Porcine Colostrum Protects the IPEC-J2 Cells and Piglet Colon Epithelium against Clostridioides (syn. Clostridium) difficile Toxin-Induced Effects

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. C. difficile Toxins
2.3. Porcine Colostrum
2.4. IPEC-J2 Cell Culture Conditions
2.5. Treatment of IPEC-J2 Cells with Toxins and Porcine Colostrum
2.6. TEER Measurement of IPEC-J2 Cells
2.7. Immunofluorescence Microscopy of IPEC-J2 Cells
2.8. IPEC-J2 Cell Viability
2.9. Animals
2.10. Ussing Chamber Assay
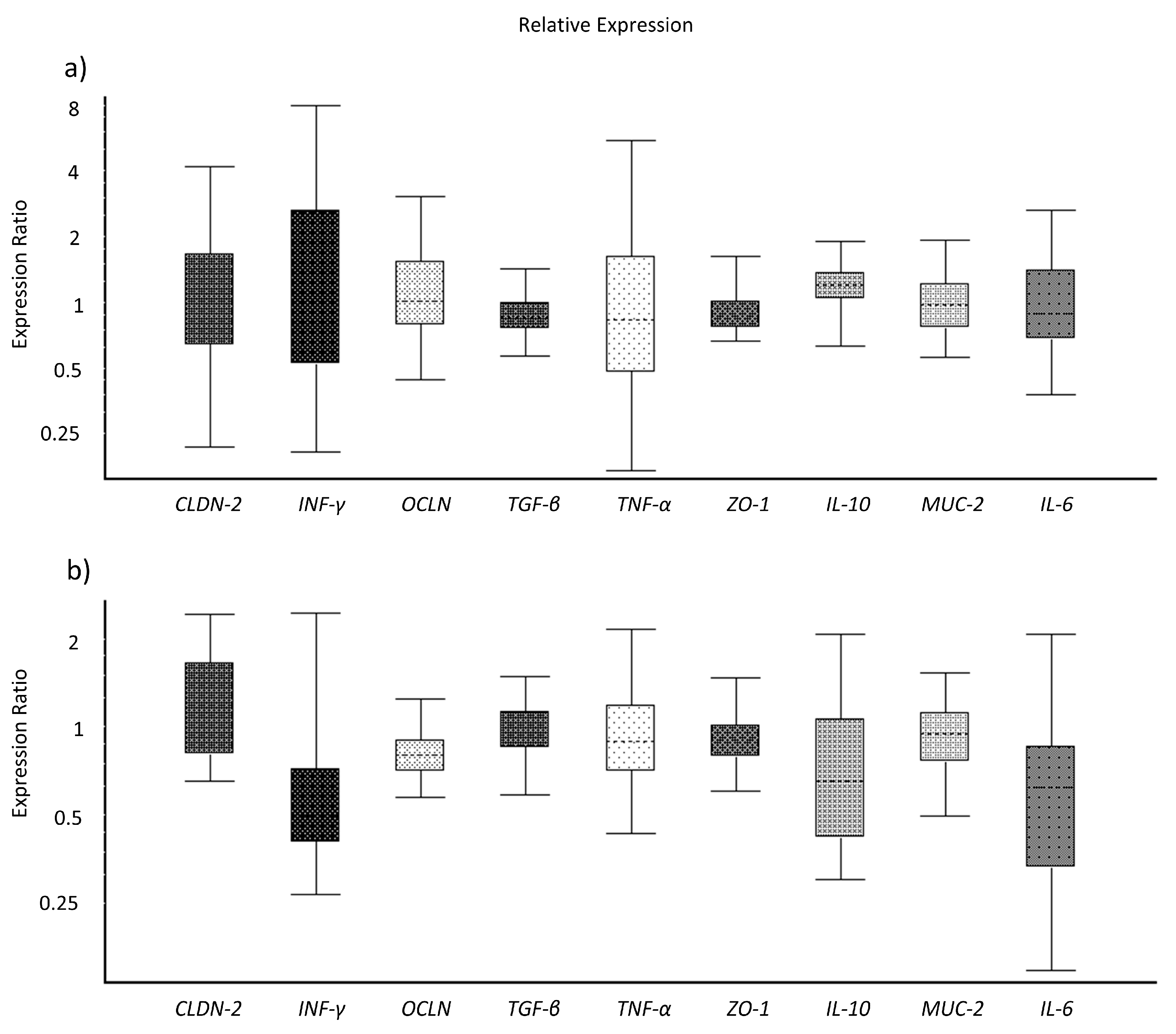
2.11. Gene Expression in IPEC-J2 Cells and in Colon Tissues
2.12. Statistics
3. Results
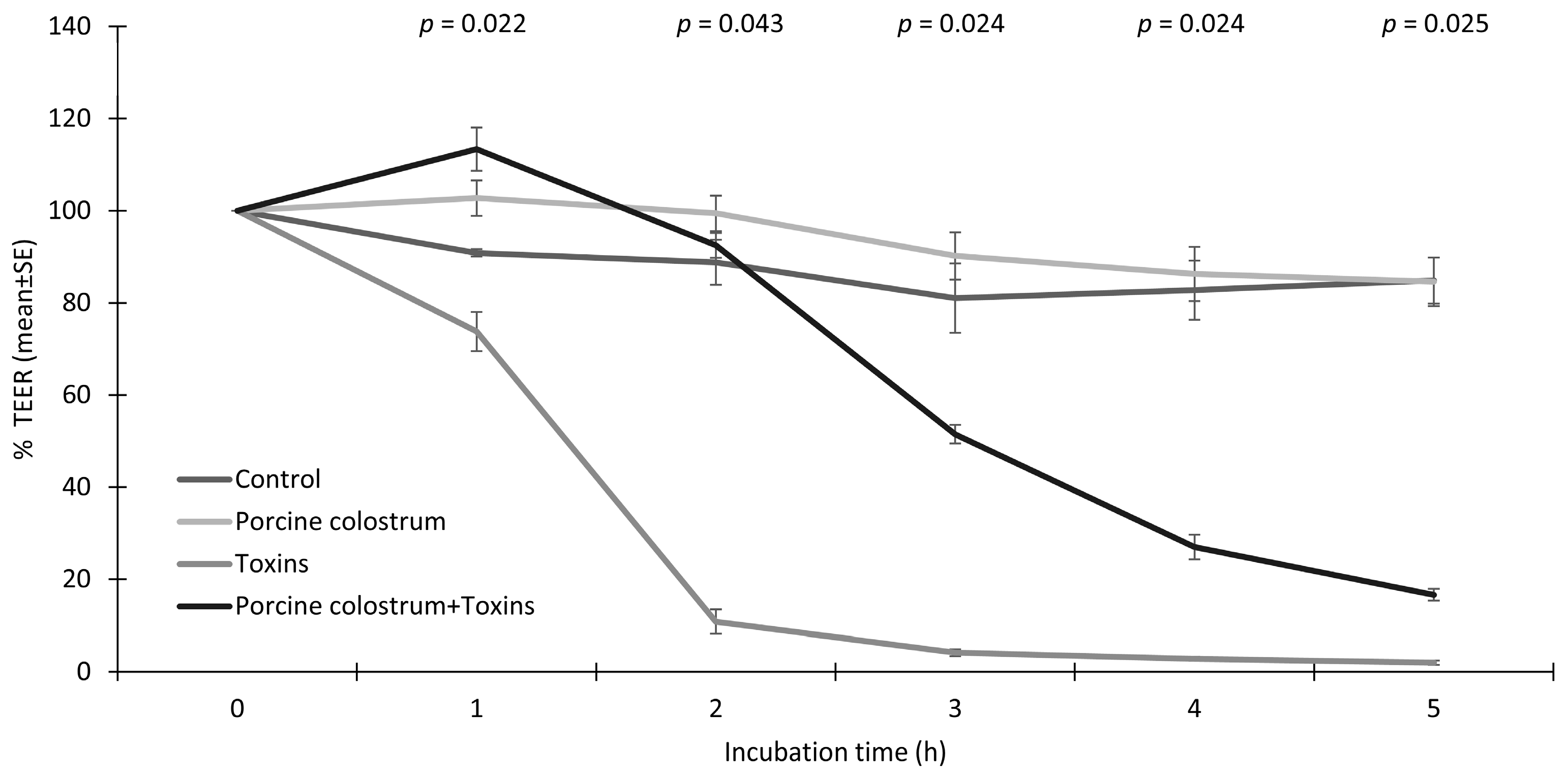
3.1. Porcine Colostrum Improves the Integrity of IPEC-J2 Cells and Protects from Toxin-Induced Effects
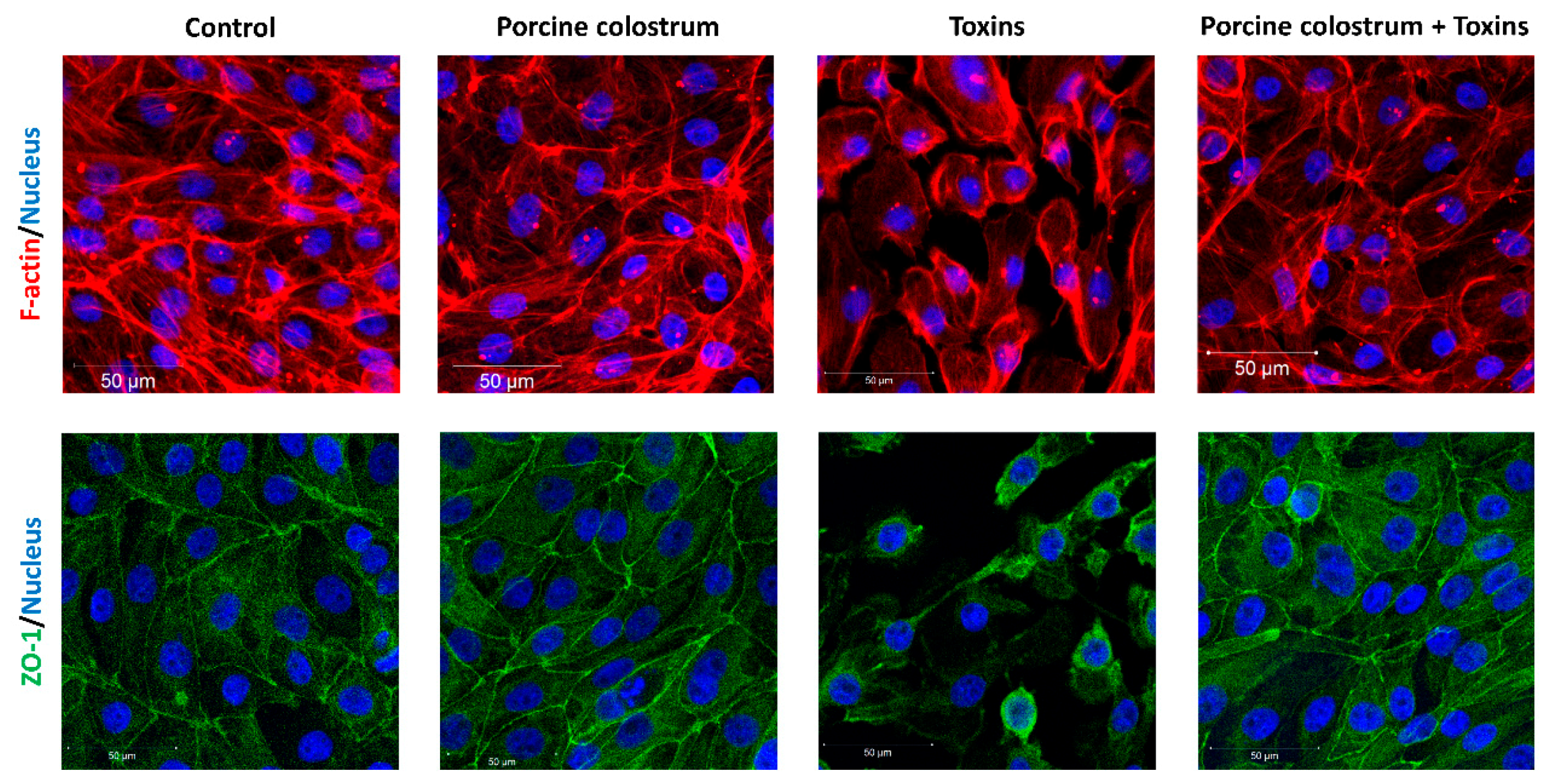
3.2. Porcine Colostrum Protects Cell Structures against C. difficile Toxins
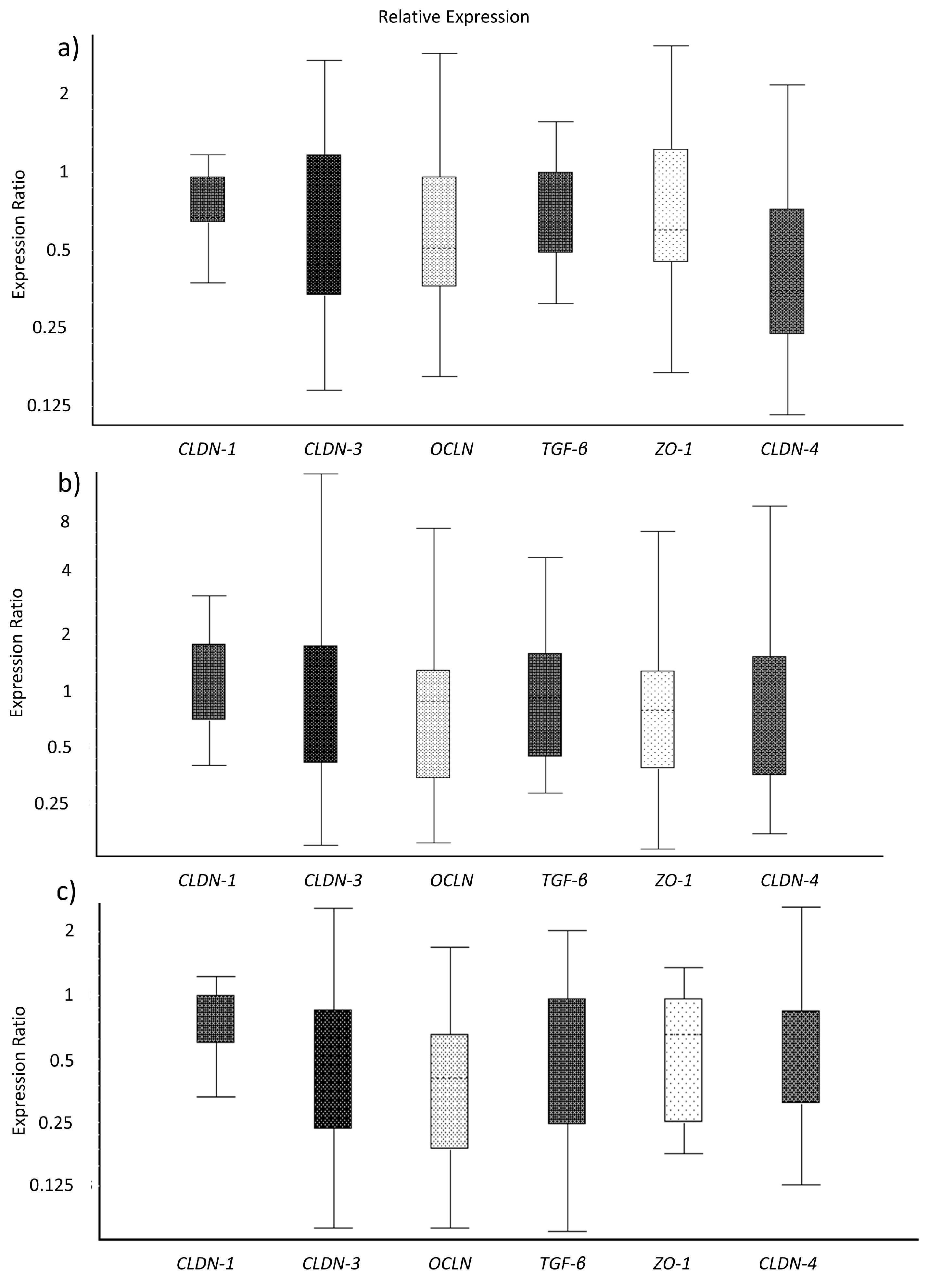
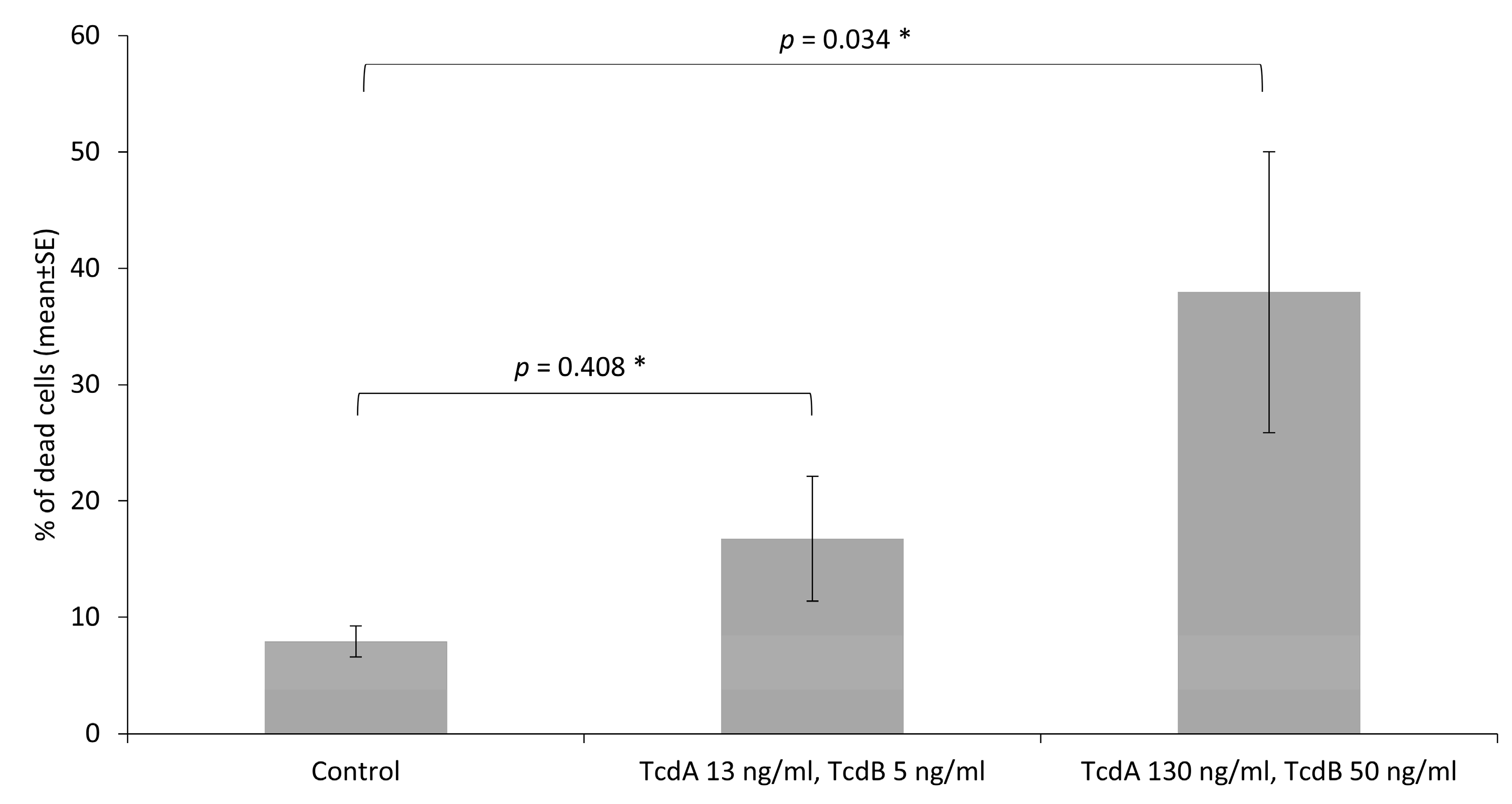
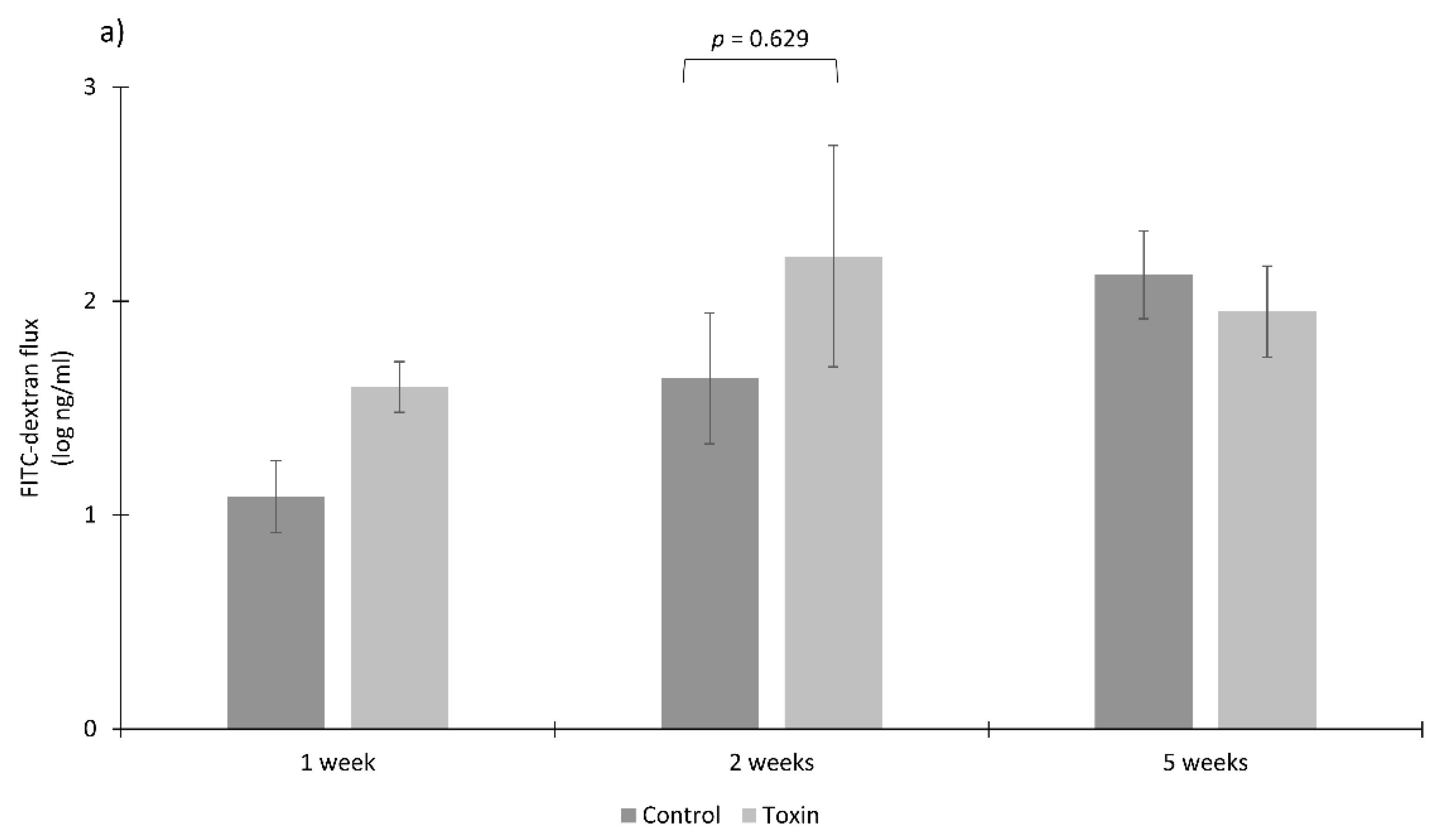
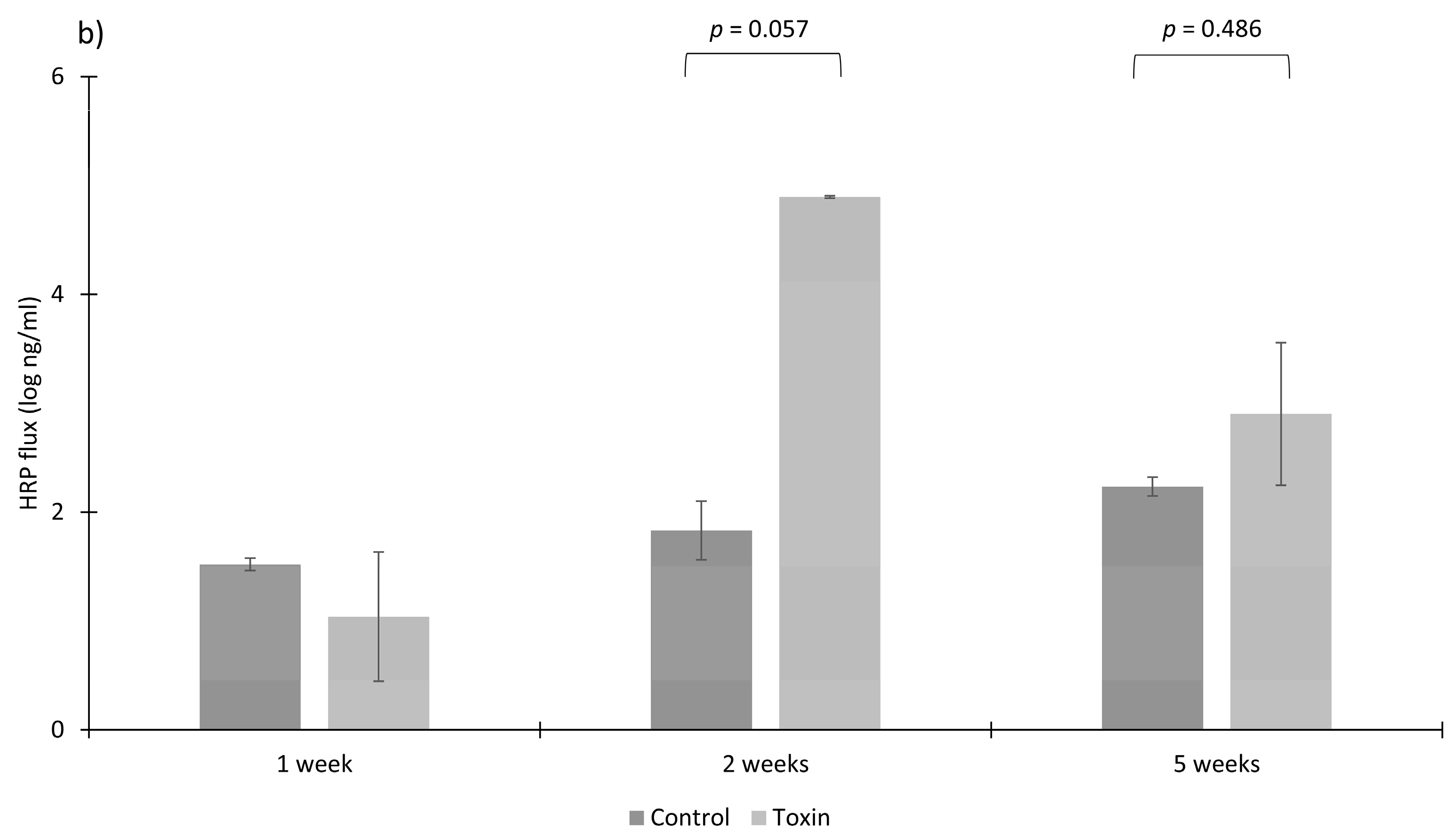
3.3. Toxins Reduce IPEC-J2 Cell Viability and Increase Permeability of Colon Epithelia
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Zhang, J.; Liu, B.; Dai, Y.; Han, C.; Wang, J. Diversity analysis of intestinal microflora between healthy and diarrheal neonatal piglets from the same litter in different regions. Anaerobe 2018, 55, 136–141. [Google Scholar]
- Moono, P.; Putsathit, P.; Knight, D.R.; Squire, M.M.; Hampson, D.J.; Foster, N.F.; Riley, T.V. Persistence of Clostridium difficile RT 237 infection in a Western Australian piggery. Anaerobe 2015, 37, 62–66. [Google Scholar] [CrossRef]
