The Conifer Root and Stem Rot Pathogen (Heterobasidion parviporum): Effectome Analysis and Roles in Interspecific Fungal Interactions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Sampling and RNA Isolation in Paired Cultures
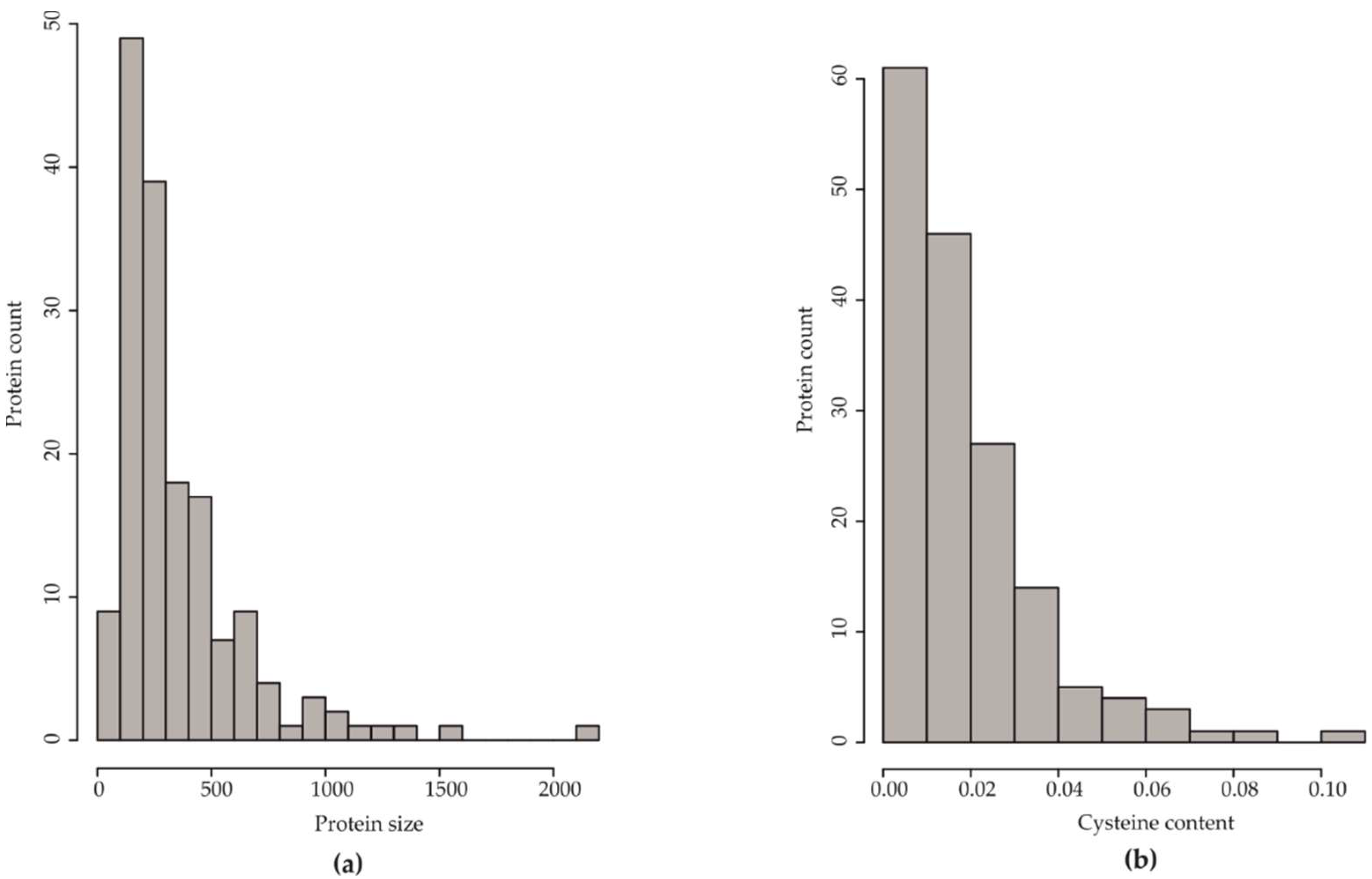
2.2. Effectome Analysis
2.2.1. Effectome Screening, Phylogenetic Analysis of Effectome, and HpSSP-Coding Genes Selection
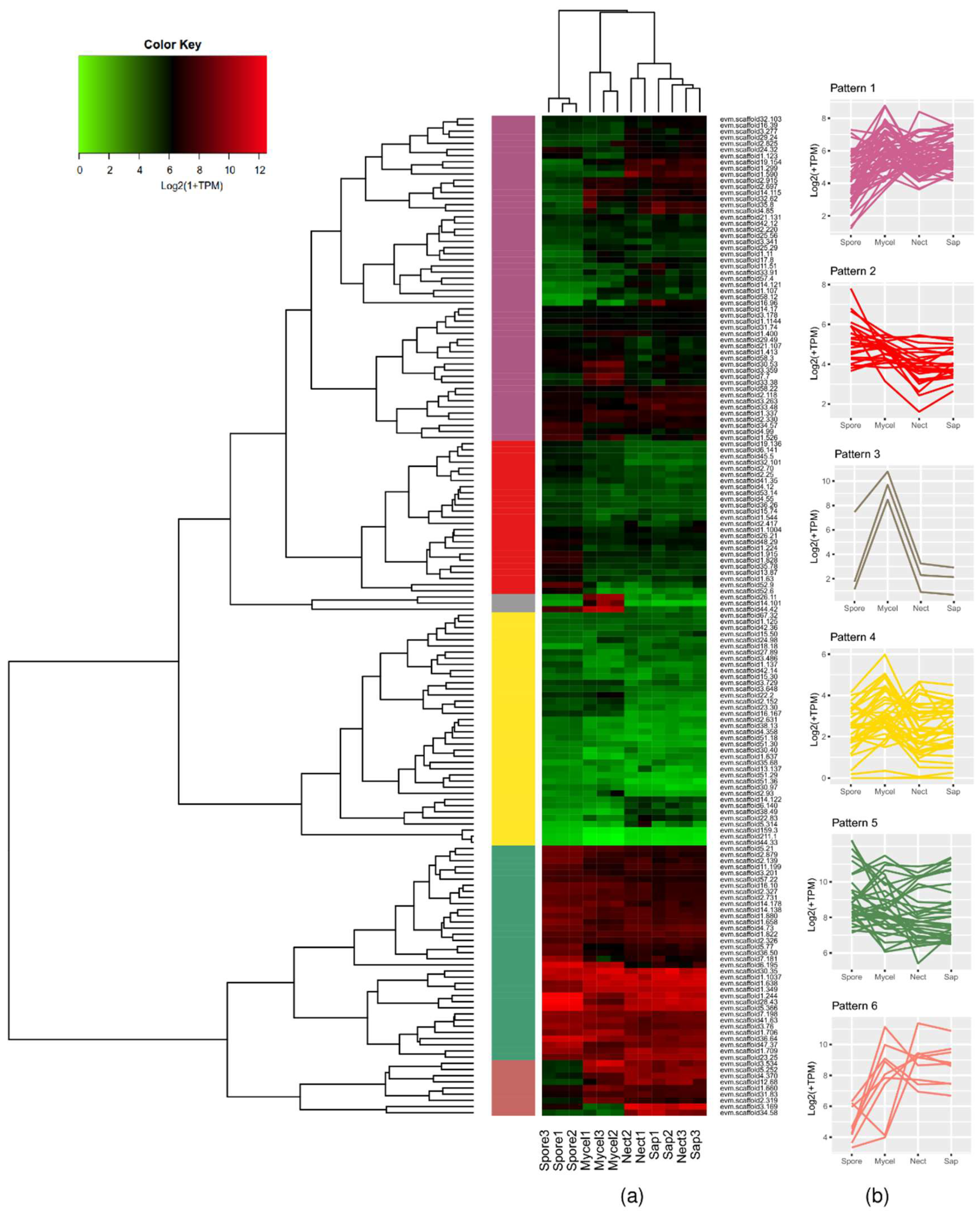
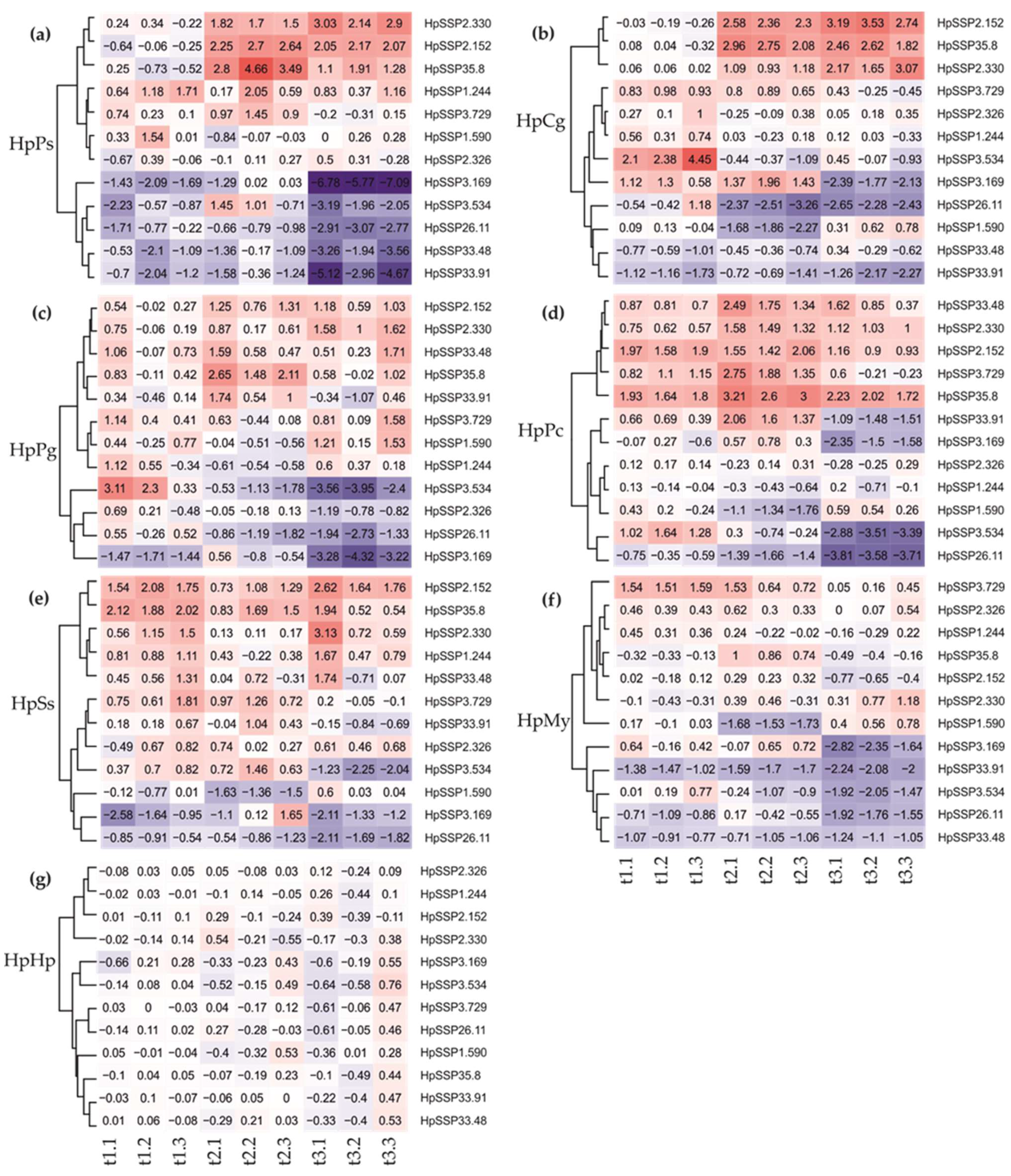
2.2.2. Effectome Expression Pattern at Diverse Developmental Stages
2.3. cDNA Synthesis
2.4. Quantitative Real-Time PCR
3. Results
3.1. Effectome Analysis
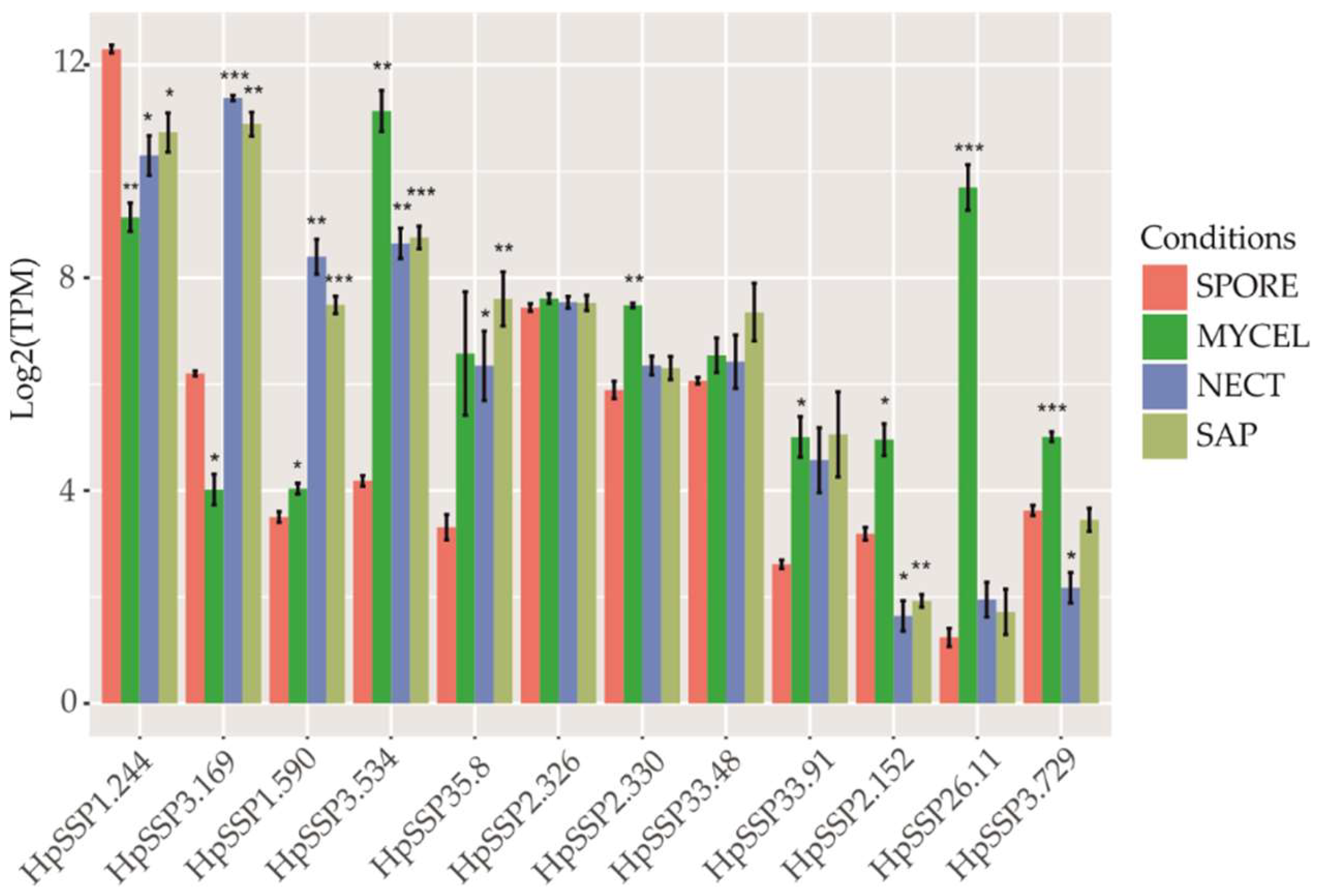
3.2. Transcriptomic Profiles of the Effector Candidates During Fungal Development and in Planta Expression
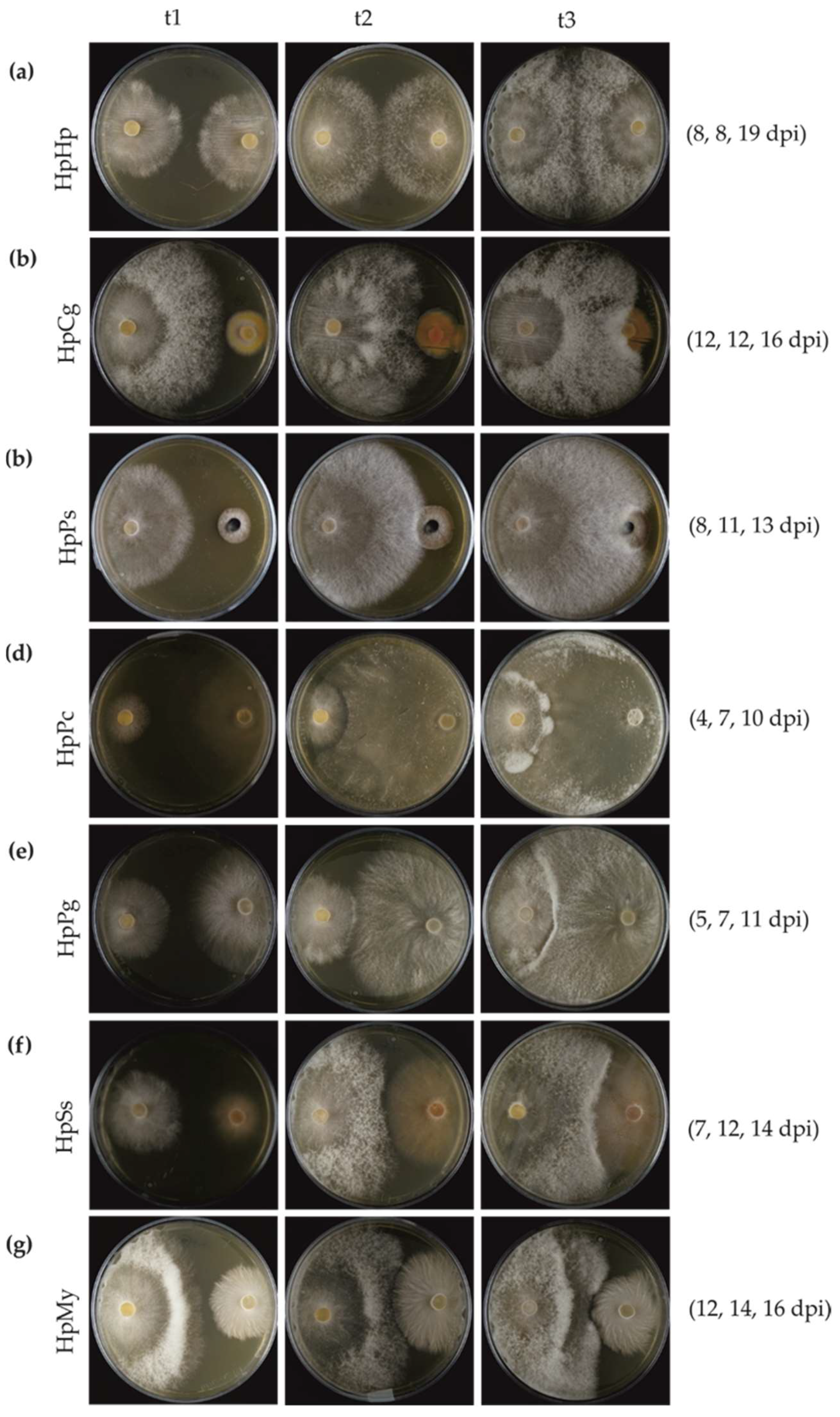
3.3. Mycelial Interaction in Dual Cultures
