Transcriptomic and Phenotypic Analyses of the Sigma B-Dependent Characteristics and the Synergism between Sigma B and Sigma L in Listeria monocytogenes EGD-e


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Motility Assays and Electron Microscopy
2.3. DNA Manipulation and Construction of the ΔsigB and ΔsigBL Strains
2.4. RNA Isolation and Gene-Expression Analysis
3. Results
3.1. Phenotypic Characterization of ΔsigB and ΔsigBL
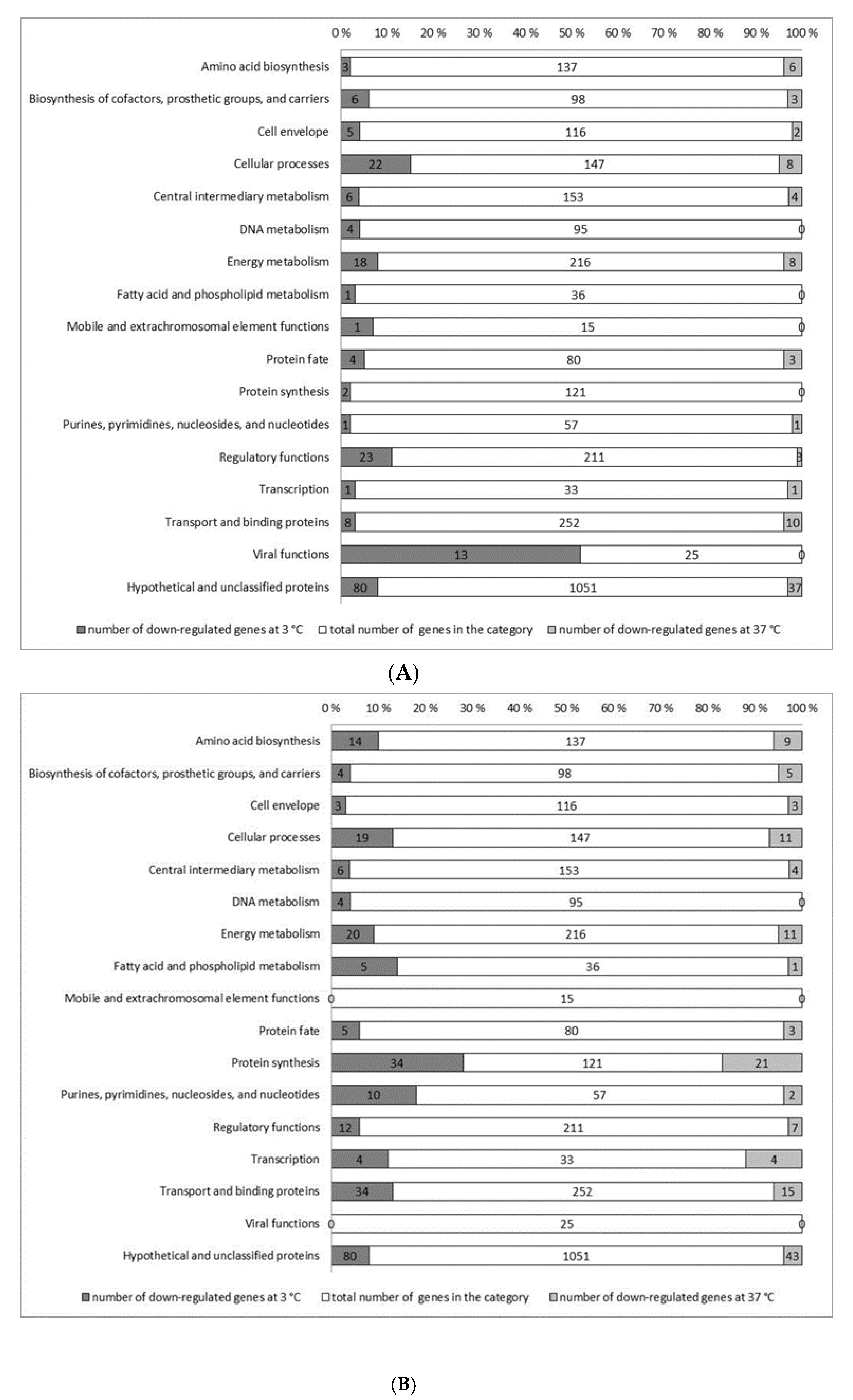
3.2. Identification of Genes de-Regulated in ΔsigB and ΔsigBL Mutants
3.3. Genes Down-Regulated in ∆sigB During Exponential Growth in BHI at 3 °C
3.4. Genes Down-Regulated in ∆sigB During Exponential Growth in BHI at 37 °C
3.5. Genes Down−Regulated in ∆sigBL during Exponential Growth in BHI at 3 °C
3.6. Genes Down−Regulated in ∆sigBL During Exponential Growth in BHI at 37 °C
4. Discussion
4.1. Identification of σB Positively Regulated Genes during Exponential Growth in BHI at 3 and 37 °C
4.2. Stress Response Phenotypic Traits Associated with a ∆sigB Mutation in L. Monocytogenes EGD−e
4.3. Impact of the ∆sigBL Mutation on the L. Monocytogenes EGD−e Transcriptome
4.4. Stress Response Phenotypic Traits Associated with σBand σL Inactivation in L. Monocytogenes EGD−e
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Junttila, J.R.; Niemelä, S.I.; Hirn, J. Minimum growth temperatures of Listeria monocytogenes and non−haemolytic Listeria. J. Appl. Bacteriol. 1988, 65, 321–327. [Google Scholar] [CrossRef]
- Cole, M.B.; Jones, M.V.; Holyoak, C. The effect of pH, salt concentration and temperature on the survival and growth of Listeria monocytogenes. J. Appl. Bacteriol. 1990, 69, 63–72. [Google Scholar] [CrossRef]
- Vasseur, C.; Baverel, L.; Hébraud, M.; Labadie, J. Effect of osmotic, alkaline, acid or thermal stresses on the growth and inhibition of Listeria monocytogenes. J. Appl. Microbiol. 1999, 86, 469–476. [Google Scholar] [CrossRef]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food−borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [CrossRef] [Green Version]
- Autio, T.; Hielm, S.; Miettinen, M.; Sjöberg, A.M.; Aarnisalo, K.; Björkroth, J.; Mattila−Sandholm, T.; Korkeala, H. Sources of Listeria monocytogenes contamination in a cold−smoked rainbow trout processing plant detected by pulsed−field gel electrophoresis typing. Appl. Environ. Microbiol. 1999, 65, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Tompkin, R.B. Control of Listeria monocytogenes in the food−processing environment. J. Food Prot. 2002, 65, 709–725. [Google Scholar] [CrossRef]
- Lundén, J.M.; Autio, T.J.; Sjöberg, A.M.; Korkeala, H.J. Persistent and nonpersistent Listeria monocytogenes contamination in meat and poultry processing plants. J. Food Prot. 2003, 66, 2062–2069. [Google Scholar] [CrossRef] [Green Version]
- Keto−Timonen, R.; Tolvanen, R.; Lundén, J.; Korkeala, H. An 8−year surveillance of the diversity and persistence of Listeria monocytogenes in a chilled food processing plant analyzed by amplified fragment length polymorphism. J. Food Prot. 2007, 70, 1866–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lianou, A.; Sofos, J.N. A review of the incidence and transmission of Listeria monocytogenes in ready−to−eat products in retail and food service environments. J. Food Prot. 2007, 70, 2172–2198. [Google Scholar] [CrossRef]
