
Pre-Growth Environmental Stresses Affect Foodborne Pathogens Response to Subsequent Chemical Treatments


Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Bacterial Strains and Inoculum Preparation
2.2. Stress Conditions
2.3. Sanitizer Preparation
2.4. Treatment
2.5. Sub-Lethal Injury Determination
2.6. Statistical Analyses
3. Results and Discussion
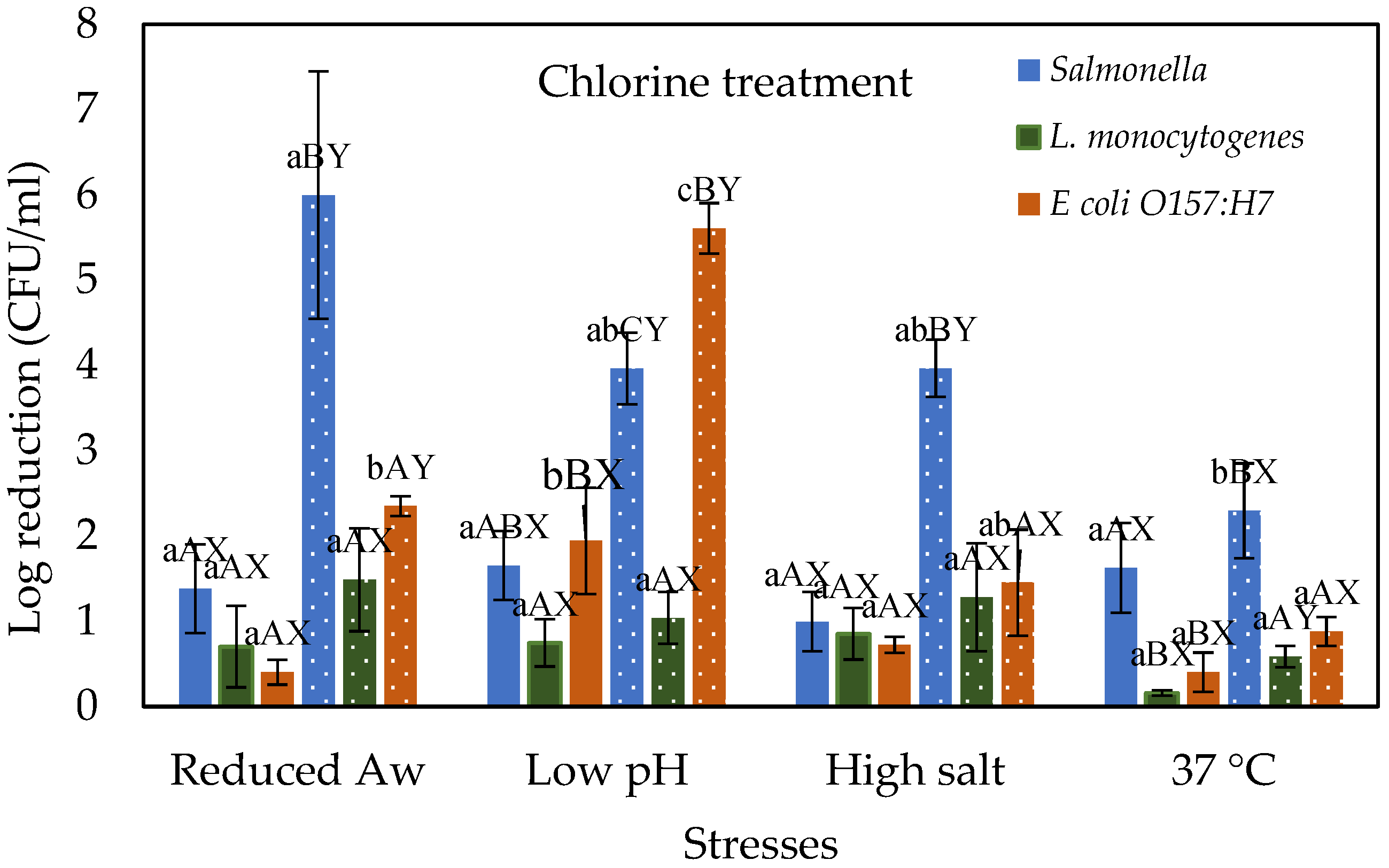
3.1. Stressed Cells Response to Chlorine Treatment
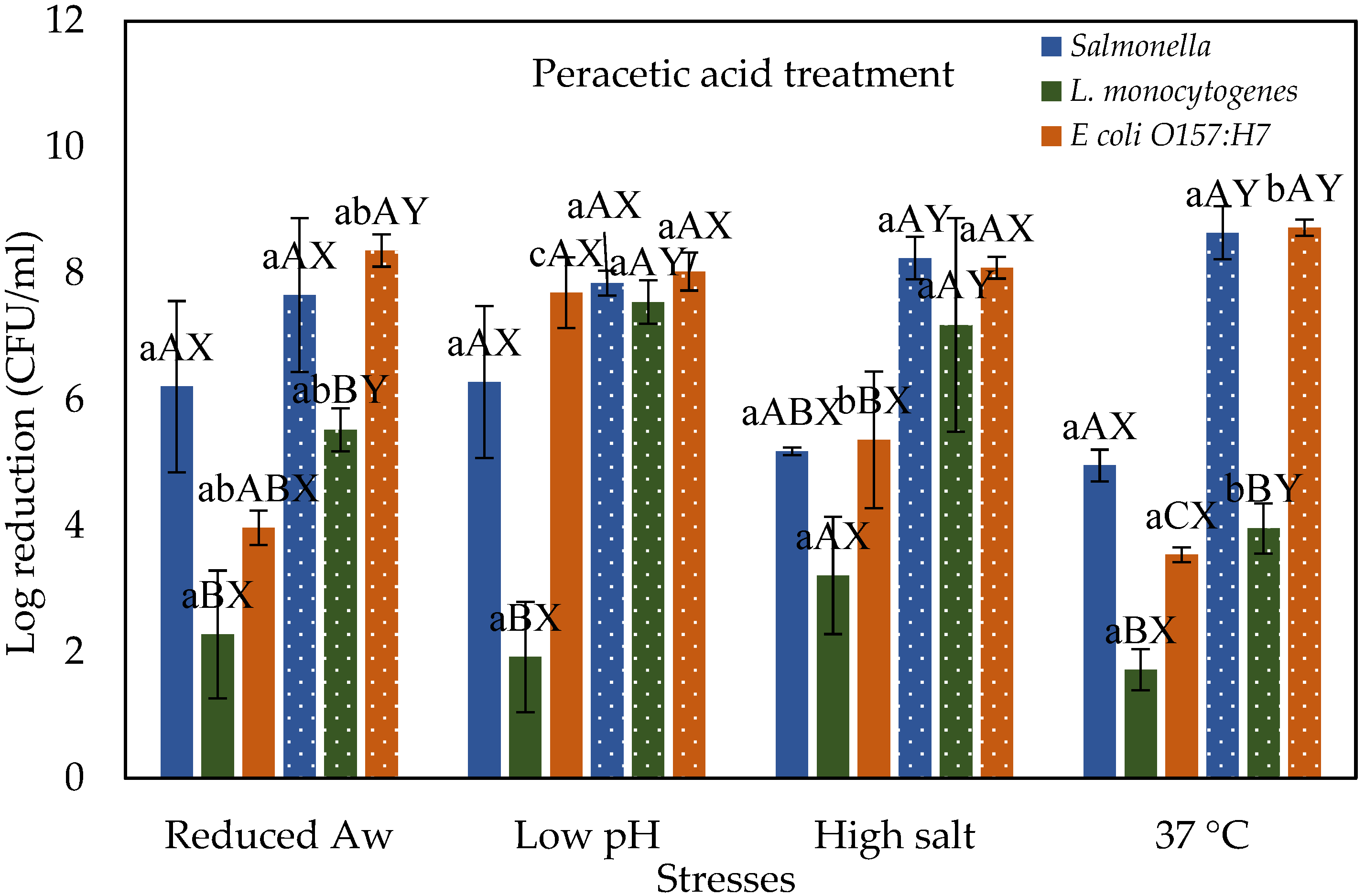
3.2. Stressed Cells Response to Peracetic Acid Treatment
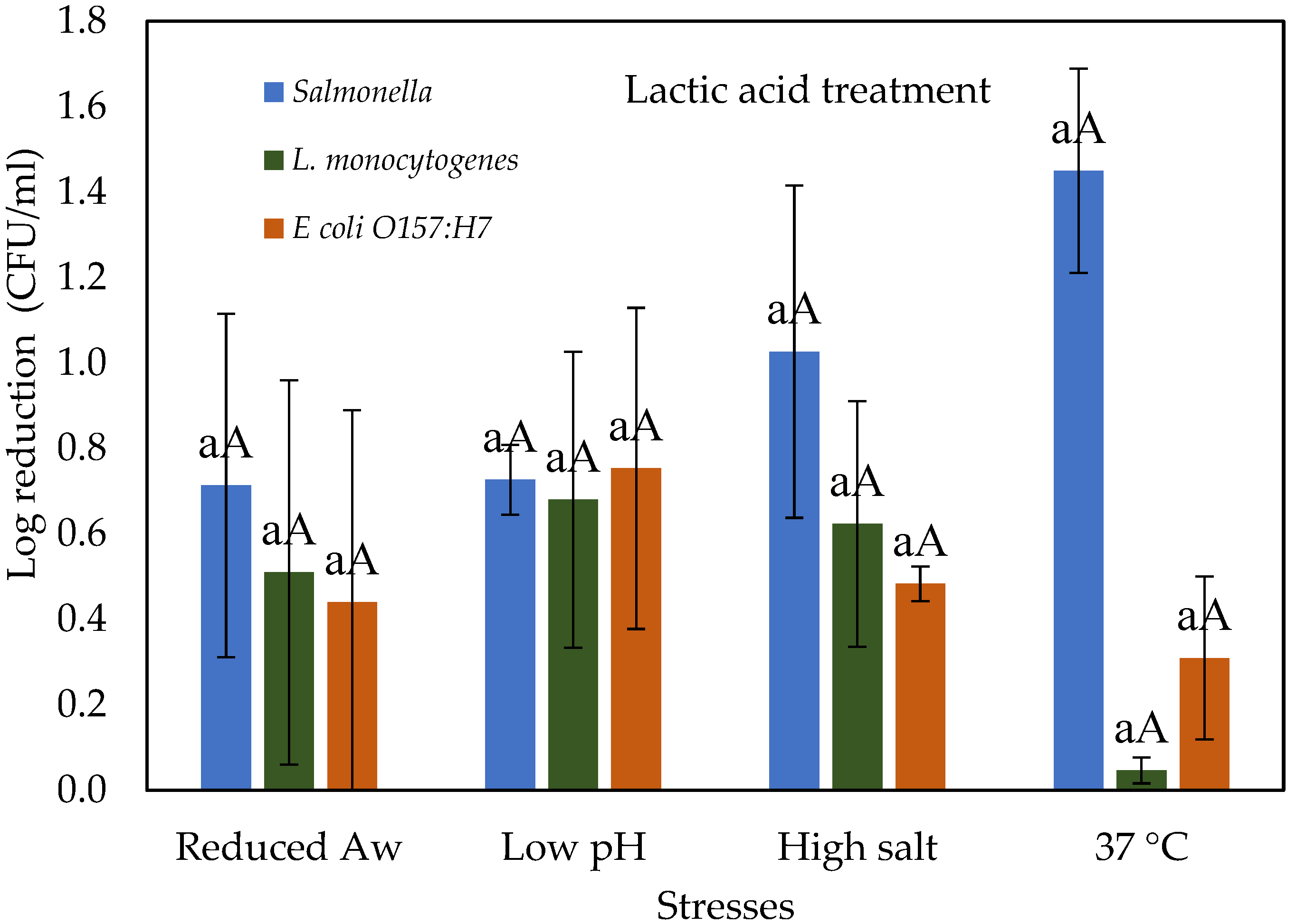
3.3. Stressed Cells Response to Lactic Acid Treatment
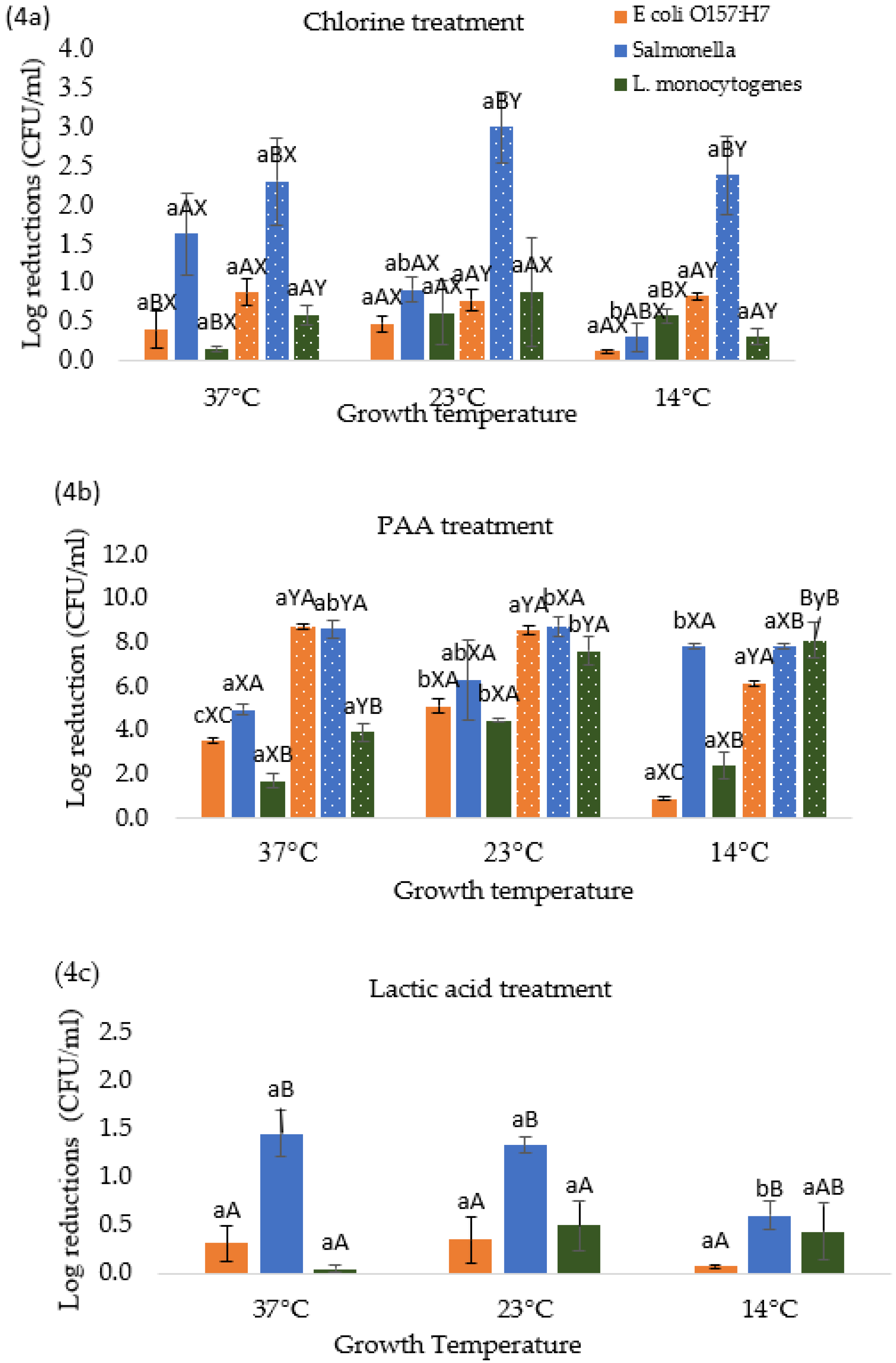
3.4. Effect of Temperature Stresses on Pathogen Response to Different Chemical Treatments
3.5. Sub-Lethal Injury Determination
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carstens, K.C.; Salazar, J.K.; Darkoh, C. Multistate Outbreaks of Foodborne Illness in the United States Associated with Fresh Produce From 2010 to 2017. Front. Microbiol. 2019, 10, 2667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, S.; Sadowsky, M.J. Escherichia coli in the environment: Implications for water quality and human health. Microbes Environ. 2008, 23, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatica, M.K.; Schneider, K.R. Salmonella and produce: Survival in the plant environment and implications in food safety. Virulence 2011, 2, 573–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dygico, L.K.; Gahan, C.G.; Grogan, H.; Burgess, C.M.J. The ability of Listeria monocytogenes to form biofilm on surfaces relevant to the mushroom production environment. Int. J. Food Microbiol. 2020, 317, 108385. [Google Scholar] [CrossRef] [PubMed]