- Voth, D.E.; Ballard, J.D. Clostridium difficile toxins: Mechanism of action and role in disease. Clin. Microbiol. Rev. 2005, 18, 247–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusrat, A.; Turner, J.R.; Verkade, P.; Madara, L.; Parkos, C.A. Clostridium difficile Toxins Disrupt Epithelial Barrier Function by Altering Membrane Microdomain Localization of Tight Junction Proteins. Infect. Immun. 2001, 69, 1329–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grześkowiak, Ł.M.; Pieper, R.; Huynh, H.A.; Cutting, S.M.; Vahjen, W.; Zentek, J. Impact of early-life events on the susceptibility to Clostridium difficile colonisation and infection in the offspring of the pig. Gut Microbes 2019, 10, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, G.; Pothoulakis, C.; LaMont, J.T.; Madara, J.L. Clostridium difficile toxin A perturbs cytoskeletal structure and tight junction permeability of cultured human intestinal epithelial monolayers. J. Clin. Invest. 1988, 82, 1516–1524. [Google Scholar] [CrossRef]
- Nam, H.J.; Kang, J.K.; Kim, S.K.; Ahn, K.J.; Seok, H.; Park, S.J.; Chang, J.S.; Pothoulakis, C.; Lamont, J.T.; Kim, H. Clostridium difficile toxin A decreases acetylation of tubulin, leading to microtubule depolymerization through activation of histone deacetylase 6, and this mediates acute inflammation. J. Biol. Chem. 2010, 285, 32888–32896. [Google Scholar] [CrossRef] [Green Version]
- Grześkowiak, Ł.; Martínez-Vallespín, B.; Dadi, T.H.; Radloff, J.; Amasheh, S.; Heinsen, F.-A.; Franke, A.; Reinert, K.; Vahjen, W.; Zentek, J.; et al. Formula Feeding Predisposes Neonatal Piglets to Clostridium difficile Gut Infection. J. Infect. Dis. 2018, 217, 1442–1452. [Google Scholar] [CrossRef]