- Kurpas, M.; Wieczorek, K.; Osek, J. Ready−to−eat meat products as a source of Listeria monocytogenes. J. Vet. Res. 2018, 62, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Koutsoumanis, K.; Alvarez−Ordóñez, A.; Bolton, D.; Bover−Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R. The public health risk posed by Listeria monocytogenes in frozen fruit and vegetables including herbs, blanched during processing. EFSA J. 2020, 18, e06092. [Google Scholar] [PubMed] [Green Version]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar]
- Francke, C.; Groot Kormelink, T.; Hagemeijer, Y.; Overmars, L.; Sluijter, V.; Moezelaar, R.; Siezen, R.J. Comparative analyses imply that the enigmatic Sigma factor 54 is a central controller of the bacterial exterior. BMC Genom. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Becker, L.A.; Cetin, M.S.; Hutkins, R.W.; Benson, A.K. Identification of the gene encoding the alternative sigma factor sigmaB from Listeria monocytogenes and its role in osmotolerance. J. Bacteriol. 1998, 180, 4547–4554. [Google Scholar] [CrossRef] [Green Version]
- Wiedmann, M.; Arvik, T.J.; Hurley, R.J.; Boor, K.J. General stress transcription factor sigmaB and its role in acid tolerance and virulence of Listeria monocytogenes. J. Bacteriol. 1998, 180, 3650–3656. [Google Scholar] [CrossRef] [Green Version]
- Kazmierczak, M.J.; Mithoe, S.C.; Boor, K.J.; Wiedmann, M. Listeria monocytogenes sigma B regulates stress response and virulence functions. J. Bacteriol. 2003, 185, 5722–5734. [Google Scholar] [CrossRef] [Green Version]
- Wemekamp−Kamphuis, H.H.; Wouters, J.A.; de Leeuw, P.P.; Hain, T.; Chakraborty, T.; Abee, T. Identification of sigma factor sigma B−controlled genes and their impact on acid stress, high hydrostatic pressure, and freeze survival in Listeria monocytogenes EGD−e. Appl. Environ. Microbiol. 2004, 70, 3457–3466. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.C.; Boor, K.J.; Wiedmann, M. SigmaB−dependent and sigmaB−independent mechanisms contribute to transcription of Listeria monocytogenes cold stress genes during cold shock and cold growth. Appl. Environ. Microbiol. 2007, 73, 6019–6029. [Google Scholar] [CrossRef] [Green Version]
- Raengpradub, S.; Wiedmann, M.; Boor, K.J. Comparative analysis of the sigma B−dependent stress responses in Listeria monocytogenes and Listeria innocua strains exposed to selected stress conditions. Appl. Environ. Microbiol. 2008, 74, 158–171. [Google Scholar] [CrossRef] [Green Version]
- Somolinos, M.; Espina, L.; Pagán, R.; Garcia, D. sigB absence decreased Listeria monocytogenes EGD−e heat resistance but not its Pulsed Electric Fields resistance. Int. J. Food Microbiol. 2010, 141, 32–38. [Google Scholar] [CrossRef]
- Guerreiro, D.N.; Arcari, T.; O’Byrne, C.P. The σB−Mediated General Stress Response of Listeria monocytogenes: Life and Death Decision Making in a Pathogen. Front. Microbiol. 2020, 11, 1505. [Google Scholar] [CrossRef] [PubMed]
