Absence of Viable Toxoplasma gondii in Artisanal Raw-Milk Ewe Cheese Derived from Naturally Infected Animals

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Collection and Cheesemaking
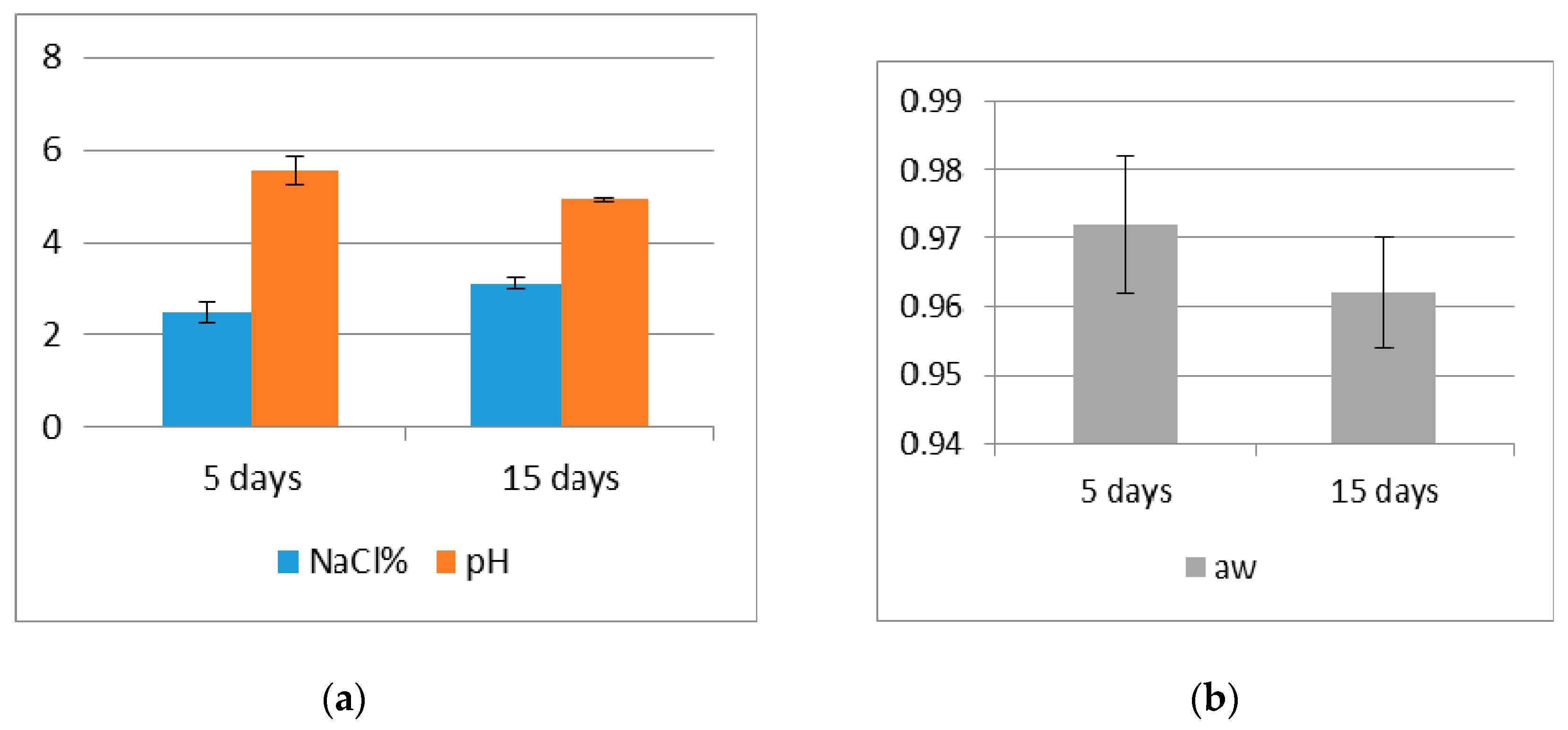
2.2. pH, Water Activity and NaCl Measurement
2.3. Purification of Total DNA and RNA from Milk and Cheese Samples
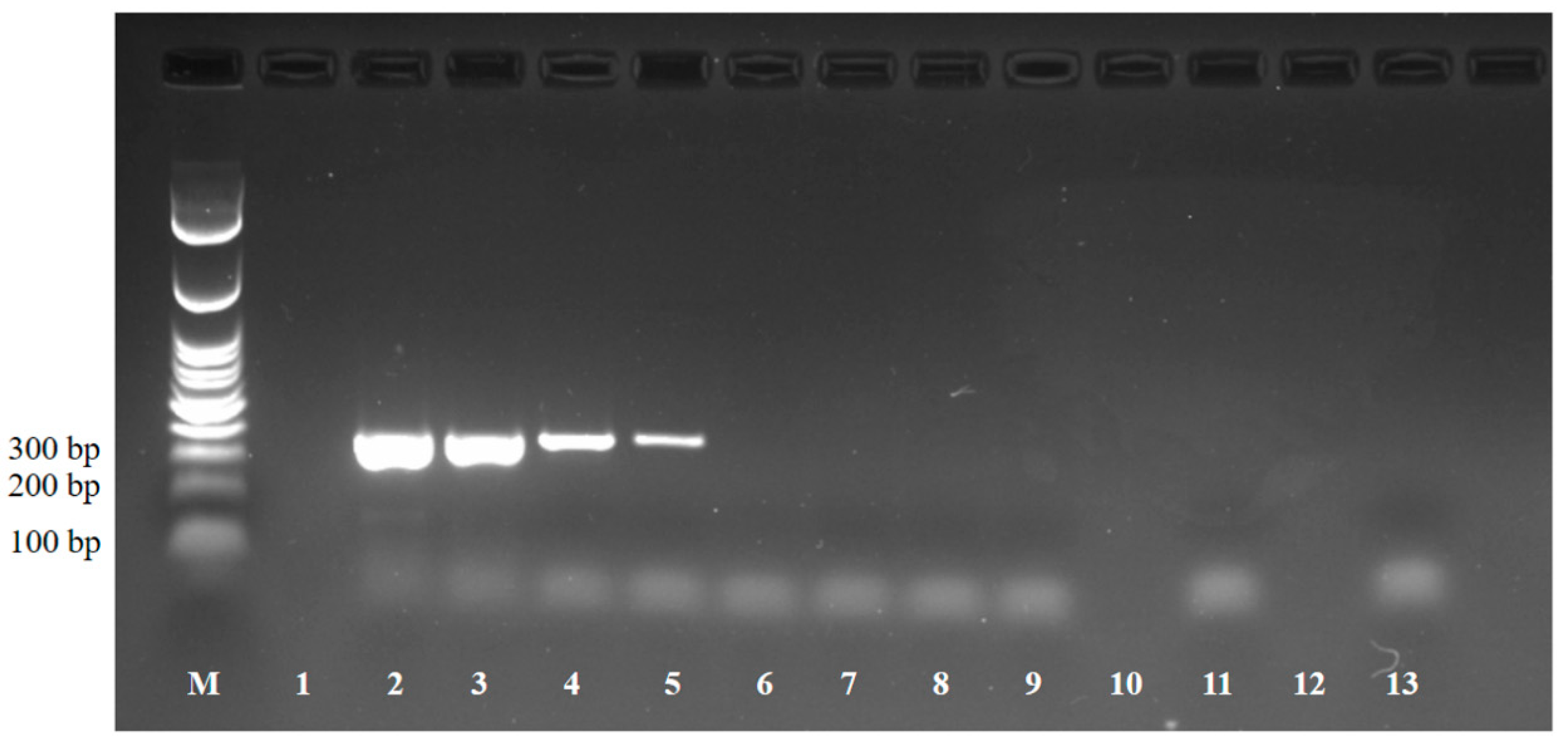
2.4. Loop Mediated Isothermal Amplification (LAMP)
2.5. Reverse Transcriptase (RT)-PCR
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Dubey, J.P. General biology. In Toxoplasmosis of Animals and Humans, 2nd ed.; Dubey, J.P., Ed.; CDC Press: Boca Raton, FL, USA, 2010; pp. 1–72. [Google Scholar]
- Dubey, J.P.; Jones, J.L. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol. 2008, 38, 1257–1278. [Google Scholar] [CrossRef] [PubMed]
- Iacobucci, E.; Taus, N.S.; Ueti, M.W.; Sukhbaatar, L.; Bastsukh, Z.; Papageorgiou, S.; Fritz, H. Detection and genotypic characterization of Toxoplasma gondii DNA within the milk of Mongolian livestock. Parasitol. Res. 2019, 118, 2005–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk factors for Toxoplasma gondii infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Alvarado-Esquivel, C.; Liesenfeld, O.; Torres-Castorena, A.; Estrada-Martinez, S.; Urbina-Alvarez, J.D.; Ramos-de la Rocha, M.; Marquez-Conde, J.A.; Dubey, J.P. Seroepidemiology of Toxoplasma gondii infection in patients with vision and hearing impairments, cancer, HIV, or undergoing hemodialysis in Durango, Mexico. J. Parasitol. 2010, 96, 505–508. [Google Scholar] [CrossRef]
- Belluco, S.; Simonato, G.; Mancin, M.; Pietrobelli, M.; Ricci, A. Toxoplasma gondii infection and food consumption: A systematic review and meta-analysis of case-controlled studies. Crit. Rev. Food Sci. Nutr. 2018, 58, 3085–3096. [Google Scholar] [CrossRef]
- Walsh, C.P.; Hammond, S.E.; Zajac, A.M.; Lindsay, D.S. Survival of Toxoplasma gondii tachyzoites in goat milk: Potential source of human toxoplasmosis. J. Eukaryot. Microbiol. 1999, 46, 73S–74S. [Google Scholar]
- Cook, A.J.C.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of toxoplasma infection in pregnant women: European multicenter case-control study. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Pereira, K.S.; Franco, R.M.; Leal, D.A. Transmission of toxoplasmosis (Toxoplasma gondii) by foods. In Advances in Food and Nutrition Research; Taylor, S.E., Ed.; Academic Press: Burlington, MA, USA, 2010; Volume 60, pp. 1–19. [Google Scholar]