3.4. Gene Expression of HpSSPs on Dual Cultures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattila, U.; Nuutinen, T. Assessing the incidence of butt rot in Norway spruce in southern Finland. Silva Fennica 2007, 41, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Kärhä, K.; Koivusalo, V.; Palander, T.; Ronkanen, M. Treatment of Picea abies and Pinus sylvestris stumps with urea and Phlebiopsis gigantea for control of Heterobasidion. Forests 2018, 9, 139. [Google Scholar] [CrossRef] [Green Version]
- Asiegbu, F.O.; Adomas, A.; Stenlid, J.A.N. Conifer root and butt rot caused by Heterobasidion annosum (Fr.) Bref. s.l. Mol. Plant Pathol. 2005, 6, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Tubby, K.V.; Scott, D.; Webber, J.F. Relationship between stump treatment coverage using the biological control product PG Suspension, and control of Heterobasidion annosum on Corsican Pine, Pinus nigra ssp. laricio. For. Pathol. 2008, 38, 37–46. [Google Scholar] [CrossRef]
- Boddy, L. Interspecific combative interactions between wood-decaying Basidiomycetes. FEMS Microbiol. Ecol. 2000, 31, 185–194. [Google Scholar] [CrossRef]
- Hiscox, J.; O’Leary, J.; Boddy, L. Fungus wars: Basidiomycete battles in wood decay. Stud. Mycol. 2018, 89, 117–124. [Google Scholar] [CrossRef]
- Hiscox, J.; Baldrian, P.; Rogers, H.J.; Boddy, L. Changes in oxidative enzyme activity during interspecific mycelial interactions involving the white-rot fungus Trametes versicolor. Fungal Genet. Biol. 2010, 47, 562–571. [Google Scholar] [CrossRef]
- Baldrian, P. Increase of laccase activity during interspecific interactions of white-rot fungi. FEMS Microbiol. Ecol. 2004, 50, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Savoie, J.