- Waters, E.H.; Orr, J.P.; Clark, E.G.; Schaufele, C.M. Typhlocolitis Caused by Clostridium Difficile in Suckling Piglets. J. Vet. Diagnostic Investig. 1998, 10, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Squire, M.M.; Carter, G.P.; Mackin, K.E.; Chakravorty, A.; Norén, T.; Elliott, B.; Lyras, D.; Riley, T.V. Novel molecular type of clostridium difficile in neonatal pigs, Western Australia. Emerg. Infect. Dis. 2013, 19, 790–792. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Zentek, J.; Vahjen, W. Determination of the extent of Clostridium difficile colonisation and toxin accumulation in sows and neonatal piglets. Anaerobe 2016, 40, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Inman, C.F.; Haverson, K.; Konstantinov, S.R.; Jones, P.H.; Harris, C.; Smidt, H.; Miller, B.; Bailey, M.; Stokes, C. Rearing environment affects development of the immune system in neonates. Clin. Exp. Immunol. 2010, 160, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Balzola, F.; Bernstein, C.; Ho, G.T.; Lees, C. Environmentally-acquired bacteria influence microbial diversity and natural innate immune responses at gut surfaces: Commentary. Inflamm. Bowel Dis. Monit. 2010, 10, 134. [Google Scholar]
- Radloff, J.; Zakrzewski, S.S.; Pieper, R.; Markov, A.G.; Amasheh, S. Porcine milk induces a strengthening of barrier function in porcine jejunal epithelium in vitro. Ann. N. Y. Acad. Sci. 2017, 1397, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Debast, S.B.; van Leengoed, L.A.M.G.; Goorhuis, A.; Harmanus, C.; Kuijper, E.J.; Bergwerff, A.A. Clostridium difficile PCR ribotype 078 toxinotype V found in diarrhoeal pigs identical to isolates from affected humans. Environ. Microbiol. 2009, 11, 505–511. [Google Scholar] [CrossRef]
- Karlsson, S.; Burman, L.G.; Åkerlund, T. Induction of toxins in Clostridium difficile is associated with dramatic changes of its metabolism. Microbiology 2008, 154, 3430–3436. [Google Scholar] [CrossRef] [Green Version]