- Dowe, M.J.; Jackson, E.D.; Mori, J.G.; Bell, C.R.J. Listeria monocytogenes survival in soil and incidence in agricultural soils. J. Food Prot. 1997, 60, 1201–1207. [Google Scholar] [CrossRef]
- Islam, M.; Doyle, M.P.; Phatak, S.C.; Millner, P.; Jiang, X.J. Persistence of enterohemorrhagic Escherichia coli O157, H7 in soil and on leaf lettuce and parsley grown in fields treated with contaminated manure composts or irrigation water. J. Food Prot. 2004, 67, 1365–1370. [Google Scholar] [CrossRef] [Green Version]
- Koutsoumanis, K.; Sofos, J. Comparative acid stress response of Listeria monocytogenes, Escherichia coli O157, H7 and Salmonella Typhimurium after habituation at different pH conditions. Lett. Appl. Microbiol. 2004, 38, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Bang, W.; Drake, M.A. Stress response of Escherichia coli. Compr. Rev. Food Sci. Food Saf. 2006, 5, 52–64. [Google Scholar] [CrossRef]
- He, Y.; Li, Y.; Salazar, J.K.; Yang, J.; Tortorello, M.L.; Zhang, W. Increased Water Activity Reduces the Thermal Resistance of Salmonella enterica in Peanut Butter. Appl. Environ. Microbiol. 2013, 79, 4763–4767. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Ross, T.; McMeekin, T.A.; Nichols, P.D. Acid habituation of Escherichia coli and the potential role of cyclopropane fatty acids in low pH tolerance. Int. J. Food Microbiol. 1997, 37, 163–173. [Google Scholar] [CrossRef]
