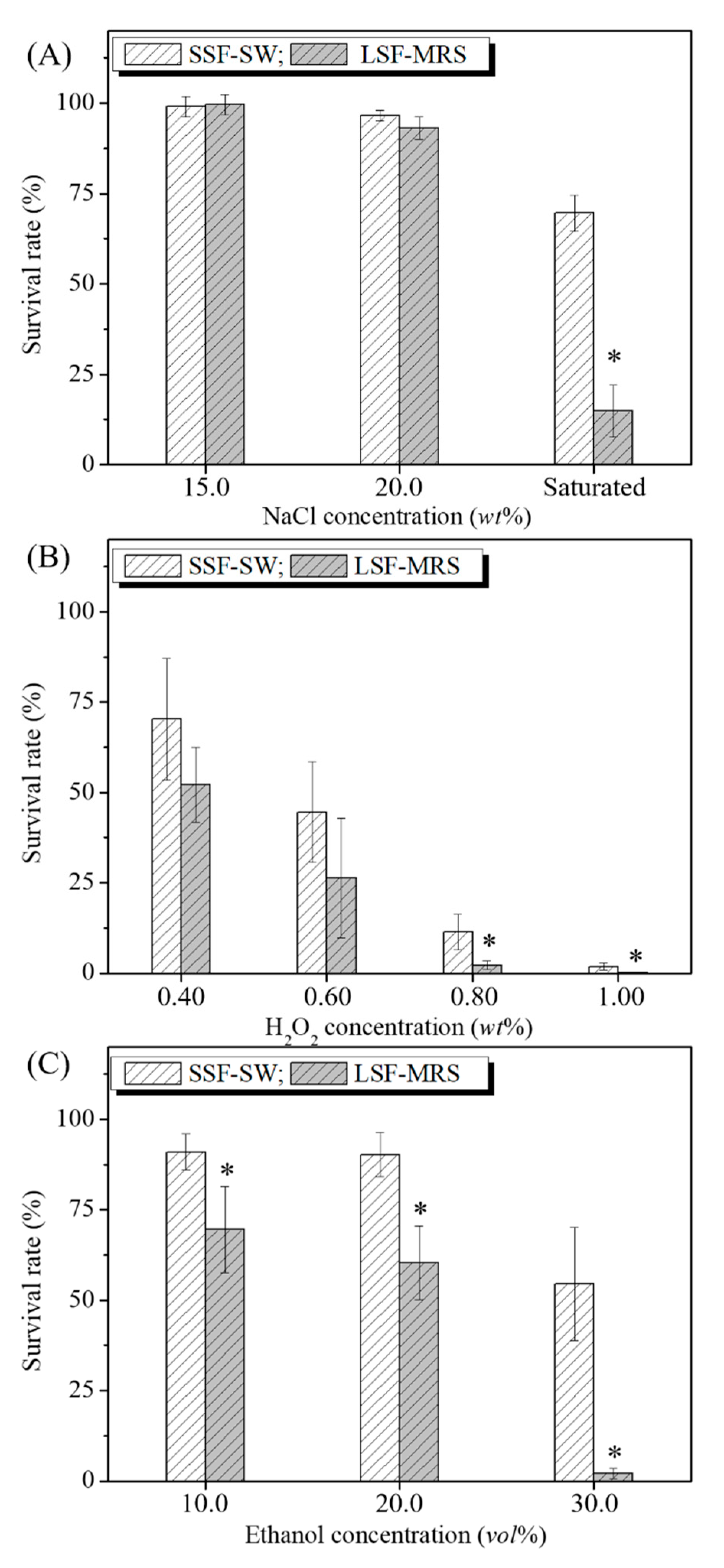
3.2. Effects of NaCl, H2O2, and Ethanol Stresses on the Survival Rates of L. casei Zhang after Fermentation with SSF-SW and LSF-MRS Media
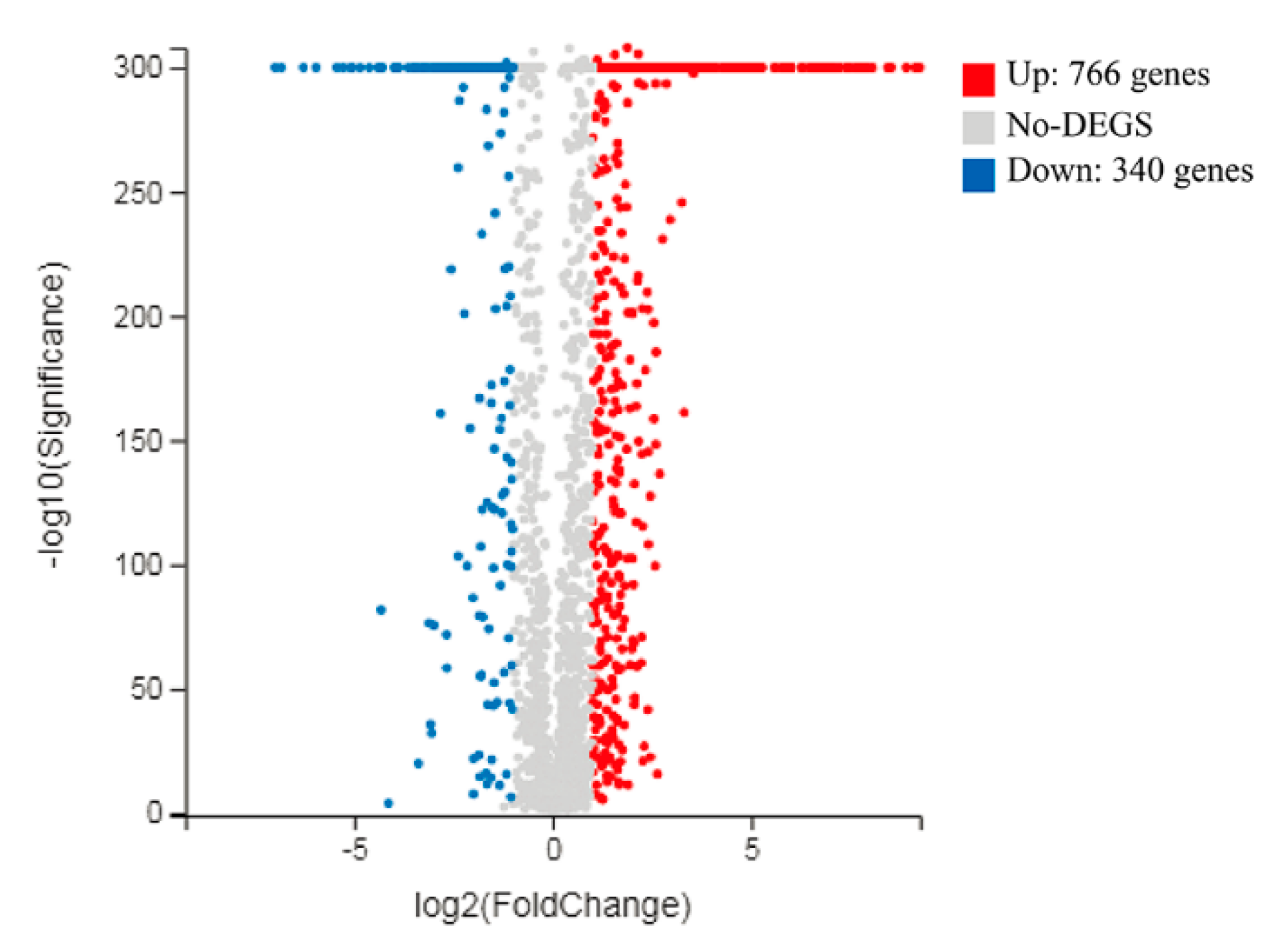
3.3. Overview of L. casei Zhang Transcriptomic Response to Fermentation with SSF-SW and LSF-MRS Media
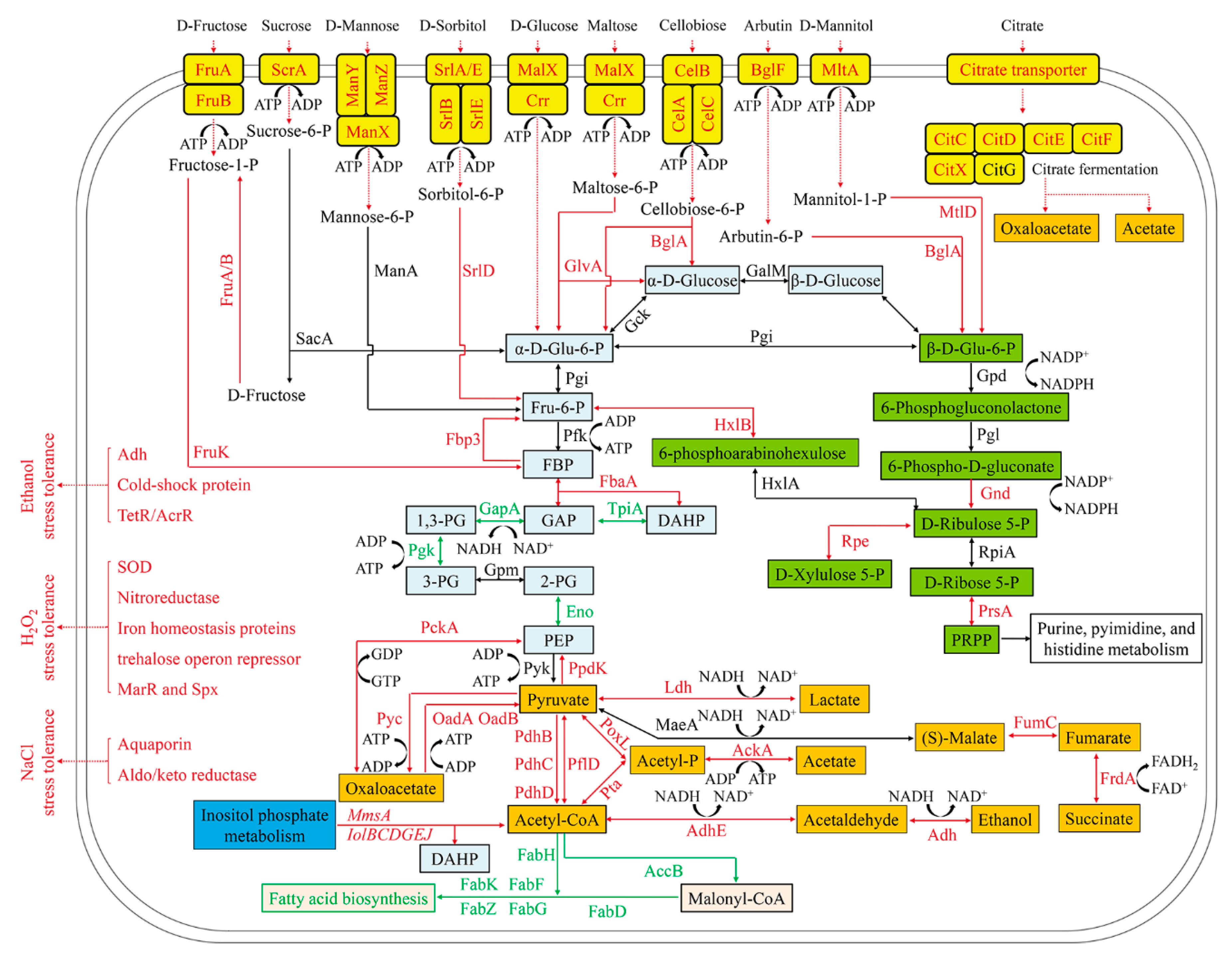
3.4. Enhancement of Carbohydrate Transport in L. casei Zhang Fermented with SSF-SW Medium
3.5. Enhancement of Gluconeogenesis and Suppression of Glycolysis in L. casei Zhang after Fermentation with SSF-SW Medium
3.6. Enhancement of Pentose Phosphate Pathway in L. casei Zhang Cultured with SSF-SW Medium
3.7. Enhancement of Inositol Phosphate Metabolism and Suppression of Fatty Acid Synthesis in L. casei Zhang Cultured with SSF-SW Medium
3.8. Enhancement of Pyruvate Metabolism in L. casei Zhang Cultured with SSF-SW Medium
3.9. Up-Regulation of Stress-Tolerance genes in L. casei Zhang Cultured with SSF-SW Medium
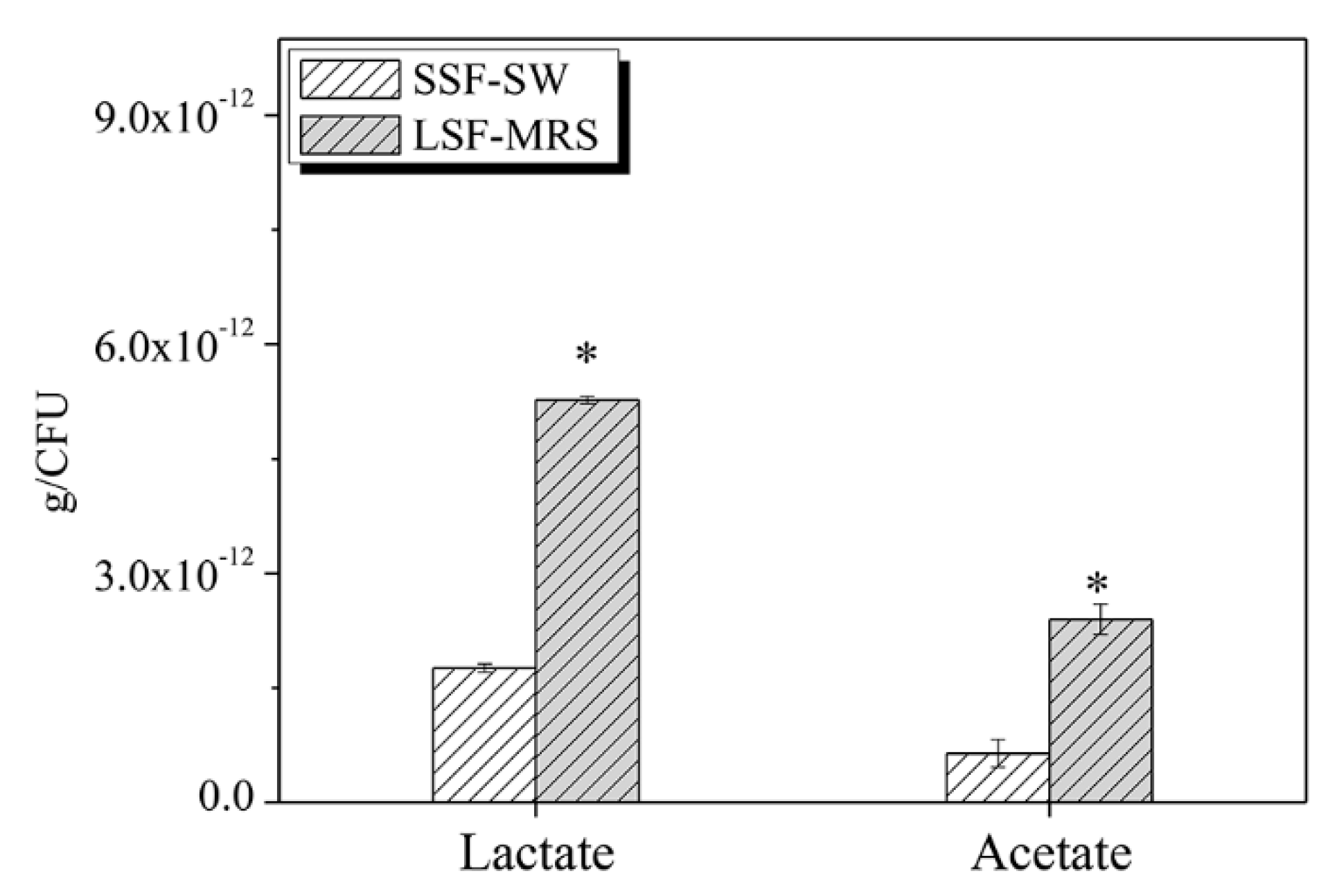
3.10. The Contents of Lactate and Acetate of L. casei Zhang after Fermentation with SSF-SW and LSF-MRS Media
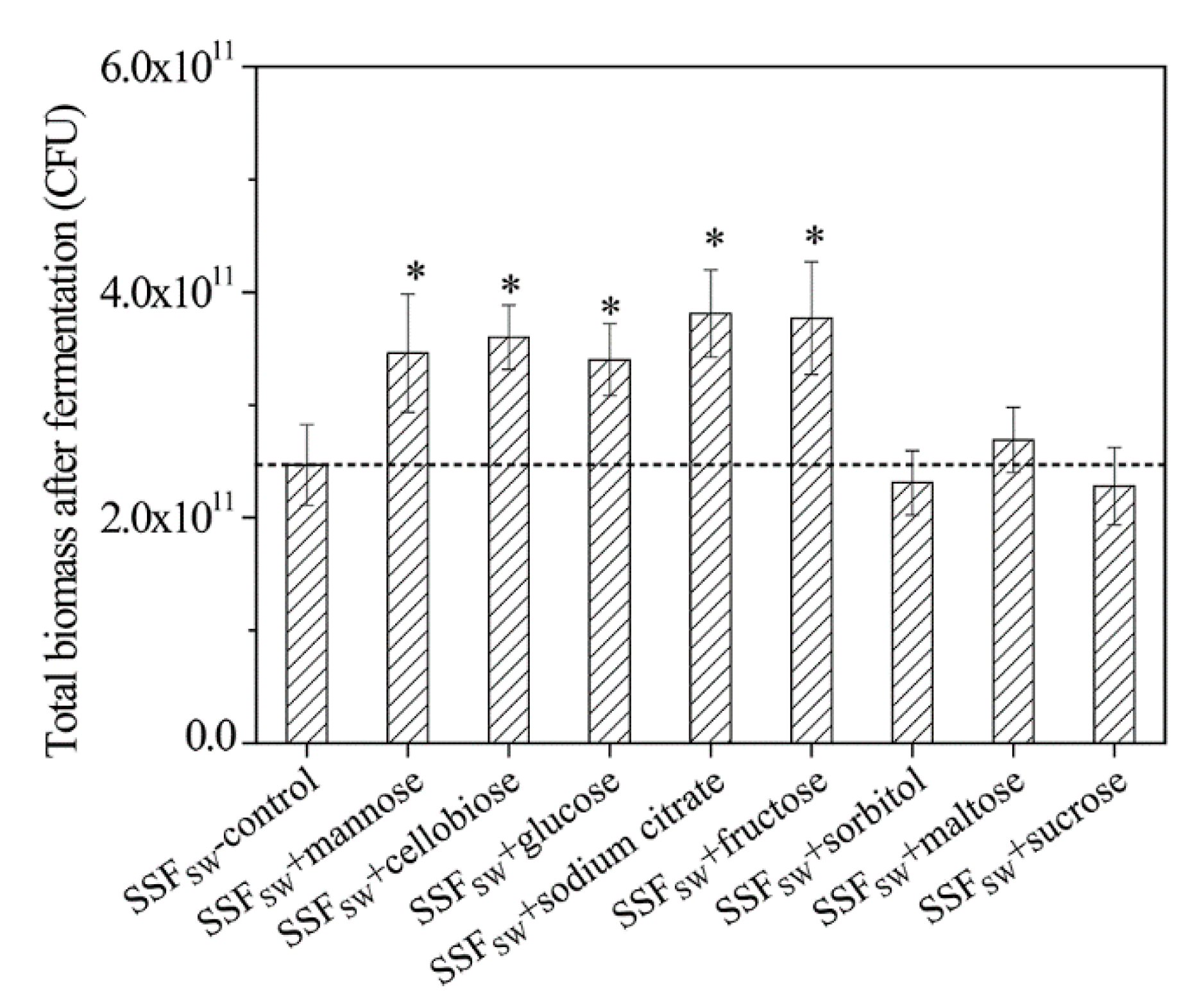
3.11. Total Biomasses of L. casei Zhang after Fermentation with SSF-SW Medium Supplemented with Different Carbon Sources
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nuraida, L. A review: Health promoting lactic acid bacteria in traditional Indonesian fermented foods. Food Sci. Hum. Well. 2015, 4, 47–55. [Google Scholar] [CrossRef] [Green Version]
