Isolation and Identification of Microvirga thermotolerans HR1, a Novel Thermo-Tolerant Bacterium, and Comparative Genomics among Microvirga Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Strain Isolation, and Cultivation
2.2. 16S rRNA and Housekeeping Gene Amplification and Analysis
2.3. Physiological Characteristics of Microvirga thermotolerans HR1
2.4. Chemotaxonomic Analysis of Microvirga thermotolerans HR1
2.5. Complete Genome Sequencing and Analysis
2.6. Comparative Genomics of Microvirga Species
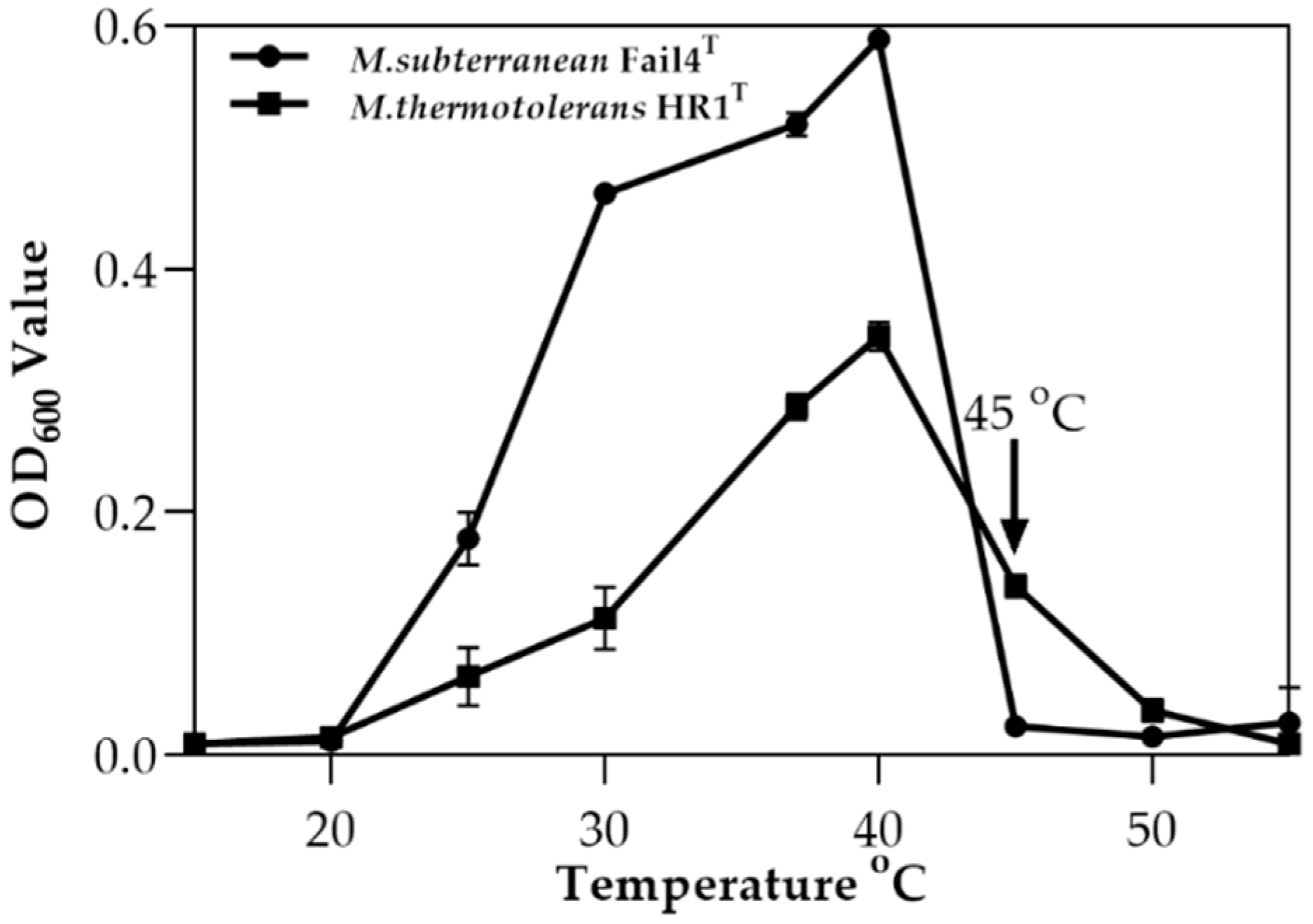
2.7. Effect of Temperature on the Growth of Microvirga thermotolerans HR1
3. Results
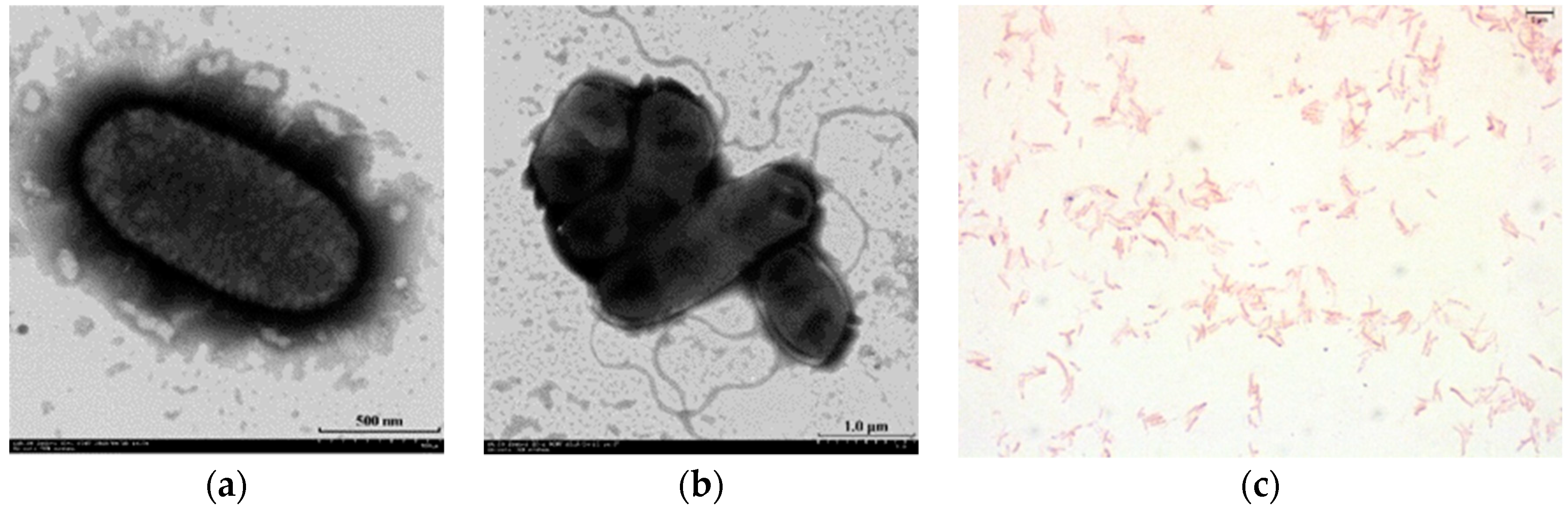
3.1. Isolation and Characterization of Microvirga thermotolerans HR1
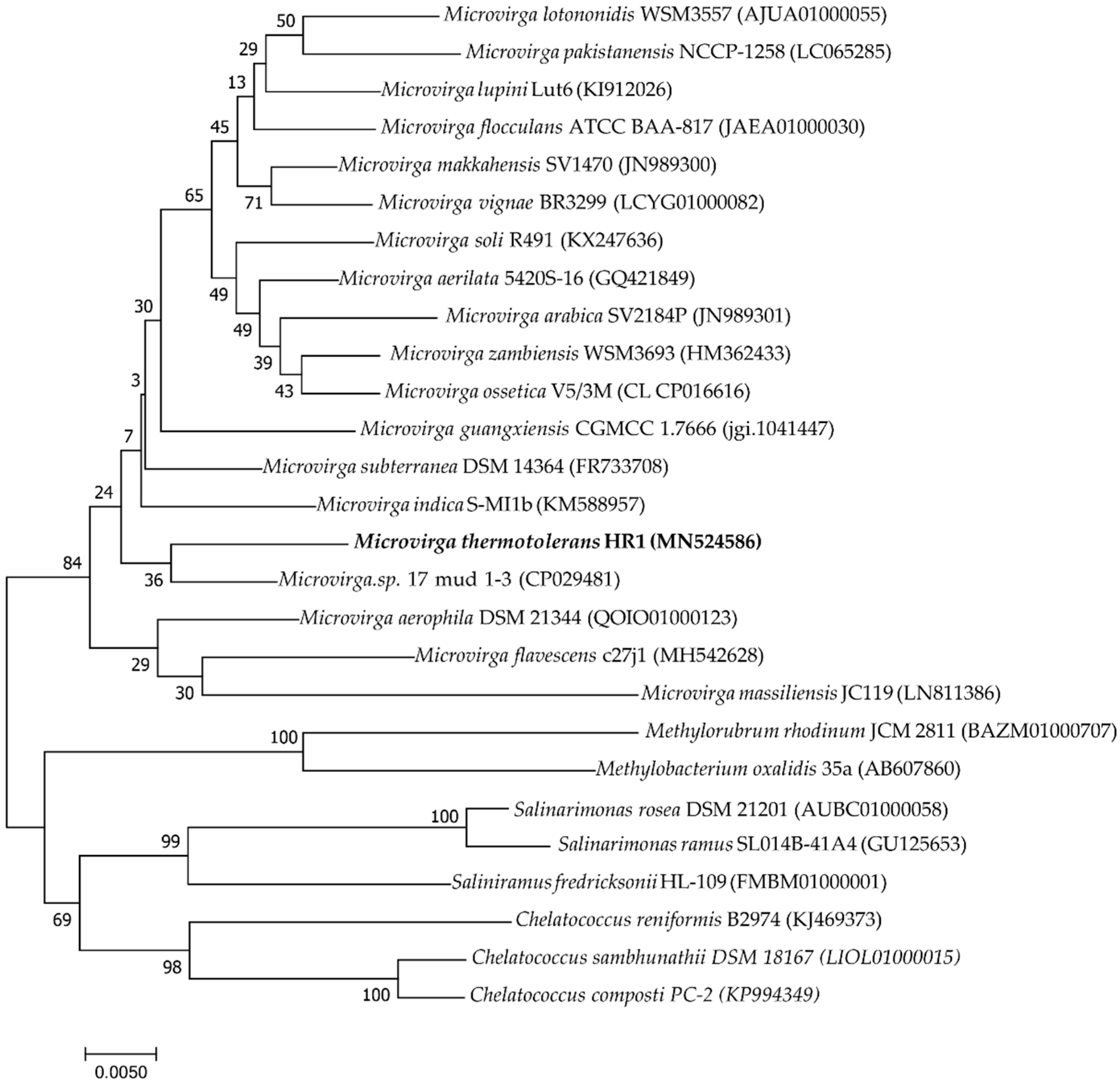
3.2. Phylogenic Analysis of 16S rRNA and Housekeeping Genes
3.3. Genomic Features of Microvirga thermotolerans HR1 Strain
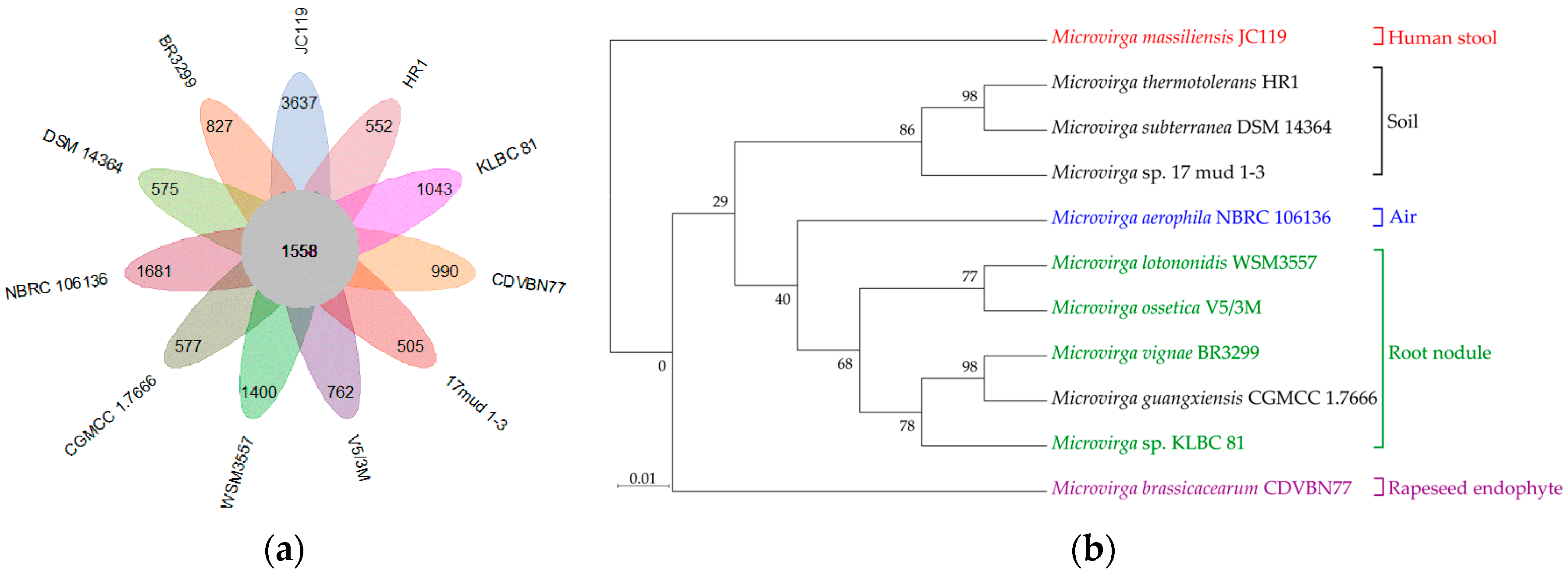
3.4. Niche Adaption of Microvirga Species
3.5. Description of Microvirga thermotolerans sp. nov.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Song, F.; Xin, Y.H.; Zhang, J.; Fang, C. Microvirga guangxiensis sp. nov., a novel alphaproteobacterium from soil, and emended description of the genus Microvirga. Int. J. Syst. Evol. Microbiol. 2009, 59, 1997–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanso, S.; Patel, B.K. Microvirga subterranea gen. nov., sp. nov., a moderate thermophile from a deep subsurface Australian thermal aquifer. Int. J. Syst. Evol. Microbiol. 2003, 53, 401–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weon, H.Y.; Kwon, S.W.; Son, J.A.; Jo, E.H.; Kim, S.J.; Kim, Y.S.; Kim, B.Y.; Ka, J.O. Description of Microvirga aerophila sp. nov. and Microvirga aerilata sp. nov., isolated from air, reclassification of Balneimonas flocculans Takeda et al. 2004 as Microvirga flocculans comb. nov. and emended description of the genus Microvirga. Int. J. Syst. Evol. Microbiol. 