- Lou, Y.; Yousef, A.E. Adaptation to sublethal environmental stresses protects Listeria monocytogenes against lethal preservation factors. Appl. Environ. Microbiol. 1997, 63, 1252–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agranovski, V.; Ristovski, Z.; Hargreaves, M.; Blackall, P.J.; Morawska, L. Performance evaluation of the UVAPS: Influence of physiological age of airborne bacteria and bacterial stress. J. Aerosol Sci. 2003, 34, 1711–1727. [Google Scholar] [CrossRef] [Green Version]
- Jasson, V.; Uyttendaele, M.; Rajkovic, A.; Debevere, J. Establishment of procedures provoking sub-lethal injury of Listeria monocytogenes, Campylobacter jejuni and Escherichia coli O157 to serve method performance testing. Int. J. Food Microbiol. 2007, 118, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Estilo, E.E.C.; Gabriel, A.A. Previous stress exposures influence subsequent UV-C resistance of Salmonella enterica in coconut liquid endosperm. LWT 2017, 86, 139–147. [Google Scholar] [CrossRef]
- Himathongkham, S.; Riemann, H. Destruction of Salmonella typhimurium, Escherichia coli O157, H7 and Listeria monocytogenes in chicken manure by drying and/or gassing with ammonia. FEMS Microbiol. Lett. 1999, 171, 179–182. [Google Scholar] [CrossRef] [Green Version]
- Harrand, A.S.; Guariglia-Oropeza, V.; Skeens, J.; Kent, D.; Wiedmann, M. Nature versus Nurture: Assessing the Impact of Strain Diversity and Pregrowth Conditions on Salmonella enterica, Escherichia coli, and Listeria Species Growth and Survival on Selected Produce Items. Appl. Environ. Microbiol. 2021, 87, e01925-20. [Google Scholar] [CrossRef]
- Gray, M.J.; Wholey, W.-Y.; Jakob, U.J. Bacterial responses to reactive chlorine species. Annu. Rev. Microbiol. 2013, 67, 141–160. [Google Scholar] [CrossRef] [Green Version]
- Dahl, T.A.; Midden, W.; Hartman, P.E. Comparison of killing of gram-negative and gram-positive bacteria by pure singlet oxygen. J. Bacteriol. 1989, 171, 2188–2194. [Google Scholar] [CrossRef] [Green Version]