- Grześkowiak, Ł.; Riedmüller, J.; de Thomasson, H.; Bordessoule, S.; Seyboldt, C.; Zentek, J.; Vahjen, W. Porcine and bovine Clostridium difficile ribotype 078 isolates demonstrate similar growth and toxigenic properties. Int. Microbiol. 2018. [Google Scholar] [CrossRef]
- Martínez-Vallespín, B.; Vahjen, W.; Zentek, J. Effects of medium-chain fatty acids on the structure and immune response of IPEC-J2 cells. Cytotechnology 2016, 68, 1925–1936. [Google Scholar] [CrossRef] [Green Version]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef]
- Kröger, S.; Pieper, R.; Schwelberger, H.G.; Wang, J.; Villodre Tudela, C.; Aschenbach, J.R.; Van Kessel, A.G.; Zentek, J. Diets high in heat-treated soybean meal reduce the histamine-induced epithelial response in the colon of weaned piglets and increase epithelial catabolism of histamine. PLoS ONE 2013, 8, e80612. [Google Scholar] [CrossRef]
- Pieper, R.; Scharek-Tedin, L.; Zetzsche, A.; Röhe, I.; Kröger, S.; Vahjen, W.; Zentek, J. Bovine milk–based formula leads to early maturation-like morphological, immunological, and functional changes in the jejunum of neonatal piglets. J. Anim. Sci. 2016, 94, 989–999. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Pieper, R.; Rieger, J.; Vahjen, W.; Davin, R.; Plendl, J.; Meyer, W.; Zentek, J. Effect of dietary zinc oxide on morphological characteristics, mucin composition and gene expression in the colon of weaned piglets. PLoS ONE 2014, 9, e91091. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Klobasa, F.; Werhahn, E.; Butler, J.E. Composition of sow milk during lactation. J. Anim. Sci. 1987, 64, 1458–1466. [Google Scholar] [CrossRef]