2010, 60, 2596–2600. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Zhang, J.; Yao, Q.; Feng, G.D.; Zhu, H.H. Microvirga flavescens sp. nov., a novel bacterium isolated from forest soil and emended description of the genus Microvirga. Int. J. Syst. Evol. Microbiol. 2019, 69, 667–671. [Google Scholar] [CrossRef]
- Huq, M.A. Microvirga rosea sp. nov.: A nanoparticle producing bacterium isolated from soil of rose garden. Arch. Microbiol. 2018, 200, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Tapase, S.R.; Mawlankar, R.B.; Sundharam, S.S.; Krishnamurthi, S.; Dastager, S.G.; Kodam, K.M. Microvirga indica sp. nov., an arsenite-oxidizing Alphaproteobacterium, isolated from metal industry waste soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 3525–3531. [Google Scholar] [CrossRef]
- Dahal, R.H.; Kim, J. Microvirga soli sp. nov., an alphaproteobacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 127–132. [Google Scholar] [CrossRef]
- Amin, A.; Ahmed, I.; Habib, N.; Abbas, S.; Hasan, F.; Xiao, M.; Hozzein, W.N.; Li, W.J. Microvirga pakistanensis sp. nov., a novel bacterium isolated from desert soil of Cholistan, Pakistan. Arch. Microbiol. 2016, 198, 933–939. [Google Scholar] [CrossRef]
- Takeda, M.; Suzuki, I.; Koizumi, J. Balneomonas flocculans gen. nov., sp. nov., a new cellulose-producing member of the alpha-2 subclass of Proteobacteria. Syst. Appl. Microbiol. 2004, 27, 139–145. [Google Scholar] [CrossRef]
- Ardley, J.K.; Parker, M.A.; De Meyer, S.E.; Trengove, R.D.; O’Hara, G.W.; Reeve, W.G.; Yates, R.J.; Dilworth, M.J.; Willems, A.; Howieson, J.G. Microvirga lupini sp. nov., Microvirga lotononidis sp. nov. and Microvirga zambiensis sp. nov. are alphaproteobacterial root-nodule bacteria that specifically nodulate and fix nitrogen with geographically and taxonomically separate legume hosts. Int. J. Syst. Evol. Microbiol. 2012, 62, 2579–2588. [Google Scholar] [CrossRef]