- Ray, B. Acetic, propionic, and lactic acids of starter culture bacteria as biopreservatives. In Food Biopreservatives of Microbial Origin; CRC Press: Boca Raton, FL, USA, 1992; p. 103. [Google Scholar]
- Ölmez, H.; Temur, S.J. Effects of different sanitizing treatments on biofilms and attachment of Escherichia coli and Listeria monocytogenes on green leaf lettuce. LWT 2010, 43, 964–970. [Google Scholar] [CrossRef]
- To, M.S.; Favrin, S.; Romanova, N.; Griffiths, M.W. Postadaptational resistance to benzalkonium chloride and subsequent physicochemical modifications of Listeria monocytogenes. Appl. Environ. Microbiol. 2002, 68, 5258–5264. [Google Scholar] [CrossRef] [Green Version]
- Bansal, M.; Nannapaneni, R.; Sharma, C.S.; Kiess, A. Listeria monocytogenes response to sublethal chlorine induced oxidative stress on homologous and heterologous stress adaptation. Front. Microbiol. 2018, 9, 2050. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Beuchat, L. Evaluation of sanitizers for killing Escherichia coli O157, H7, Salmonella, and naturally occurring microorganisms on cantaloupes, honeydew melons, and asparagus. Dairy Food Environ. Sanit. 1999, 15, 842–847. [Google Scholar]
- Denyer, S.P.; Stewart, G. Mechanisms of action of disinfectants. Int. Biodeterior. Biodegrad. 1998, 41, 261–268. [Google Scholar] [CrossRef]
- Harrand, A.S.; Kovac, J.; Carroll, L.M.; Guariglia-Oropeza, V.; Kent, D.J.; Wiedmann, M. Assembly and characterization of a pathogen strain collection for produce safety applications: Pre-growth conditions have a larger effect on peroxyacetic acid tolerance than strain diversity. Front. Microbiol. 2019, 10, 1223. [Google Scholar] [CrossRef] [Green Version]