- Klobasa, F.; Butler, J.E. Absolute and relative concentrations of immunoglobulins G, M, and A, and albumin in the lacteal secretion of sows of different lactation numbers. Am. J. Vet. Res. 1987, 48, 176–182. [Google Scholar]
- Huygelen, V.; De Vos, M.; Willemen, S.; Tambuyzer, B.; Casteleyn, C.; Knapen, D.; Van Ginneken, C.; Van Cruchten, S. Increased intestinal barrier function in the small intestine of formula-fed neonatal piglets. J. Anim. 2012, 90, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Chiba, H.; Osanai, M.; Murata, M.; Kojima, T.; Sawada, N. Transmembrane proteins of tight junctions. Biochim. Biophys. Acta Biomembr. 2008, 1778, 588–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, J.; Feng, H.; Parry, N.; Tzipori, S. Piglet models of acute or chronic Clostridium difficile illness. J. Infect. Dis. 2010, 201, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieper, R.; Vahjen, W.; Zentek, J. Intestinal lactose and mineral concentration affect the microbial ecophysiology along the gastrointestinal tract of formula-fed neonatal piglets. J. Anim. Sci. 2016, 94, 3786–3795. [Google Scholar] [CrossRef] [PubMed]
- Vergauwen, H. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Chapter 12; pp. 125–134. [Google Scholar]
- Hockley, S.L.; Arlt, V.M.; Brewer, D.; Giddings, I.; Phillips, D.H. Time- and concentration-dependent changes in gene expression induced by benzo(a)pyrene in two human cell lines, MCF-7 and HepG2. BMC Genomics 2006, 7, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Persson, A. Clinical Assessment of Udder Health Status of Sows at Time of Weaning with Special Reference to Bacteriology and Cytology in Milk. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 1997, 44, 143–158. [Google Scholar] [CrossRef] [PubMed]