- Safronova, V.I.; Kuznetsova, I.G.; Sazanova, A.L.; Belimov, A.A.; Andronov, E.E.; Chirak, E.R.; Osledkin, Y.S.; Onishchuk, O.P.; Kurchak, O.N.; Shaposhnikov, A.I.; et al. Microvirga ossetica sp. nov., a species of rhizobia isolated from root nodules of the legume species Vicia alpestris Steven. Int. J. Syst. Evol. Microbiol. 2017, 67, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Radl, V.; Simoes-Araujo, J.L.; Leite, J.; Passos, S.R.; Martins, L.M.; Xavier, G.R.; Rumjanek, N.G.; Baldani, J.I.; Zilli, J.E. Microvirga vignae sp. nov., a root nodule symbiotic bacterium isolated from cowpea grown in semi-arid Brazil. Int. J. Syst. Evol. Microbiol. 2014, 64, 725–730. [Google Scholar] [CrossRef]
- Caputo, A.; Lagier, J.C.; Azza, S.; Robert, C.; Mouelhi, D.; Fournier, P.E.; Raoult, D. Microvirga massiliensis sp. nov., the human commensal with the largest genome. Microbiologyopen 2016, 5, 307–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Gomez, A.; Saati-Santamaria, Z.; Igual, J.M.; Rivas, R.; Mateos, P.F.; Garcia-Fraile, P. Genome Insights into the Novel Species Microvirga brassicacearum, a Rapeseed Endophyte with Biotechnological Potential. Microorganisms 2019, 7, 354. [Google Scholar] [CrossRef] [Green Version]
- Sathiyaraj, G.; Kim, M.K.; Kim, J.-Y.; Kim, S.-J.; Jang, J.H.; Maeng, S.; Kang, M.-S.; Srinivasan, S. Complete genome sequence of Microvirga sp. 17 mud 1–3, a radiation-resistant bacterium. Mol. Cell. Toxicol. 2018, 14, 347–352. [Google Scholar] [CrossRef]