- Boughattas, S. Commentary on: Detection of Toxoplasma gondii in raw caprine, ovine, buffalo, bovine, and camel milk using cell cultivation, cat bioassay, capture ELISA, and PCR methods in Iran. Front. Microbiol. 2015, 6, 215. [Google Scholar] [CrossRef]
- EFSA, European Food Safety Agency. Scientific opinion on the public health risks related to the consumption of raw drinking milk. EFSA J. 2015, 13, 3940. [Google Scholar] [CrossRef] [Green Version]
- Branciari, R.; Ranucci, D.; Trabalza-Marinucci, M.; Codini, M.; Orru, M.; Ortenzi, R.; Valiani, A. Evaluation of the antioxidant properties and oxidative stability of Pecorino cheese made from the raw milk of ewes fed Rosmarinus officinalis L. leaves. Int. J. Food. Sci. Technol. 2015, 50, 558–565. [Google Scholar] [CrossRef]
- Hiramoto, R.M.; Mayrbaurl-Borges, M.; Galisteo, A.J., Jr.; Meireles, L.R.; Macre, M.S.; Andrade, H.F. Infectivity of cysts of the ME-49 Toxoplasma gondii strain in bovine milk and homemade cheese. Rev. Saude Publica 2001, 35, 113–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, J.P.; Verma, S.K.; Ferreira, L.R.; Oliveira, S.; Cassinelli, A.B.; Ying, Y.; Jones, J.L. Detection and survival of Toxoplasma gondii in milk and cheese from experimentally infected sheep. J. Food Protect. 2014, 77, 1747–1753. [Google Scholar] [CrossRef] [PubMed]
- Tavares, T.; Malcata, F.X. Alternative dairy products made with raw milk. In Raw Milk; Nero, L.A., Fernandes De Carvalho, A., Eds.; Academic Press: Oxford, UK, 2019; pp. 223–234. [Google Scholar]
- Veronesi, F.; Chiesa, F.; Zanet, S.; Battisti, E.; Sebastiani, V.; Branciari, R.; Valiani, A.; Ranucci, D. Screening of Toxoplasma gondii positive sheep flocks in Perugia Province using bulk milk analyses. LAR 2018, 24, 185–187. [Google Scholar]
- Association of Official Analytical Chemists. AOAC method 935.43. In Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 2000. [Google Scholar]
- Mura, M.C.; Daga, C.; Bodano, S.; Paludo, M.; Luridiana, S.; Pazzola, M.; Dettori, M.L.; Vacca, G.M.; Carcangiu, V. Development of a RNA extraction method from milk for gene expression study in the mammary gland of sheep. Mol. Biol. Rep. 2013, 40, 2169–2173. [Google Scholar] [CrossRef]