- El-Hawiet, A.; Kitova, E.N.; Kitov, P.I.; Eugenio, L.; Ng, K.K.S.; Mulvey, G.L.; Dingle, T.C.; Szpacenko, A.; Armstrong, G.D.; Klassen, J.S. Binding of Clostridium difficile toxins to human milk oligosaccharides. Glycobiology 2011, 21, 1217–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, J.; Mukherjee, J.; Parry, N.; Tzipori, S. Antibody against TcdB, but not TcdA, prevents development of gastrointestinal and systemic clostridium difficile disease. J. Infect. Dis. 2013, 207, 323–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, O.R.; Steele, J.A.; Zhang, Q.; Schmidt, D.J.; Wang, Y.; Hamel, P.E.S.; Beamer, G.; Xu, B.; Tzipori, S. Systemically Administered IgG Anti-Toxin Antibodies Protect the Colonic Mucosa during Infection with Clostridium difficile in the Piglet Model. PLoS ONE 2014, 9, e111075. [Google Scholar] [CrossRef] [PubMed]
- Babcock, G.J.; Broering, T.J.; Hernandez, H.J.; Mandell, R.B.; Donahue, K.; Boatright, N.; Stack, A.M.; Lowy, I.; Graziano, R.; Molrine, D.; et al. Human monoclonal antibodies directed against toxins A and B prevent Clostridium difficile-induced mortality in hamsters. Infect. Immun. 2006, 74, 6339–6347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, C.D.; Mahoney, M.V. Bezlotoxumab for the prevention of clostridium difficile infection: A review of current evidence and safety profile. Infect. Drug Resist. 2019, 12, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weese, J.S.; Wakeford, T.; Reid-Smith, R.; Rousseau, J.; Friendship, R. Longitudinal investigation of Clostridium difficile shedding in piglets. Anaerobe 2010, 16, 501–504. [Google Scholar] [CrossRef]
- Kyne, L.; Warny, M.; Qamar, A.; Kelly, C.P. Asymptomatic carriage of Clostridium difficile and serum levels of IgG antibody against toxin A. N. Engl. J. Med. 2000, 342, 390–397. [Google Scholar] [CrossRef]