-M.; Mata, G.; Billette, C. Extracellular laccase production during hyphal interactions between Trichoderma sp. and Shiitake, Lentinula edodes. Appl. Microbiol. Biotechnol. 1998, 49, 589–593. [Google Scholar] [CrossRef]
- Evans, J.A.; Eyre, C.A.; Rogers, H.J.; Boddy, L.; Müller, C.T. Changes in volatile production during interspecific interactions between four wood rotting fungi growing in artificial media. Fungal Ecol. 2008, 1, 57–68. [Google Scholar] [CrossRef]
- El Ariebi, N.; Hiscox, J.; Scriven, S.A.; Müller, C.T.; Boddy, L. Production and effects of volatile organic compounds during interspecific interactions. Fungal Ecol. 2016, 20, 144–154. [Google Scholar] [CrossRef]
- Eyre, C.; Muftah, W.; Hiscox, J.; Hunt, J.; Kille, P.; Boddy, L.; Rogers, H.J. Microarray analysis of differential gene expression elicited in Trametes versicolor during interspecific mycelial interactions. Fungal Biol. 2010, 114, 646–660. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Sun, H.; Vainio, E.J.; Raffaello, T.; Kovalchuk, A.; Morin, E.; Duplessis, S.; Asiegbu, F.O. Intraspecific comparative genomics of isolates of the Norway spruce pathogen (Heterobasidion parviporum) and identification of its potential virulence factors. BMC Genom. 2018, 19, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting fungal effector proteins from secretomes using machine learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jonge, R. In Silico identification and characterization of effector catalogs. In Plant Fungal Pathogens: Methods and Protocols; Methods in Molecular Biology; Bolton, M.D., Thomma, B.P.H.J., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 415–425. [Google Scholar] [CrossRef]
- Hacquard, S.; Joly, D.L.; Lin, Y.C.; Tisserant, E.; Feau, N.; Delaruelle, C.; Legue, V.; Kohler, A.; Tanguay, P.; Petre, B.; et al. A comprehensive analysis of genes encoding small secreted proteins identifies candidate effectors in Melampsora larici-populina (Poplar Leaf Rust). Mol. Plant. Microbe Interact. 2012, 25, 279–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Manners, J.M.; Singh, K.B.; Taylor, J.M. Advances and challenges in computational prediction of effectors from plant pathogenic fungi. PLoS Pathog. 2015, 11, e1004806. [Google Scholar] [CrossRef] [Green Version]
- Terhonen, E.; Keriö, S.; Sun, H.; Asiegbu, F.O. Endophytic fungi of Norway spruce roots in boreal pristine mire, drained peatland and mineral soil and their inhibitory effect on Heterobasidion parviporum in vitro. Fungal Ecol. 2014, 9, 17–26. [Google Scholar] [CrossRef]
- TRI Reagent Protocol. Available online: https://www.sigmaaldrich.com/technical-documents/protocols/biology/tri-reagent.html (accessed on 27 June 2019).