- Nadon, C.A.; Bowen, B.M.; Wiedmann, M.; Boor, K.J. Sigma B contributes to PrfA−mediated virulence in Listeria monocytogenes. Infect. Immun. 2002, 70, 3948–3952. [Google Scholar] [CrossRef] [Green Version]
- McGann, P.; Wiedmann, M.; Boor, K.J. The alternative sigma factor sigma B and the virulence gene regulator PrfA both regulate transcription of Listeria monocytogenes internalins. Appl. Environ. Microbiol. 2007, 73, 2919–2930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollinger, J.; Wiedmann, M.; Boor, K.J. SigmaB− and PrfA−dependent transcription of genes previously classified as putative constituents of the Listeria monocytogenes PrfA regulon. Foodborne Pathog. Dis. 2008, 5, 281–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, K.R.; Sue, D.; Wiedmann, M.; Boor, K.; O’Byrne, C.P. Role of SigmaB in regulating the compatible solute uptake systems of Listeria monocytogenes: Osmotic induction of opuC is SigmaB dependent. Appl. Environ. Microbiol. 2003, 69, 2015–2022. [Google Scholar] [CrossRef] [Green Version]
- Abram, F.; Starr, E.; Karatzas, K.A.; Matlawska−Wasowska, K.; Boyd, A.; Wiedmann, M.; Boor, K.J.; Connally, D.; O’Byrne, C.P. Identification of components of the Sigma B regulon in Listeria monocytogenes that contribute to acid and salt tolerance. Appl. Environ. Microbiol 2008, 74, 6848–6858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, H.F.; Orsi, R.H.; Wiedmann, M.; Boor, K.J. Listeria monocytogenes {Sigma}B has a small core regulon and a conserved role in virulence but makes differential contributions to stress tolerance across a diverse collection of strains. Appl. Environ. Microbiol. 2010, 76, 4216–4232. [Google Scholar] [CrossRef] [Green Version]
- Chaturongakul, S.; Raengpradub, S.; Palmer, M.E.; Bergholz, T.M.; Orsi, R.H.; Hu, Y.; Ollinger, J.; Wiedmann, M.; Boor, K.J. Transcriptomic and phenotypic analyses identify coregulated, overlapping regulons among PrfA, CtsR, HrcA, and the alternative sigma factors SigmaB, SigmaC, SigmaH, and SigmaL in Listeria monocytogenes. Appl. Environ. Microbiol. 2011, 77, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Mujahid, S.; Orsi, R.H.; Boor, K.J.; Wiedmann, M. Protein level identification of the Listeria monocytogenes sigma H, sigma L, and sigma C regulons. BMC Microbiol. 2013, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Mujahid, S.; Orsi, R.H.; Vangay, P.; Boor, K.J.; Wiedmann, M. Refinement of the Listeria monocytogenes σB regulon through quantitative proteomic analysis. Microbiology 2013, 159, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Orsi, R.H.; Boor, K.J.; Wiedmann, M.; Guariglia−Oropeza, V. Home alone: Elimination of all but one alternative sigma factor in Listeria monocytogenes allows prediction of new roles for σB. Front. Microbiol. 2017, 8, 1910. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Orsi, R.H.; Gaballa, A.; Wiedmann, M.; Boor, K.J.; Guariglia−Oropeza, V. Systematic review of the Listeria monocytogenes σB regulon supports a role in stress response, virulence and metabolism. Future Microbiol. 2019, 14, 801–828. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Okada, N.; Makino, S.; Asakura, H.; Yamamoto, S.; Igimi, S. The sigma factor RpoN (sigma54) is involved in osmotolerance in Listeria monocytogenes. FEMS Microbiol. Lett. 2006, 263, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.C.; Hu, Y.; Chaturongakul, S.; Files, K.D.; Bowen, B.M.; Boor, K.J.; Wiedmann, M. Contributions of two−component regulatory systems, alternative sigma factors, and negative regulators to Listeria monocytogenes cold adaptation and cold growth. J. Food Prot. 2008, 71, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Raimann, E.; Schmid, B.; Stephan, R.; Tasara, T. The alternative sigma factor Sigma(L) of L. monocytogenes promotes growth under diverse environmental stresses. Foodborne Pathog. Dis. 2009, 6, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Mattila, M.; Somervuo, P.; Rattei, T.; Korkeala, H.; Stephan, R.; Tasara, T. Phenotypic and transcriptomic analyses of Sigma L−dependent characteristics in Listeria monocytogenes EGD−e. Food Microbiol. 2012, 32, 152–164. [Google Scholar] [CrossRef]
- Liu, S.; Graham, J.E.; Bigelow, L.; Morse, P.D.; Wilkinson, B.J. Identification of Listeria monocytogenes genes expressed in response to growth at low temperature. Appl. Environ. Microbiol. 2002, 68, 1697–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arous, S.; Buchrieser, C.; Folio, P.; Glaser, P.; Namane, A.; Hébraud, M.; Héchard, Y. Global analysis of gene expression in an rpoN mutant of Listeria monocytogenes. Microbiology 2004, 150, 1581–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premaratne, R.J.; Lin, W.J.; Johnson, E.A. Development of an improved chemically defined minimal medium for Listeria monocytogenes. Appl. Environ. Microbiol. 1991, 57, 3046–3048. [Google Scholar] [CrossRef] [Green Version]
- Michel, E.; Stephan, R.; Tasara, T. The lmo0501 gene coding for a putative transcription activator protein in Listeria monocytogenes promotes growth under cold, osmotic and acid stress conditions. Food Microbiol. 2011, 28, 1261–1265. [Google Scholar] [CrossRef] [Green Version]
- Loepfe, C.; Raimann, E.; Stephan, R.; Tasara, T. Reduced host cell invasiveness and oxidative stress tolerance in double and triple csp gene family deletion mutants of Listeria monocytogenes. Foodborne Pathog. Dis. 2010, 7, 775–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high−throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Kathariou, S.; Mizumoto, C.; Kanenaka, R.; Allen, R.D.; Fok, A.K. Repression of motility and flagellin production at 37 C is stronger in Listeria monocytogenes than in the nonpathogenic species Listeria innocua. Can. J. Microbiol. 1995, 41, 572–577. [Google Scholar] [CrossRef]
- Schmid, B.; Klumpp, J.; Raimann, E.; Loessner, M.J.; Stephan, R.; Tasara, T. Role of cold shock proteins in growth of Listeria monocytogenes under cold and osmotic stress conditions. Appl. Environ. Microbiol. 2009, 75, 1621–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, R.M.; Cai, Z.L.; Ho, S.N.; Pease, L.R. Gene splicing by overlap extension: Tailor−made genes using the polymerase chain reaction. Biotechniques 1990, 8, 528–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.; Youngman, P. Use of a new integrational vector to investigate compartment−specific expression of the Bacillus subtilis spoIIM gene. Biochimie 1992, 74, 705–711. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Toledo−Arana, A.; Dussurget, O.; Nikitas, G.; Sesto, N.; Guet−Revillet, H.; Balestrino, D.; Loh, E.; Gripenland, J.; Tiensuu, T.; Vaitkevicius, K.; et al. The Listeria transcriptional landscape from saprophytism to virulence. Nature 2009, 459, 950–956. [Google Scholar] [CrossRef]