- Zilli, J.E.; Passos, S.R.; Leite, J.; Xavier, G.R.; Rumjaneck, N.G.; Simoes-Araujo, J.L. Draft Genome Sequence of Microvirga vignae Strain BR 3299T, a Novel Symbiotic Nitrogen-Fixing Alphaproteobacterium Isolated from a Brazilian Semiarid Region. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBio-Cloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Buck, J.D. Nonstaining (KOH) method for determination of gram reactions of marine bacteria. Appl. Environ. Microbiol. 1982, 44, 992–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA—An ultra-fast pan-genome analysis pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Salaemae, W.; Booker, G.W.; Polyak, S.W. The Role of Biotin in Bacterial Physiology and Virulence: a Novel Antibiotic Target for Mycobacterium tuberculosis. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Choby, J.E.; Skaar, E.P. Heme Synthesis and Acquisition in Bacterial Pathogens. J. Mol. Biol. 2016, 428, 3408–3428. [Google Scholar] [CrossRef] [Green Version]
- Bobay, L.M.; Ochman, H. The Evolution of Bacterial Genome Architecture. Front. Genet. 2017, 8, 72. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Kwok, L.Y.; Hou, Q.; Sun, Y.; Li, W.; Zhang, H.; Sun, Z. Comparative genomic analysis revealed great plasticity and environmental adaptation of the genomes of Enterococcus faecium. BMC Genom. 2019, 20, 602. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Varghese, N.J.; Mukherjee, S.; Ivanova, N.; Konstantinidis, K.; Mavrommatis, K.; Kyrpides, N.C.; Pati, A. Microbial species delineation using whole genome sequences. Nucleic Acids Res. 2015, 43, 6761–6771. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A place for dna-dna reassociation and 16s rrna sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Noll, K.M. Thermophilic Bacteria. In Encyclopedia of Genetics; Sydney, B., Jefferey, H.M., Eds.; Molecular Sciences Institute Inc.: Berkeley, CA, USA, 2001; pp. 1961–1963. [Google Scholar]