- Krasteva, D.; Toubiana, M.; Hartati, S.; Kusumawati, A.; Dubremetz, J.F.; Widada, J.S. Development of loop-mediated isothermal amplification (LAMP) as a diagnostic tool of toxoplasmosis. Vet. Parasitol. 2009, 162, 327–331. [Google Scholar] [CrossRef]
- Trisciuoglio, A.; Zanet, S.; Marello, G.; Chiesa, F.; Nucera, D.M.; Bergallo, M.; Gennero, M.S.; Ferroglio, E. The use of loop-mediated isothermal amplification improves Toxoplasma gondii detection in wildlife. J. Vet. Diagn. Invest. 2015, 27, 754–757. [Google Scholar] [CrossRef] [Green Version]
- Mahittikorn, A.; Wickert, H.; Sukthana, Y. Toxoplasma gondii: Simple duplex RT-PCR assay for detecting SAG1 and BAG1 genes during stage conversion in immunosuppressed mice. Exp. Parasitol. 2010, 124, 225–231. [Google Scholar] [CrossRef]
- Bezerra, M.J.; Kim, P.C.; Moraes, É.P.; Sá, S.G.; Albuquerque, P.P.; Silva, J.G.; Alves, B.H.; Mota, R.A. Detection of Toxoplasma gondii in the milk of naturally infected goats in the Northeast of Brazil. Transbound. Emerg. Dis. 2015, 62, 421–424. [Google Scholar] [CrossRef]
- Branciari, R.; Valiani, A.; Trabalza Marinucci, M.; Miraglia, D.; Ranucci, D.; Acuti, G.; Esposto, S.; Mughetti, L. Consumer acceptability of ovine cheese from ewes fed extruded linseed-enriched diet. Small Ruminant Res. 2012, 106, S43–S48. [Google Scholar] [CrossRef]
- Cultrera, R.; Seraceni, S.; Contini, C. Efficacy of a novel reverse transcriptase-polymerase chain reaction (RT-PCR) for detecting Toxoplasma gondii bradyzoite gene expression in human clinical specimens. Mol. Cell Probes 2002, 16, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Travaillé, E.; La Carbona, S.; Gargala, G.; Aubert, D.; Guyot, K.; Dumètre, A.; Villena, I.; Houssin, M. Development of a qRT-PCR method to assess the viability of Giardia intestinalis cysts, Cryptosporidium spp. and Toxoplasma gondii oocysts. Food Control 2016, 59, 359–365. [Google Scholar] [CrossRef]
- Rousseau, A.; La Carbona, S.; Dumètre, A.; Robertson, L.J.; Gargala, G.; Escotte-Binet, S.; Favennec, L.; Villena, I.; Gerard, C.; Aubert, D. Assessing viability and infectivity of foodborne and waterborne stages (cysts/oocysts) of Giardia duodenalis, Cryptosporidium spp., and Toxoplasma gondii: A review of methods. Parasite 2018, 25, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas, E.N.; Augustine, S.A.; Villegas, L.F.; Ware, M.W.; See, M.J.; Lindquist, H.A.; Dubey, J.P. Using quantitative reverse transcriptase PCR and cell culture plaque assays to determine resistance of Toxoplasma gondii oocysts to chemical sanitizers. J. Microbiol. Meth. 2010, 81, 219–225. [Google Scholar] [CrossRef]