- van Schaik, W.; Abee, T. The role of sigmaB in the stress response of Gram–positive bacteria–targets for food preservation and safety. Curr. Opin. Biotechnol. 2005, 16, 218–224. [Google Scholar] [CrossRef]
- Marles−Wright, J.; Lewis, R.J. Stress responses of bacteria. Curr. Opin. Struct. Biol. 2007, 17, 755–760. [Google Scholar] [CrossRef]
- Chaturongakul, S.; Raengpradub, S.; Wiedmann, M.; Boor, K.J. Modulation of stress and virulence in Listeria monocytogenes. Trends Microbiol. 2008, 16, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorski, L.; Duhé, J.M.; Flaherty, D. The Sigma B operon is a determinant of fitness for a Listeria monocytogenes serotype 4b strain in soil. Foodborne Pathog. Dis. 2011, 8, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Palonen, E.; Lindström, M.; Somervuo, P.; Korkeala, H. Alternative sigma factor σE has an important role in stress tolerance of Yersinia pseudotuberculosis IP32953. Appl. Environ. Microbiol. 2013, 79, 5970–5977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hain, T.; Hossain, H.; Chatterjee, S.S.; Machata, S.; Volk, U.; Wagner, S.; Brors, B.; Haas, S.; Kuenne, C.T.; Billion, A.; et al. Temporal transcriptomic analysis of the Listeria monocytogenes EGD−e sigmaB regulon. BMC Microbiol. 2008, 28, 20. [Google Scholar]
- Anderson, L.A.; McNairn, E.; Lubke, T.; Pau, R.N.; Boxer, D.H. ModE−dependent molybdate regulation of the molybdenum cofactor operon moa in Escherichia coli. J. Bacteriol. 2000, 182, 7035–7043. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.; O’Byrne, C.P.; Boor, K.J. Role of sigma(B) in heat, ethanol, acid, and oxidative stress resistance and during carbon starvation in Listeria monocytogenes. Appl. Environ. Microbiol. 2001, 67, 4454–4457. [Google Scholar] [CrossRef] [Green Version]
- Moorhead, S.M.; Dykes, G.A. The role of the sigB gene in the general stress response of Listeria monocytogenes varies between a strain of serotype 1/2a and a strain of serotype 4c. Curr. Microbiol. 2003, 46, 461–466. [Google Scholar] [CrossRef]
- Hu, Y.; Raengpradub, S.; Schwab, U.; Loss, C.; Orsi, R.H.; Wiedmann, M.; Boor, K.J. Phenotypic and transcriptomic analyses demonstrate interactions between the transcriptional regulators CtsR and Sigma B in Listeria monocytogenes. Appl. Environ. Microbiol. 2007, 73, 7967–7980. [Google Scholar] [CrossRef] [Green Version]
- Garmyn, D.; Augagneur, Y.; Gal, L.; Vivant, A.L.; Piveteau, P. Listeria monocytogenes differential transcriptome analysis reveals temperature−dependent Agr regulation and suggests overlaps with other regulons. PLoS ONE 2012, 7, e43154. [Google Scholar] [CrossRef]