- Beier, R. Interactions and Inhibition of Pathogenic Foodborne Bacteria with Individual Dissociated Organic Acid Species: A Review. J. Food Chem. Nanotechnol. 2021, 7, 4–17. [Google Scholar] [CrossRef]
- Booth, I.R.; Kroll, R.G.; Gould, G.W. The preservation of foods by low pH. In Mechanism of Action of Food Preservation Procedures; Elsevier Science Publishers Ltd.: London, UK, 1989; pp. 119–160. [Google Scholar]
- Sagong, H.-G.; Lee, S.-Y.; Chang, P.-S.; Heu, S.; Ryu, S.; Choi, Y.-J.; Kang, D.-H. Combined effect of ultrasound and organic acids to reduce Escherichia coli O157, H7, Salmonella Typhimurium, and Listeria monocytogenes on organic fresh lettuce. Int. J. Food Microbiol. 2011, 145, 287–292. [Google Scholar] [CrossRef]
- Qiao, Z.-H.; Chen, Y.; Lu, Z.-H.; Guan, L.-J.; Li, L. Antibacterial and bactericide activity of lactic acid against three strains of food-borne pathogenic bacteria. Front. Cell. Infect. Microbiol. 2008, 10, 187–190. [Google Scholar]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Chang, J.-M.; Fang, T.J. Survival of Escherichia coli O157, H7 and Salmonella enterica serovars Typhimurium in iceberg lettuce and the antimicrobial effect of rice vinegar against E. coli O157, H7. Food Microbiol. 2007, 24, 745–751. [Google Scholar] [CrossRef]
- Al-Nabulsi, A.A.; Osaili, T.M.; Obaidat, H.M.; Shaker, R.R.; Awaisheh, S.S.; Holley, R.A. Inactivation of stressed Escherichia coli O157, H7 cells on the surfaces of rocket salad leaves by chlorine and peroxyacetic acid. J. Food Prot. 2014, 77, 32–39. [Google Scholar] [CrossRef]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Akbas, M.; Ölmez, H. Inactivation of Escherichia coli and Listeria monocytogenes on iceberg lettuce by dip wash treatments with organic acids. Lett. Appl. Microbiol. 