- Zeng, Z.; Wu, J.; Kovalchuk, A.; Raffaello, T.; Wen, Z.; Liu, M.; Asiegbu, F.O. Genome-wide DNA methylation and transcriptomic profiles in the lifestyle strategies and asexual development of the forest fungal pathogen Heterobasidion parviporum. Epigenetics 2019, 14, 16–40. [Google Scholar] [CrossRef] [Green Version]
- Morón-Ríos, A.; Gómez-Cornelio, S.; Ortega-Morales, B.O.; De la Rosa-García, S.; Partida-Martínez, L.P.; Quintana, P.; Alayón-Gamboa, J.A.; Cappello-García, S.; González-Gómez, S. Interactions between abundant fungal species influence the fungal community assemblage on limestone. PLoS ONE 2017, 12, e0188443. [Google Scholar] [CrossRef] [Green Version]
- Terhonen, E.; Sipari, N.; Asiegbu, F.O. Inhibition of phytopathogens by fungal root endophytes of Norway spruce. Biol. Control 2016, 99, 53–63. [Google Scholar] [CrossRef]
- Adomas, A.; Eklund, M.; Johansson, M.; Asiegbu, F.O. Identification and analysis of differentially expressed cDNAs during nonself-competitive interaction between Phlebiopsis gigantea and Heterobasidion parviporum. FEMS Microbiol. Ecol. 2006, 57, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boddy, L. Interactions between fungi and other microbes. In The Fungi; Elsevier: Amsterdam, The Netherlands, 2016; pp. 337–360. [Google Scholar] [CrossRef]
- McCotter, S.W.; Horianopoulos, L.C.; Kronstad, J.W. Regulation of the fungal secretome. Curr. Genet. 2016, 62, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Pellegrin, C.; Morin, E.; Martin, F.M.; Veneault-Fourrey, C. Comparative analysis of secretomes from ectomycorrhizal fungi with an emphasis on small-secreted proteins. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Feldman, D.; Amedi, N.; Carmeli, S.; Yarden, O.; Hadar, Y. Manipulating the expression of small secreted protein 1 (ssp1) alters patterns of development and metabolism in the white-rot fungus Pleurotus ostreatus. Appl. Environ. Microbiol. 2019, 85, e00761-19. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-T.; Jeon, J.; Choi, J.; Cheong, K.; Song, H.; Choi, G.; Kang, S.; Lee, Y.-H. Kingdom-wide analysis of fungal small secreted proteins (SSPs) reveals their potential role in host association. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Porto, B.N.; Caixeta, E.T.; Mathioni, S.M.; Vidigal, P.M.P.; Zambolim, L.; Zambolim, E.M.; Donofrio, N.; Polson, S.W.; Maia, T.A.; Chen, C.; et al. Genome sequencing and transcript analysis of Hemileia vastatrix reveal expression dynamics of candidate effectors dependent on host compatibility. PLoS ONE 2019, 14, e0215598. [Google Scholar] [CrossRef]
- Kettles, G.J.; Bayon, C.; Sparks, C.A.; Canning, G.; Kanyuka, K.; Rudd, J.J. Characterization of an antimicrobial and phytotoxic ribonuclease secreted by the fungal wheat pathogen Zymoseptoria tritici. New Phytol. 2018, 217, 320–331. [Google Scholar] [CrossRef] [Green Version]
- Raffaello, T.; Asiegbu, F.O. Small secreted proteins from the necrotrophic conifer pathogen Heterobasidion annosum s.l. (HaSSPs) induce cell death in Nicotiana benthamiana. Sci. Rep. 2017, 7, 8000. [Google Scholar] [CrossRef]
- Wen, Z.; Raffaello, T.; Zeng, Z.; Pavicic, M.; Asiegbu, F.O. Chlorophyll fluorescence imaging for monitoring effects of Heterobasidion parviporum small secreted protein induced cell death and in planta defense gene expression. Fungal Genet. Biol. 2019, 126, 37–49. [Google Scholar] [CrossRef]
- Mukherjee, S.; Khowala, S. Unraveling the secretome of Termitomyces clypeatus grown on agroresidues as a potential source for bioethanol production. Process Biochem. 2016, 51, 1793–1807. [Google Scholar] [CrossRef]