- Tang, S.; Orsi, R.H.; den Bakker, H.C.; Wiedmann, M.; Boor, K.J.; Bergholz, T.M. Transcriptomic analysis of the adaptation of Listeria monocytogenes to growth on vacuum−packed cold smoked salmon. Appl. Environ. Microbiol. 2015, 81, 6812–6824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Z.; Smid, E.J.; Boeren, S.; Notebaart, R.A.; Abee, T. Bacterial Microcompartment−Dependent 1,2−Propanediol Utilization Stimulates Anaerobic Growth of Listeria monocytogenes EGDe. Front. Microbiol. 2019, 10, 2660. [Google Scholar] [CrossRef] [PubMed]
- Tojo, S.; Satomura, T.; Morisaki, K.; Yoshida, K.; Hirooka, K.; Fujita, Y. Negative transcriptional regulation of the ilv−leu operon for biosynthesis of branched−chain amino acids through the Bacillus subtilis global regulator TnrA. J. Bacteriol. 2004, 186, 7971–7979. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M.E.; Wiedmann, M.; Boor, K.J. Sigma(B) and sigma(L) contribute to Listeria monocytogenes 10403S response to the antimicrobial peptides SdpC and nisin. Foodborne Pathog Dis. 2009, 6, 1057–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]



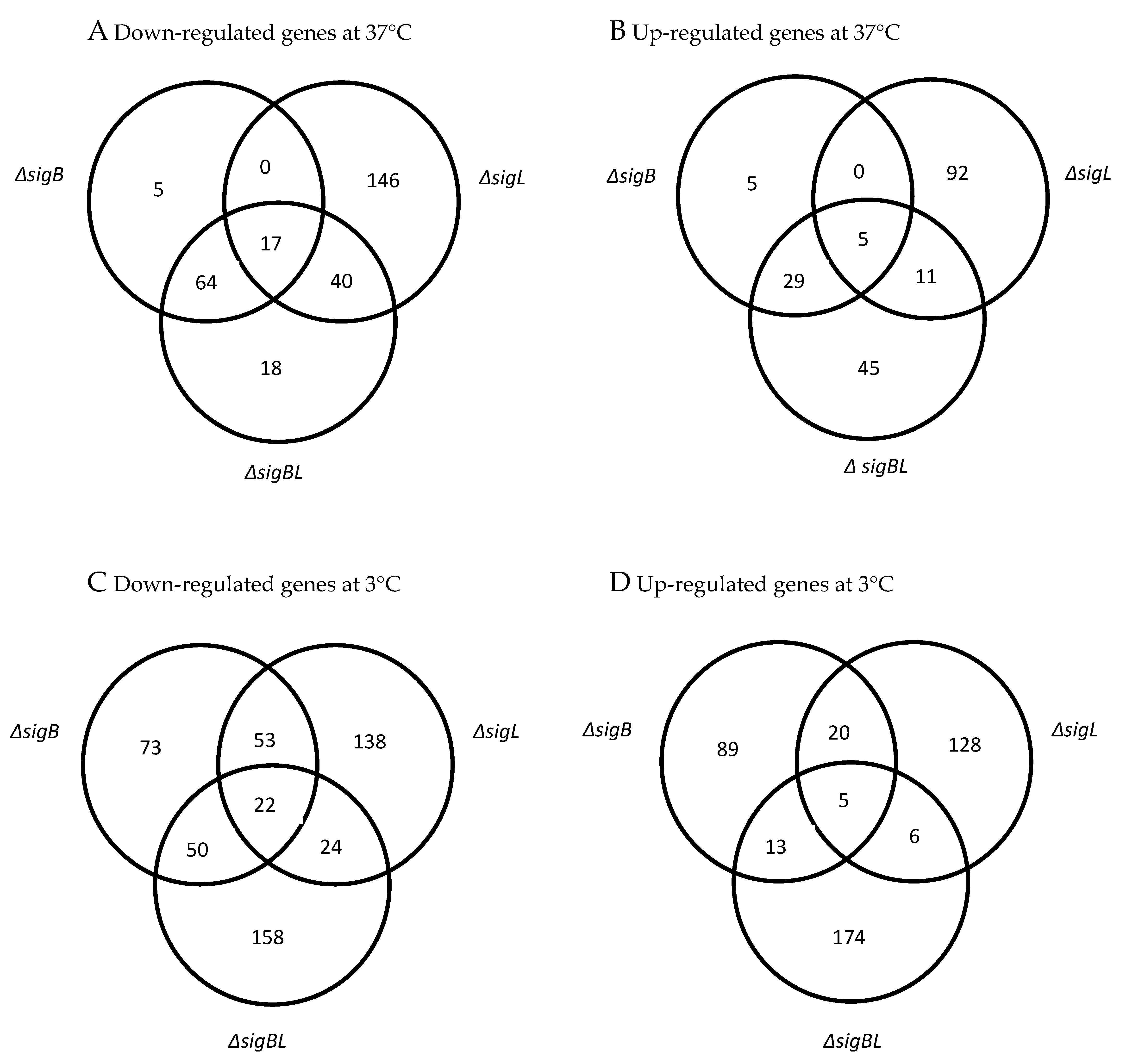{kind=link}
{kind=link}
| Growth Condition | Growth Parameter | WT | ΔsigB | ΔsigBL | Statistical Significance |
|---|---|---|---|---|---|
| 4 °C BHI (pH 7.4) | LPD (wk) GR (Log10(CFU/mL)/wk) | 0.00 0.89 ± 0.03 | 0.00 0.88 ± 0.04 | 0.00 0.85 ± 0.04 | NS a NS |
| BHI plus 2.5% lactic acid (pH 6.0) | LPD (wk) GR (Log10(CFU/mL)/wk) | 3.84 ± 0.10 1.01 ± 0.01 | 4.51 ± 0.15 1.10 ± 0.02 | 5.48 ± 0.67 0.32 ± 0.01 | S b,c S |
| BHI plus 0.35% acetic acid (pH 5.5) | LPD (wk) GR (Log10(CFU/mL)/wk) | 2.10 ± 1.32 0.13 ± 0.01 | 7.33 ± 0.47 0.23 ± 0.03 | NG NG | S S |
| BHI plus 0.35% citric acid (pH 5.5) | LPD (wk) GR (Log10(CFU/mL)/wk) | 3.98 ± 0.96 1.11 ± 0.15 | 2.82 ± 0.10 1.21 ± 0.11 | NG NG | S S |
| DM | LPD (wk) GR (Log10(CFU/mL)/wk) | 2.44 ± 0.18 1.27 ± 0.07 | 2.87 ± 0.05 1.63 ± 0.11 | 4.60 ± 0.27 0.49 ± 0.17 | S S |
| 37 °C DM | LPD (wk) GR (Log10(CFU/mL)/wk) | 0.00 0.14 ± 0.01 | 0.00 0.12 ± 0.01 | 0.00 0.11 ± 0.03 | NS NS |
| DM plus 5% Ethanol | LPD (wk) GR (Log10(CFU/mL)/wk) | 5.87 ± 2.51 0.12 ± 0.02 | 17.79 ± 1.78 0.04 ± 0.05 | 14.27 ± 1.58 0.03 ± 0.02 | S S |
| Gene | 37 °C | 3 °C | ||||||
|---|---|---|---|---|---|---|---|---|
| ΔsigB | ΔsigBL | ΔsigB | ΔsigBL | |||||
| Microarray | RT-qPCR | Microarray | RT-qPCR | Microarray | RT-qPCR | Microarray | RT-qPCR | |
| lmo0096 | ▼ | ▼ | ▼ | ▼ | ▲ | ▲ | NS | NS |
| lmo0137 | ▼ | ▼ | ▼ | ▼ | NS | NS | ▼ | ▼ |
| lmo0685 | NS a | NS | NS | NS | NS | NS | ▼ | ▼ |
| lmo2625 | ▼ | ▼ | ▼ | ▼ | NS | NS | ▼ | ▼ |
| Gene | Functional Category and Protein b | Differential Expression (∆sigB vs. WT) Fold Change at 3 °C | Differential Expression (∆sigB vs. WT) Fold Change at 37 °C |
|---|---|---|---|
| Amino acid biosynthesis | |||
| lmo0781 | PTS system component | −1.5 | −2.3 |
| Cell envelope | |||
| lmo1079 | membrane protein, putative | −1.4 | −2.1 |
| Cellular processes | |||
| lmo0263 | internalin H | −3 | −4.1 |
| lmo0515 | universal stress protein family | −2.9 | −2.5 |
| lmo0669 | general stress protein 39 | −1.8 | −3.5 |
| lmo1694 | cell division inhibitor | −1.5 | −2.4 |
| lmo2230 | arsenate reductase, putative | −4 | −3.9 |
| lmo2673 | universal stress protein family | −2.4 | −2.8 |
| Central intermediary metabolism | |||
| lmo0134 | acetyltransferase, GNAT family | −2.4 | −1.4 |