- Schmid, A.K.; Howell, H.A.; Battista, J.R.; Peterson, S.N.; Lidstrom, M.E. Global transcriptional and proteomic analysis of the Sig1 heat shock regulon of Deinococcus radiodurans. J. Bacteriol. 2005, 187, 3339–3351. [Google Scholar] [CrossRef] [Green Version]
- Pitsikas, P.; Polosina, Y.Y.; Cupples, C.G. Interaction between the mismatch repair and nucleotide excision repair pathways in the prevention of 5-azacytidine-induced CG-to-GC mutations in Escherichia coli. DNA Repair 2009, 8, 354–359. [Google Scholar] [CrossRef]
- Yang, B.; Zheng, J.; Brown, E.W.; Zhao, S.; Meng, J. Characterisation of antimicrobial resistance-associated integrons and mismatch repair gene mutations in Salmonella serotypes. Int. J. Antimicrob. Agents 2009, 33, 120–124. [Google Scholar] [CrossRef]
- Handa, N.; Morimatsu, K.; Lovett, S.T.; Kowalczykowski, S.C. Reconstitution of initial steps of dsDNA break repair by the RecF pathway of E. coli. Genes Dev. 2009, 23, 1234–1245. [Google Scholar] [CrossRef] [Green Version]
- Ling, J.; Wang, H.; Wu, P.; Li, T.; Tang, Y.; Naseer, N.; Zheng, H.; Masson-Boivin, C.; Zhong, Z.; Zhu, J. Plant nodulation inducers enhance horizontal gene transfer of Azorhizobium caulinodans symbiosis island. Proc. Natl. Acad. Sci. USA 2016, 113, 13875–13880. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Isolation source | Rice paddy | Forest soil | Soil | Thermal aquifer |
| Cell dimensions (µm) | 0.5–0.9 × 1.2–2.8 | 0.4–0.7 × 1.8–4.2 | 0.6–0.65 × 0.9–1.2 | 1.06 × 1.5–4.0 |
| Motility | + | + | + | + |
| Conditions for growth: | ||||
| Temperature range (°C) | 15–50 | 15–37 | 25–45 | 25–45 |
| Optimum temperature (°C) | 40 | 30 | 40 | 41 |
| NaCl tolerance (%, w/v) | 0–0.5 | 0–1 | 0–7 | <1.0 |
| pH range | 6–8 | 6–10 | 6–12 | 6–9 |
| Nitrate reduction | − | − | + | + |
| Hydrolysis of gelatin | − | − | − | + |
| Assimilation: | ||||
| Arabinose | − | − | w | + |