- Martinez, D.; Challacombe, J.; Morgenstern, I.; Hibbett, D.; Schmoll, M.; Kubicek, C.P.; Ferreira, P.; Ruiz-Duenas, F.J.; Martinez, A.T.; Kersten, P.; et al. Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proc. Natl. Acad. Sci. USA 2009, 106, 1954–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mgbeahuruike, A.C.; Kohler, A.; Asiegbu, F.O. Expression analysis of the impact of culture filtrates from the biocontrol agent, Phlebiopsis gigantea on the conifer pathogen, Heterobasidion annosum s.s. transcriptome. Microb. Ecol. 2013, 66, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Mgbeahuruike, A.C.; Kovalchuk, A.; Chen, H.; Ubhayasekera, W.; Asiegbu, F.O. Evolutionary analysis of hydrophobin gene family in two wood-degrading Basidiomycetes, Phlebia brevispora and Heterobasidion annosum s.l. BMC Evol. Biol. 2013, 13, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nygren, K.; Dubey, M.; Zapparata, A.; Iqbal, M.; Tzelepis, G.D.; Durling, M.B.; Jensen, D.F.; Karlsson, M. The mycoparasitic fungus Clonostachys rosea responds with both common and specific gene expression during interspecific interactions with fungal prey. Evol. Appl. 2018, 11, 931–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveau, A.; Palin, B.; Delaruelle, C.; Peter, M.; Kohler, A.; Pierrat, J.C.; Sarniguet, A.; Garbaye, J.; Martin, F.; Frey-Klett, P. The mycorrhiza Helper Pseudomonas fluorescens BBc6R8 has a specific priming effect on the growth, morphology and gene expression of the ectomycorrhizal fungus Laccaria bicolor S238N. New Phytol. 2007, 175, 743–755. [Google Scholar] [CrossRef]
- Pratt, J.E.; Niemi, M.; Sierota, Z.H. Comparison of three products based on Phlebiopsis gigantea for the control of Heterobasidion annosum in Europe. Biocontrol Sci. Technol. 2000, 10, 467–477. [Google Scholar] [CrossRef]
- Ikediugwu, F.E.O. The interface in hyphal interference by Peniophora gigantea against Heterobasidion annosum. Trans. Br. Mycol. Soc. 1976, 66, 291–296. [Google Scholar] [CrossRef]
- Raffaello, T.; Asiegbu, F.O. Evaluation of potential reference genes for use in gene expression studies in the conifer pathogen (Heterobasidion annosum). Mol. Biol. Rep. 2013, 40, 4605–4611. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HpSSP | Homolog to H. annosum.s.s a | Sequence Length | Cysteins Number | Prediction in EffetorP | Transcriptomic Pattern |
|---|---|---|---|---|---|
| HpSSP1.244 | Hetan1.estExt_Genewise1Plus.C_10476 | 194 | 10 | Effector | Pattern 5 |
| HpSSP2.152 | Hetan1.Genemark.3574_g | 152 | 4 | Effector | Pattern 4 |
| HpSSP2.326 | Hetan1.estExt_Genewise1Plus.C_40816 | 242 | 12 | Effector | Pattern 5 |
| HpSSP2.330 | fgenesh1_pm.03_#_457 | 192 | 2 | Effector | Pattern 1 |
| HpSSP26.11 | e_gw1.11.833.1 | 254 | 12 | Noneffector | Pattern 3 |
| HpSSP3.534 | Hetan1.Genemark.10137_g | 185 | 10 | Effector | Pattern 6 |
| HpSSP3.729 | Hetan1.gw1.14.303.1 | 216 | 4 | Effector | Pattern 4 |
| HpSSP33.48 | Hetan1.fgenesh2_pg.C_scaffold_13000272 | 243 | 4 | Effector | Pattern 1 |
| HpSSP33.91 | Hetan1.fgenesh2_pg.C_scaffold_13000272 | 245 | 4 | Effector | Pattern 1 |
| HpSSP35.8 | estExt_fgenesh1_pg.C_130285 | 177 | 2 | Unlikely effector | Pattern 1 |
| HpSSP1.590 | Genemark.573_g | 321 | 0 | Noneffector | Pattern 1 |
| HpSSP3.169 | gw1.07.1673.1 | 524 | 0 | Noneffector | Pattern 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Z.; Zeng, Z.; Ren, F.; Asiegbu, F.O. The Conifer Root and Stem Rot Pathogen (Heterobasidion parviporum): Effectome Analysis and Roles in Interspecific Fungal Interactions. Microorganisms 2019, 7, 658. https://doi.org/10.3390/microorganisms7120658
Wen Z, Zeng Z, Ren F, Asiegbu FO. The Conifer Root and Stem Rot Pathogen (Heterobasidion parviporum): Effectome Analysis and Roles in Interspecific Fungal Interactions. Microorganisms. 2019; 7(12):658. https://doi.org/10.3390/microorganisms7120658
Chicago/Turabian StyleWen, Zilan, Zhen Zeng, Fei Ren, and Fred O. Asiegbu. 2019. "The Conifer Root and Stem Rot Pathogen (Heterobasidion parviporum): Effectome Analysis and Roles in Interspecific Fungal Interactions" Microorganisms 7, no. 12: 658. https://doi.org/10.3390/microorganisms7120658