- García-Burgos, M.; Moreno-Fernández, J.; Alférez, M.J.M.; Díaz-Castro, J.; López-Aliaga, I. New perspectives in fermented dairy products and their health relevance. J. Funct. Foods 2020, 72, 104059. [Google Scholar] [CrossRef]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic acid bacteria isolation from spontaneous sourdough and their characterization including antimicrobial and antifungal properties evaluation. Microorganisms 2019, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health benefits of lactic acid bacteria (LAB) fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef]
- Salas-Lais, A.G.; Robles-Contreras, A.; Balderas-López, J.A.; Bautista-de Lucio, V.M. Immunobiotic and paraprobiotic potential effect of Lactobacillus casei in a systemic toxoplasmosis murine model. Microorganisms 2020, 8, 113. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Wang, J.; Yan, L.; Chen, W.; Liu, X.; Zhang, H. In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains. LWT Food Sci. Technol. 2009, 42, 1640–1646. [Google Scholar] [CrossRef]
- Ya, T.; Zhang, Q.; Chu, F.; Merritt, J.; Bilige, M.; Sun, T.; Du, R.; Zhang, H. Immunological evaluation of Lactobacillus casei Zhang: A newly isolated strain from koumiss in Inner Mongolia, China. BMC Immunol. 2008, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, W.; Zhong, Z.; Wei, A.; Bao, Q.; Zhang, Y.; Sun, T.; Postnikoff, A.; Meng, H.; Zhang, H. Gene expression profile of probiotic Lactobacillus casei Zhang during the late stage of milk fermentation. Food Control 2012, 25, 321–327. [Google Scholar] [CrossRef]