| Citric acid | − | − | + | − |
| Phenylacetic acid | − | + | − | − |
| Enzyme activity: | ||||
| Alkaline phosphatase | + | + | + | + |
| Acid phosphatase | w | + | + | + |
| Esterase(C4) | + | + | + | + |
| Esterase lipase (C8) | w | + | + | + |
| Lipid enzyme (C14) | − | − | − | − |
| Carbon source utilization: | ||||
| α-d-Glucose | − | w | w | − |
| D-Fucose | − | w | w | − |
| Fusidic Acid | − | w | w | w |
| Myo-Inositol | − | w | w | − |
| Minocycline | w | − | − | − |
| Tetrazolium Violet | + | w | w | w |
| α-Ketoglutaric Acid | − | w | w | w |
| Tween 40 | − | w | + | w |
| DNA G+C content (%) | 67.7 | 62.2 | 67.2 | 65.1 |
| Fatty Acid | 1 | 2 | 4 | 5 |
|---|---|---|---|---|
| C16:0 | 8.17 | 6.12 | 13.87 | 7.09 |
| C17:0 | 2.78 | - | 0.46 | 12.14 |
| C18:0 | 4.57 | 2.81 | 4.52 | 3.67 |
| C17:0 cyclo | - | 2.65 | - | 3.46 |
| C19:0 cyclo ω8c | 16.8 | 43.12 | 4.52 | 20.46 |
| C20:2 ω6,9c | 0.74 | 2.12 | - | 0.8 |
| 11-Methyl C18:1ω7c | 0.85 | 2.65 | - | 1.44 |
| C14:0 2-OH | 2.03 | - | - | - |
| C18:0 3-OH | 1.48 | 2.25 | 1.77 | 1.26 |
| * Summed feature 2 | 2.16 | 5.57 | 2.96 | 2.2 |
| * Summed feature 3 | 1.36 | 2.47 | 3.68 | 0.84 |
| * Summed feature 7 | 2.28 | 0.55 | - | - |
| * Summed feature 8 | 56.76 | 28.7 | 68.49 | 43.72 |
| Strain Name | Accession No. | Level | Contigs | N50 | L50 | Size (Mb) | GC% | No. of Genes | No. of Proteins | Isolation |
|---|---|---|---|---|---|---|---|---|---|---|
| Microvirga massiliensis JC119 | LN811386 | Scaffold | 365 | 56347 | 49 | 9.1 | 62.7 | 8575 | 8045 | Human stool |
| Microvirga vignae BR3299 | LCYG01000000 | Scaffold | 165 | 99306 | 19 | 6.47 | 61.1 | 6304 | 5674 | Nodule |
| Microvirga subterranea DSM 14364 | QQBB01000000 | Scaffold | 36 | 519753 | 4 | 5.15 | 65.1 | 4975 | 4902 | Thermal aquifer |
| Microvirga aerophila NBRC 106136 | BJYU00000000 | Contig | 628 | 43211 | 42 | 7.36 | 61.6 | 7220 | 7162 | Air |