2007, 44, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Ban, G.-H.; Park, S.-H.; Kim, S.-O.; Ryu, S.; Kang, D.-H. Synergistic effect of steam and lactic acid against Escherichia coli O157, H7, Salmonella Typhimurium, and Listeria monocytogenes biofilms on polyvinyl chloride and stainless steel. Int. J. Food Microbiol. 2012, 157, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Brashears, M.; Amezquita, A.; Stratton, J. Validation of methods used to recover Escherichia coli O157, H7 and Salmonella spp. subjected to stress conditions. J. Food Prot. 2001, 64, 1466–1471. [Google Scholar] [CrossRef]
- Noriega, E.; Velliou, E.; van Derlinden, E.; Mertens, L.; van Impe, J.F. Effect of cell immobilization on heat-induced sublethal injury of Escherichia coli, Salmonella Typhimurium and Listeria innocua. Food Microbiol. 2013, 36, 355–364. [Google Scholar] [CrossRef]
- Finn, S.; Condell, O.; McClure, P.; Amézquita, A.; Fanning, S.J. Mechanisms of survival, responses and sources of Salmonella in low-moisture environments. Front. Microbiol. 2013, 4, 331. [Google Scholar] [CrossRef] [Green Version]
- Gruzdev, N.; Pinto, R.; Sela, S.J. Persistence of Salmonella enterica during dehydration and subsequent cold storage. Food Microbiol. 2012, 32, 415–422. [Google Scholar] [CrossRef]
- Maserati, A.; Fink, R.C.; Lourenco, A.; Julius, M.L.; Diez-Gonzalez, F. General response of Salmonella enterica serovar Typhimurium to desiccation: A new role for the virulence factors sopD and sseD in survival. PLoS ONE 2017, 12, e0187692. [Google Scholar] [CrossRef] [Green Version]