- Dantas, A.B.; Jesus, V.F.; Silva, R.; Almada, C.N.; Esmerino, E.A.; Cappato, L.P.; Silva, M.C.; Raices, R.S.; Cavalcanti, R.N.; Carvalho, C.C.; et al. Manufacture of probiotic Minas Frescal cheese with Lactobacillus casei Zhang. J. Dairy Sci. 2016, 99, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, L.; Zhang, J.; Li, Y.; He, Q.; Li, H.; Guo, X.; Guo, J.; Zhang, H. Probiotic Lactobacillus casei Zhang ameliorates high-fructose-induced impaired glucose tolerance in hyperinsulinemia rats. Eur. J. Nutr. 2014, 53, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Zhang, W.; Du, R.; Meng, H.; Zhang, H. Lactobacillus casei Zhang stimulates lipid metabolism in hypercholesterolemic rats by affecting gene expression in the liver. Eur. J. Lipid Sci. Tech. 2012, 114, 244–252. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Xie, J.; Zhang, Y.; Wang, J.; Sun, X.; Zhang, H. Protective effects of probiotic Lactobacillus casei Zhang against endotoxin- and d-galactosamine-induced liver injury in rats via anti-oxidative and anti-inflammatory capacities. Int. Immunopharmacol. 2013, 15, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, C.; Zhao, J.; Xu, H.; Hou, Q. Lactobacillus casei Zhang and vitamin K2 prevent intestinal tumorigenesis in mice via adiponectin-elevated different signaling pathways. Oncotarget 2017, 8, 24719–24727. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Hou, Q.; Wang, Y.; Shen, L.; Sun, Z.; Zhang, H.; Liong, M.-T.; Kwok, L.-Y. Long-term administration of Lactobacillus casei Zhang stabilized gut microbiota of adults and reduced gut microbiota age index of older adults. J. Funct. Foods 2020, 64, 103682. [Google Scholar] [CrossRef]