| Microvirga guangxiensis CGMCC 1.7666 | FMVJ01000000 | Scaffold | 34 | 407384 | 5 | 4.72 | 61.4 | 4572 | 4499 | Paddy soil |
| Microvirga lotononidis WSM3557 | AJUA01000000 | Scaffold | 104 | 261220 | 10 | 7.08 | 63.1 | 6915 | 6524 | Nodule |
| Microvirga brassicacearum CDVBN77 | VCMV01000000 | Scaffold | 78 | 150733 | 10 | 5.22 | 62.3 | 4953 | 4755 | Rapeseed |
| Microvirga ossetica V5/3M | CP016616 | Complete | - | - | - | 5.84 | 63.5 | 5543 | 5265 | Nodule |
| Microvirga sp. 17 mud 1-3 | CP029481 | Complete | - | - | - | 4.4 | 64 | 4143 | 4001 | Soil |
| Microvirga sp. KLBC 81 | QDAH01000000 | Scaffold | 357 | 70985 | 26 | 6.86 | 61.3 | 6588 | 6085 | Nodule |
| Microvirga thermotolerans HR1 | CP045423 | Complete | - | - | - | 3.8 | 67.71 | 3875 | 3818 | Soil |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Gao, R.; Chen, Y.; Xue, D.; Han, J.; Wang, J.; Dai, Q.; Lin, M.; Ke, X.; Zhang, W. Isolation and Identification of Microvirga thermotolerans HR1, a Novel Thermo-Tolerant Bacterium, and Comparative Genomics among Microvirga Species. Microorganisms 2020, 8, 101. https://doi.org/10.3390/microorganisms8010101
Li J, Gao R, Chen Y, Xue D, Han J, Wang J, Dai Q, Lin M, Ke X, Zhang W. Isolation and Identification of Microvirga thermotolerans HR1, a Novel Thermo-Tolerant Bacterium, and Comparative Genomics among Microvirga Species. Microorganisms. 2020; 8(1):101. https://doi.org/10.3390/microorganisms8010101
Chicago/Turabian StyleLi, Jiang, Ruyu Gao, Yun Chen, Dong Xue, Jiahui Han, Jin Wang, Qilin Dai, Min Lin, Xiubin Ke, and Wei Zhang. 2020. "Isolation and Identification of Microvirga thermotolerans HR1, a Novel Thermo-Tolerant Bacterium, and Comparative Genomics among Microvirga Species" Microorganisms 8, no. 1: 101. https://doi.org/10.3390/microorganisms8010101