- Kocaman, N.; Sarimehmetoğlu, B. Stress responses of Listeria monocytogenes. Ankara Univ. Vet Fak. Derg. 2016, 63, 421–427. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Organism | Type of Stress Conditions | Growth Conditions | Treatment and % Injury | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| pH | aw | Temp (°C) | Time (h) | Chlorine Treatment | Lactic Acid | Peracetic Acid Treatment | ||||
| 100 ppm | 200 ppm | 0.5% | 40 ppm | 80 ppm | ||||||
| S. enterica | Control | 7.2 | 0.99 | 37 | 18 | 2.80 ± 1.1 | 4.62 ± 2.90 | −4.62 ± 6.15 | 14.78 ± 13.88 | 0.00 ± 0.00 |
| Acid | 5 | 0.99 | 37 | 18 | 6.16 ± 4.32 | 6.15 ± 4.99 | 1.23 ± 0.18 | 16.00 ± 21.33 | 0.00 ± 0.00 | |
| Desiccation | 7.2 | 0.96 | 37 | 18 | 6.95 ± 5.38 | 2.00 ± 1.81 | −0.70 ± 6.96 | 5.95 ± 5.95 | 33.33 ± 57.74 | |
| High salt | 7.3 | 0.99 | 37 | 18 | 2.47 ± 1.64 | 8.45 ± 0.68 | 19.06 ± 7.07 | 1.97 ± 10.11 | 0.00 ± 0.00 | |
| Ambient Temp | 7.2 | 0.99 | 23 | 18 | 2.08 ± 1.32 | 3.77 ± 1.57 | 5.43 ± 6.67 | 17.61 ± 27.70 | 0.00 ± 0.00 | |
| Low Temperature | 7.2 | 0.99 | 14 | 70 | 0.20 ± 2.27 | 3.28 ± 3.32 | −2.07 ± 3.37 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| L. monocytogenes | Control | 7.2 | 0.99 | 37 | 18 | 2.70 ± 0.85 | 4.62 ± 1.28 | 1.61 ± 0.57 | 8.13 ± 6.47 | 64.80 ± 32.25 |
| Acid | 5.5 | 0.99 | 37 | 18 | 3.79 ± 3.16 | 0.81 ± 1.48 | −0.03 ± 3.15 | 58.51 ± 27.03 | 0.00 ± 0.00 | |
| Desiccation | 7.2 | 0.95 | 37 | 18 | 3.67 ± 0.32 | 2.15 ± 4.42 | 1.08 ± 0.96 | 15.62 ± 3.90 | 64.13 ± 6.54 | |
| High salt | 7.3 | 0.99 | 37 | 18 | 1.24 ± 0.86 | 4.07 ± 0.65 | −0.32 ± 1.86 | 9.24 ± 10.33 | 20.11 ± 17.41 | |