- Kwok, L.Y.; Wang, L.; Zhang, J.; Guo, Z.; Zhang, H. A pilot study on the effect of Lactobacillus casei Zhang on intestinal microbiota parameters in Chinese subjects of different age. Benef. Microbes 2014, 5, 295–304. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Guo, Z.; Sun, Z.; Gesudu, Q.; Kwok, L.; Menghebilige; Zhang, H. 454 pyrosequencing reveals changes in the faecal microbiota of adults consuming Lactobacillus casei Zhang. FEMS Microbiol. Ecol. 2014, 88, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Mbye, M.; Baig, M.A.; AbuQamar, S.F.; El-Tarabily, K.A.; Obaid, R.S.; Osaili, T.M.; Al-Nabulsi, A.A.; Turner, M.S.; Shah, N.P.; Ayyash, M.M. Updates on understanding of probiotic lactic acid bacteria responses to environmental stresses and highlights on proteomic analyses. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1110–1124. [Google Scholar] [CrossRef] [Green Version]
- Grujović, M.Ž.; Mladenović, K.G.; Nikodijević, D.D.; Čomić, L.R. Autochthonous lactic acid bacteria-presentation of potential probiotics application. Biotechnol. Lett. 2019, 41, 1319–1331. [Google Scholar] [CrossRef]
- De Angelis, M.; Gobbetti, M. Environmental stress responses in Lactobacillus: A review. Proteomics 2004, 4, 106–122. [Google Scholar] [CrossRef]