| lmo2434 | glutamate decarboxylase | −3.2 | −3.1 |
| Energy metabolism | |||
| lmo0722 | pyruvate oxidase | −2.3 | −2.4 |
| lmo0913 | succinate−semialdehyde dehydrogenase | −2.5 | −1.6 |
| lmo2674 | ribose 5−phosphate isomerase B | −1.8 | −1.6 |
| Protein fate | |||
| lmo1407 | pyruvate formate−lyase activating enzyme | −1.4 | −1.5 |
| lmo2157 | secretory protein (sepA) | −1.4 | −2.2 |
| Regulatory functions | |||
| lmo2085 | Gram positive anchor domain protein | −3.3 | −3 |
| Transcription | |||
| lmo0895 | RNA polymerase sigma factor B | −5.8 | −5.3 |
| Transport and binding proteins | |||
| lmo0169 | transporter, putative | −2.1 | −2.4 |
| lmo0782 | PTS system component | −3 | −3.4 |
| lmo0783 | PTS system component | −3.3 | −2.8 |
| lmo0784 | PTS system component | −3.7 | −3.2 |
| Hypothetical and unclassified proteins | |||
| lmo0265 | peptidase, M20/M25/M40 family | −2.2 | −2.4 |
| lmo0602 | protease synthase and sporulation negative regulatory protein pai 1, putative | −1.3 | −2.8 |
| lmo0937 | hypothetical protein | −3.2 | −2.8 |
| lmo1241 | conserved hypothetical protein | −2.5 | −3.9 |
| lmo2067 | choloylglycine hydrolase | −3.1 | −3.5 |
| lmo2158 | conserved domain protein | −6.2 | −3.9 |
| lmo2213 | conserved hypothetical protein | −1.6 | −2.6 |
| lmo2748 | conserved hypothetical protein | −1.9 | −3 |
| Gene | Functional Category and Protein b | Differential Expression (∆sigBL vs. WT) Fold Change at 3 °C | Differential Expression (∆sigBL vs. WT) Fold Change at 37 °C |
|---|---|---|---|
| Biosynthesis of cofactors, prosthetic groups, and carriers | |||
| lmo2571 | pyrazinamidase/nicotinamidase, putative | −1.4 | −3.4 |
| Cellular processes | |||
| lmo0263 | internalin H | −2.7 | −4.1 |
| lmo0433 | internalin A | −3.8 | −1.3 |
| lmo0515 | universal stress protein family | −3.5 | −2.2 |
| lmo0669 | general stress protein 39 | −1.7 | −3.6 |
| lmo1694 | cell division inhibitor | −1.9 | −3 |
| lmo2230 | arsenate reductase, putative | −3.1 | −4.2 |
| lmo2673 | universal stress protein family | −1.4 | −3.3 |
| Central intermediary metabolism | |||
| lmo0134 | acetyltransferase, GNAT family | −2.5 | −3 |
| lmo2434 | glutamate decarboxylase | −3.1 | −3.4 |
| Energy metabolism | |||
| lmo0210 | L−lactate dehydrogenase | −3.2 | −1.3 |
| lmo0913 | succinate−semialdehyde dehydrogenase | −2.1 | −1.6 |
| lmo1634 | aldehyde−alcohol dehydrogenase | −6.1 | −1.7 |
| lmo2674 | ribose 5−phosphate isomerase B | −2 | −1.6 |
| Protein fate | |||
| lmo1407 | pyruvate formate−lyase activating enzyme | −3.3 | −1.5 |
| Protein synthesis | |||
| lmo0250 | ribosomal protein L10 | −2.2 | −1.3 |
| lmo0251 | ribosomal protein L7/L12 | −2.6 | −1.4 |
| lmo1783 | ribosomal protein L20 | −1.5 | −1.3 |
| lmo2605 | ribosomal protein L17 | −1.4 | −1.6 |
| lmo2613 | ribosomal protein L15 | −1.3 | −1.4 |
| lmo2614 | ribosomal protein L30 | −1.5 | −2 |
| lmo2615 | ribosomal protein S5 | −1.8 | −2 |
| lmo2616 | ribosomal protein L18 | −1.6 | −1.9 |
| lmo2617 | ribosomal protein L6 | −1.7 | −1.7 |
| lmo2618 | ribosomal protein S8 | −1.8 | −1.7 |
| lmo2619 | ribosomal protein S14p/S29e | −1.6 | −1.6 |
| lmo2620 | ribosomal protein L5 | −1.9 | −1.7 |
| lmo2621 | ribosomal protein L24 | −1.7 | −1.5 |