- Markowska-Daniel, I.; Pomorska-Mól, M. Shifts in immunoglobulins levels in the porcine mammary secretions during whole lactation period. Bull. Vet. Inst. Pulawy 2010, 54, 345–349. [Google Scholar]
- Devillers, N.; Farmer, C.; Le Dividich, J.; Prunier, A. Variability of colostrum yield and colostrum intake in pigs. Animal 2007, 1, 1033–1041. [Google Scholar] [CrossRef] [Green Version]
- Richard, O.K.; Grahofer, A.; Nathues, H.; Posthaus, H. Vaccination against Clostridium perfringens type C enteritis in pigs: A field study using an adapted vaccination scheme. Porc. Heal. Manag. 2019, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Grönlund, M.M.; Arvilommi, H.; Kero, P.; Lehtonen, O.P.; Isolauri, E. Importance of intestinal colonisation in the maturation of humoral immunity in early infancy: A prospective follow up study of healthy infants aged 0–6 months. Arch. Dis. Child. Fetal Neonatal Ed. 2000, 83, F186–F192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönlund, M.; Grzeskowiak, Ł.; Isolauri, E.; Salminen, S. Influence of mother’s intestinal microbiota on gut colonization in the infant. Gut Microbes 2011, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Šinkora, M.; Sun, J.; Šinkorová, J.; Christenson, R.K.; Ford, S.P.; Butler, J.E. Antibody Repertoire Development in Fetal and Neonatal Piglets. VI. B Cell Lymphogenesis Occurs at Multiple Sites with Differences in the Frequency of In-frame Rearrangements. J. Immunol. 2003, 170, 1781–1788. [Google Scholar] [CrossRef] [Green Version]
- Grześkowiak, Ł.; Dadi, T.H.; Zentek, J.; Vahjen, W. Developing Gut Microbiota Exerts Colonisation Resistance to Clostridium (syn. Clostridioides) difficile in Piglets. Microorganisms 2019, 7, 218. [Google Scholar] [CrossRef] [Green Version]