- Wu, R.; Zhang, W.; Sun, T.; Wu, J.; Yue, X.; Meng, H.; Zhang, H. Proteomic analysis of responses of a new probiotic bacterium Lactobacillus casei Zhang to low acid stress. Int. J. Food Microbiol. 2011, 147, 181–187. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, J.; Wang, M.; Du, G.; Chen, J. Lactobacillus casei combats acid stress by maintaining cell membrane functionality. J. Ind. Microbiol. Biotechnol. 2012, 39, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, J.; Du, G.; Chen, J. Aspartate protects Lactobacillus casei against acid stress. Appl. Microbiol. Biotechnol. 2013, 97, 4083–4093. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, H.; Cao, C.; Li, L.; Kwok, L.Y.; Zhang, H.; Sun, Z. Adaptation of Lactobacillus casei Zhang to gentamycin involves an alkaline shock protein. Front. Microbiol. 2017, 8, 2316. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Sun, Z.; Wu, J.; Meng, H.; Zhang, H. Effect of bile salts stress on protein synthesis of Lactobacillus casei Zhang revealed by 2-dimensional gel electrophoresis. J. Dairy Sci. 2010, 93, 3858–3868. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Huang, S.; Wang, J.; Jan, G.; Jeantet, R.; Chen, X.D. Mg(2+) improves the thermotolerance of probiotic Lactobacillus rhamnosus GG, Lactobacillus casei Zhang and Lactobacillus plantarum P-8. Lett. Appl. Microbiol. 2017, 64, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Oberg, C.J.; Moyes, L.V.; Domek, M.J.; Brothersen, C.; McMahon, D.J. Survival of probiotic adjunct cultures in cheese and challenges in their enumeration using selective media. J. Dairy Sci. 2011, 94, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Renschler, M.A.; Wyatt, A.; Anene, N.; Robinson-Hill, R.; Pickerill, E.S.; Fox, N.E.; Griffith, J.A.; McKillip, J.L. Using nitrous acid-modified de Man, Rogosa, and Sharpe medium to selectively isolate and culture lactic acid bacteria from dairy foods. J. Dairy Sci. 2020, 103, 1215–1222. [Google Scholar] [CrossRef]