| Ambient Temp | 7.2 | 0.99 | 23 | 18 | −0.55 ± 2.32 | 1.29 ± 0.36 | 0.04 ± 2.63 | 7.57 ± 14.45 | 66.67 ± 57.74 | |
| Low Temperature | 7.2 | 0.99 | 14 | 70 | 1.42 ± 0.73 | 5.55 ± 1.06 | 3.10 ± 2.42 | 25.41 ± 16.78 | 66.67 ± 57.74 | |
| E. coli O157:H7 | Control | 7.2 | 0.99 | 37 | 18 | 2.49 ± 1.82 | 3.95 ± 2.25 | 1.85 ± 1.17 | 18.30 ± 21.26 | 0.00 ± 0.00 |
| Acid | 5 | 0.99 | 37 | 18 | 0.12 ± 1.72 | 7.93 ± 3.90 | 0.30 ± 0.30 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Desiccation | 7.2 | 0.96 | 37 | 18 | 4.22 ± 3.06 | 5.17 ± 2.34 | −1.50 ± 1.85 | 12.92 ± 14.57 | 0.00 ± 0.00 | |
| High salt | 7.3 | 0.99 | 37 | 18 | −1.03 ± 0.55 | 20.43 ± 4.02 | −1.16 ± 1.24 | 86.38 ± 23.59 | 0.00 ± 0.00 | |
| Ambient Temp | 7.2 | 0.99 | 23 | 18 | 0.60 ± 1.02 | 0.85 ± 1.56 | 0.98 ± 1.09 | 1.65 ± 3.12 | 0.00 ± 0.00 | |
| Low Temperature | 7.2 | 0.99 | 14 | 70 | 0.28 ± 0.10 | 0.24 ± 0.60 | 0.50 ± 0.62 | 3.30 ± 2.81 | 32.52 ± 15.17 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.; Yemmireddy, V. Pre-Growth Environmental Stresses Affect Foodborne Pathogens Response to Subsequent Chemical Treatments. Microorganisms 2022, 10, 786. https://doi.org/10.3390/microorganisms10040786
Singh A, Yemmireddy V. Pre-Growth Environmental Stresses Affect Foodborne Pathogens Response to Subsequent Chemical Treatments. Microorganisms. 2022; 10(4):786. https://doi.org/10.3390/microorganisms10040786
Chicago/Turabian StyleSingh, Amandeep, and Veerachandra Yemmireddy. 2022. "Pre-Growth Environmental Stresses Affect Foodborne Pathogens Response to Subsequent Chemical Treatments" Microorganisms 10, no. 4: 786. https://doi.org/10.3390/microorganisms10040786