| lmo2622 | ribosomal protein L14 | −1.5 | −1.6 |
| lmo2624 | ribosomal protein L29 | −1.9 | −1.5 |
| lmo2625 | ribosomal protein L16 | −1.8 | −1.5 |
| lmo2626 | ribosomal protein S3 | −1.6 | −1.5 |
| Purines, pyrimidines, nucleosides, and nucleotides | |||
| lmo2611 | adenylate kinase | −1.5 | −1.9 |
| Regulatory functions | |||
| lmo1956 | transcriptional regulator, Fur family | −1.5 | −1.4 |
| lmo2085 | Gram positive anchor domain protein | −3.2 | −3.6 |
| Transcription | |||
| lmo0895 | RNA polymerase sigma factor B | −2.3 | −5.3 |
| lmo2461 | Sigma−54 factors family | −2.7 | −3.7 |
| Transport and binding proteins | |||
| lmo0135 | oligopeptide−binding protein appa precursor, putative | −2.2 | −2.8 |
| lmo0136 | peptide ABC transporter, permease protein | −1.8 | −2.4 |
| lmo0137 | oligopeptide transport permease protein appc | −1.6 | −2.6 |
| lmo0169 | transporter, putative | −1.3 | −2.7 |
| lmo0781 | PTS system component | −2 | −2.3 |
| lmo0782 | PTS system component | −2.2 | −3.9 |
| lmo0783 | PTS system component | −2.1 | −2.9 |
| lmo0784 | PTS system component | −2.1 | −3.8 |
| Hypothetical and unclassified proteins | |||
| lmo0265 | peptidase, M20/M25/M40 family | ||
| lmo0355 | succinate dehydrogenase/fumarate reductase, flavoprotein | −2.2 | −2.4 |
| lmo0602 | subunit | −1.9 | −1.9 |
| lmo2067 | protease synthase and sporulation negative regulatory protein pai 1, putative | −2.1 | −3 |
| lmo2158 | choloylglycine hydrolase | ||
| lmo0170 | conserved domain protein | −3 | −3.6 |
| lmo0596 | conserved hypothetical protein | −4.7 | −3.8 |
| lmo0911 | conserved hypothetical protein | −1.7 | −1.5 |
| lmo0995 | conserved hypothetical protein | −1.6 | −3.9 |
| lmo1241 | conserved hypothetical protein | −2 | −1.6 |
| lmo1776 | conserved hypothetical protein | −2.3 | −1.9 |
| lmo2213 | conserved hypothetical protein | −1.4 | −4.2 |
| lmo2572 | conserved hypothetical protein | −2.8 | −1.4 |
| lmo2748 | conserved hypothetical protein | −1.5 | −2.8 |
| lmo0654 | conserved hypothetical protein | −1.4 | −3.5 |
| lmo0937 | hypothetical protein | −2.5 | −3.1 |
| lmo0994 | hypothetical protein | −1.3 | −1.6 |
| lmo2454 | hypothetical protein | −3.3 | −2.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattila, M.; Somervuo, P.; Korkeala, H.; Stephan, R.; Tasara, T. Transcriptomic and Phenotypic Analyses of the Sigma B-Dependent Characteristics and the Synergism between Sigma B and Sigma L in Listeria monocytogenes EGD-e. Microorganisms 2020, 8, 1644. https://doi.org/10.3390/microorganisms8111644
Mattila M, Somervuo P, Korkeala H, Stephan R, Tasara T. Transcriptomic and Phenotypic Analyses of the Sigma B-Dependent Characteristics and the Synergism between Sigma B and Sigma L in Listeria monocytogenes EGD-e. Microorganisms. 2020; 8(11):1644. https://doi.org/10.3390/microorganisms8111644
Chicago/Turabian StyleMattila, Mirjami, Panu Somervuo, Hannu Korkeala, Roger Stephan, and Taurai Tasara. 2020. "Transcriptomic and Phenotypic Analyses of the Sigma B-Dependent Characteristics and the Synergism between Sigma B and Sigma L in Listeria monocytogenes EGD-e" Microorganisms 8, no. 11: 1644. https://doi.org/10.3390/microorganisms8111644