- Li, S.; Jina, Z.; Hu, D.; Yang, W.; Yan, Y.; Nie, X.; Lin, J.; Zhang, Q.; Gai, D.; Jia, Y.; et al. Effect of solid-state fermentation with Lactobacillus casei on the nutritional value, isoflavones, phenolic acids and antioxidant activity of whole soybean flour. LWT Food Sci. Technol. 2020, 125, 109264. [Google Scholar] [CrossRef]
- Zhang, S.; Shi, Y.; Zhang, S.; Shang, W.; Gao, X.; Wang, H. Whole soybean as probiotic lactic acid bacteria carrier food in solid-state fermentation. Food Control 2014, 41, 1–6. [Google Scholar] [CrossRef]
- Cock, P.J.; Fields, C.J.; Goto, N.; Heuer, M.L.; Rice, P.M. The Sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, J. Citrate metabolism in lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 165–178. [Google Scholar] [CrossRef]
- De Angelis, M.; Gobbetti, M. Stress Responses of Lactobacilli. In Stress Responses of Lactic Acid Bacteria. Food Microbiology and Food Safety; Tsakalidou, E., Papadimitriou, K., Eds.; Springer: Boston, MA, USA, 2011; pp. 219–249. [Google Scholar]
- Zhai, Z.; Yang, Y.; Wang, H.; Wang, G.; Ren, F.; Li, Z.; Hao, Y. Global transcriptomic analysis of Lactobacillus plantarum CAUH2 in response to hydrogen peroxide stress. Food Microbiol. 2020, 87, 103389. [Google Scholar] [CrossRef]