- Ware, M.W.; Augustine, S.A.; Erisman, D.O.; See, M.J.; Wymer, L.; Hayes, S.L.; Ayes, S.L.; Dubey, J.P.; Villegas, E.N. Determining UV inactivation of Toxoplasma gondii oocysts by using cell culture and a mouse bioassay. Appl. Environ. Microbiol. 2010, 76, 5140–5147. [Google Scholar] [CrossRef] [Green Version]
- Habtewold, T.; Groom, Z.; Duchateau, L.; Christophides, G.K. Detection of viable Plasmodium ookinetes in the midguts of Anopheles coluzzi using PMA-qrtPCR. Parasites Vectors 2015, 8, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Fredericks, J.; Hawkins-Cooper, D.S.; Hill, D.E.; Luchansky, J.; Porto-Fett, A.; Gamble, H.R.; Fournet, V.M.; Urban, J.F.; Holley, R.; Dubey, J.P. Low salt exposure results in inactivation of Toxoplasma gondii bradyzoites during formulation of dry cured ready-to-eat pork sausage. Food Waterborne Parasitol. 2019, 12, e00047. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Toxoplasma gondii-Positive 1 | Milk | Cheese | |
|---|---|---|---|
| 5 Days of Ripening | 15 Days of Ripening | ||
| LAMP | 16/16 | 32/32 | 32/32 |
| RT-PCR | 16/16 | 0/32 | 0/32 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranucci, D.; Battisti, E.; Veronesi, F.; Diaferia, M.; Morganti, G.; Branciari, R.; Ferroglio, E.; Valiani, A.; Chiesa, F. Absence of Viable Toxoplasma gondii in Artisanal Raw-Milk Ewe Cheese Derived from Naturally Infected Animals. Microorganisms 2020, 8, 143. https://doi.org/10.3390/microorganisms8010143
Ranucci D, Battisti E, Veronesi F, Diaferia M, Morganti G, Branciari R, Ferroglio E, Valiani A, Chiesa F. Absence of Viable Toxoplasma gondii in Artisanal Raw-Milk Ewe Cheese Derived from Naturally Infected Animals. Microorganisms. 2020; 8(1):143. https://doi.org/10.3390/microorganisms8010143
Chicago/Turabian StyleRanucci, David, Elena Battisti, Fabrizia Veronesi, Manuela Diaferia, Giulia Morganti, Raffaella Branciari, Ezio Ferroglio, Andrea Valiani, and Francesco Chiesa. 2020. "Absence of Viable Toxoplasma gondii in Artisanal Raw-Milk Ewe Cheese Derived from Naturally Infected Animals" Microorganisms 8, no. 1: 143. https://doi.org/10.3390/microorganisms8010143