- Antharam, V.C.; Li, E.C.; Ishmael, A.; Sharma, A.; Mai, V.; Rand, K.H.; Wang, G.P. Intestinal dysbiosis and depletion of butyrogenic bacteria in Clostridium difficile infection and nosocomial diarrhea. J. Clin. Microbiol. 2013, 51, 2884–2892. [Google Scholar] [CrossRef] [Green Version]
- Kociolek, L.K.; Espinosa, R.O.; Gerding, D.N.; Hauser, A.R.; Ozer, E.A.; Budz, M.; Balaji, A.; Chen, X.; Tanz, R.R.; Yalcinkaya, N.; et al. Natural Clostridioides difficile Toxin Immunization in Colonized Infants. Clin. Infect. Dis. 2019, 60611, 1–8. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grześkowiak, Ł.; Pieper, R.; Kröger, S.; Martínez-Vallespín, B.; Hauser, A.E.; Niesner, R.; Vahjen, W.; Zentek, J. Porcine Colostrum Protects the IPEC-J2 Cells and Piglet Colon Epithelium against Clostridioides (syn. Clostridium) difficile Toxin-Induced Effects. Microorganisms 2020, 8, 142. https://doi.org/10.3390/microorganisms8010142
Grześkowiak Ł, Pieper R, Kröger S, Martínez-Vallespín B, Hauser AE, Niesner R, Vahjen W, Zentek J. Porcine Colostrum Protects the IPEC-J2 Cells and Piglet Colon Epithelium against Clostridioides (syn. Clostridium) difficile Toxin-Induced Effects. Microorganisms. 2020; 8(1):142. https://doi.org/10.3390/microorganisms8010142
Chicago/Turabian StyleGrześkowiak, Łukasz, Robert Pieper, Susan Kröger, Beatriz Martínez-Vallespín, Anja E. Hauser, Raluca Niesner, Wilfried Vahjen, and Jürgen Zentek. 2020. "Porcine Colostrum Protects the IPEC-J2 Cells and Piglet Colon Epithelium against Clostridioides (syn. Clostridium) difficile Toxin-Induced Effects" Microorganisms 8, no. 1: 142. https://doi.org/10.3390/microorganisms8010142