- Serrano, L.M.; Molenaar, D.; Wels, M.; Teusink, B.; Bron, P.A.; de Vos, W.M.; Smid, E.J. Thioredoxin reductase is a key factor in the oxidative stress response of Lactobacillus plantarum WCFS1. Microb. Cell Fact. 2007, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Pawłowicz, I.; Masajada, K. Aquaporins as a link between water relations and photosynthetic pathway in abiotic stress tolerance in plants. Gene 2019, 687, 166–172. [Google Scholar] [CrossRef]
- Bienert, G.P.; Desguin, B.; Chaumont, F.; Hols, P. Channel-mediated lactic acid transport: A novel function for aquaglyceroporins in bacteria. Biochem. J. 2013, 454, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Vemanna, R.S.; Babitha, K.C.; Solanki, J.K.; Amarnatha Reddy, V.; Sarangi, S.K.; Udayakumar, M. Aldo-keto reductase-1 (AKR1) protect cellular enzymes from salt stress by detoxifying reactive cytotoxic compounds. Plant Physiol. Biochem. 2017, 113, 177–186. [Google Scholar] [CrossRef]
- Ma’ayeh, S.Y.; Knörr, L.; Svärd, S.G. Transcriptional profiling of Giardia intestinalis in response to oxidative stress. Int. J. Parasitol. 2015, 45, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.L.; Lindsay, E.L.; Faustoferri, R.C.; To, T.T.; Hendrickson, E.L.; He, X.; Shi, W.; McLean, J.S.; QuiveyJr, R.G. Characterization of the trehalose utilization operon in Streptococcus mutans reveals that the TreR transcriptional regulator is involved in stress response pathways and toxin production. J. Bacteriol. 2018, 200, e00057-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Teng, K.; Su, R.; Li, L.; Zhang, T.; Fan, K.; Zhang, J.; Zhong, J. AcrR and Rex control mannitol and sorbitol utilization through their cross-regulation of aldehyde-alcohol dehydrogenase (AdhE) in Lactobacillus plantarum. Appl. Environ. Microbiol. 2019, 85, e02035-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez, L.; Solopova, A.; Fernández-Pérez, R.; González, M.; Tenorio, C.; Kuipers, O.P.; Ruiz-Larrea, F. Transcriptome analysis shows activation of the arginine deiminase pathway in Lactococcus lactis as a response to ethanol stress. Int. J. Food Microbiol. 2017, 257, 41–48. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.; An, J.; Chen, L.; Zhu, Q.; Li, Y.; Mei, Y.; Chen, Z.; Liang, Y. Differential Analysis of Stress Tolerance and Transcriptome of Probiotic Lacticaseibacillus casei Zhang Produced from Solid-State (SSF-SW) and Liquid-State (LSF-MRS) Fermentations. Microorganisms 2020, 8, 1656. https://doi.org/10.3390/microorganisms8111656
Wu P, An J, Chen L, Zhu Q, Li Y, Mei Y, Chen Z, Liang Y. Differential Analysis of Stress Tolerance and Transcriptome of Probiotic Lacticaseibacillus casei Zhang Produced from Solid-State (SSF-SW) and Liquid-State (LSF-MRS) Fermentations. Microorganisms. 2020; 8(11):1656. https://doi.org/10.3390/microorganisms8111656
Chicago/Turabian StyleWu, Pengyu, Jing An, Liang Chen, Qiuyan Zhu, Yingjun Li, Yuxia Mei, Zhenmin Chen, and Yunxiang Liang. 2020. "Differential Analysis of Stress Tolerance and Transcriptome of Probiotic Lacticaseibacillus casei Zhang Produced from Solid-State (SSF-SW) and Liquid-State (LSF-MRS) Fermentations" Microorganisms 8, no. 11: 1656. https://doi.org/10.3390/microorganisms8111656