Microbial and Phenyl Acid Dynamics during the Start-up Phase of Anaerobic Straw Degradation in Meso- and Thermophilic Batch Reactors


Abstract
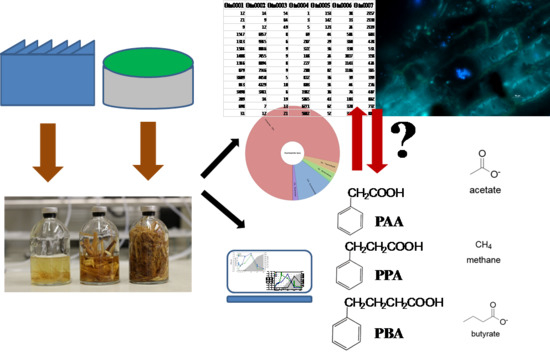
:
1. Introduction
2. Materials and Methods
3. Results
3.1. Mesophilic Communities
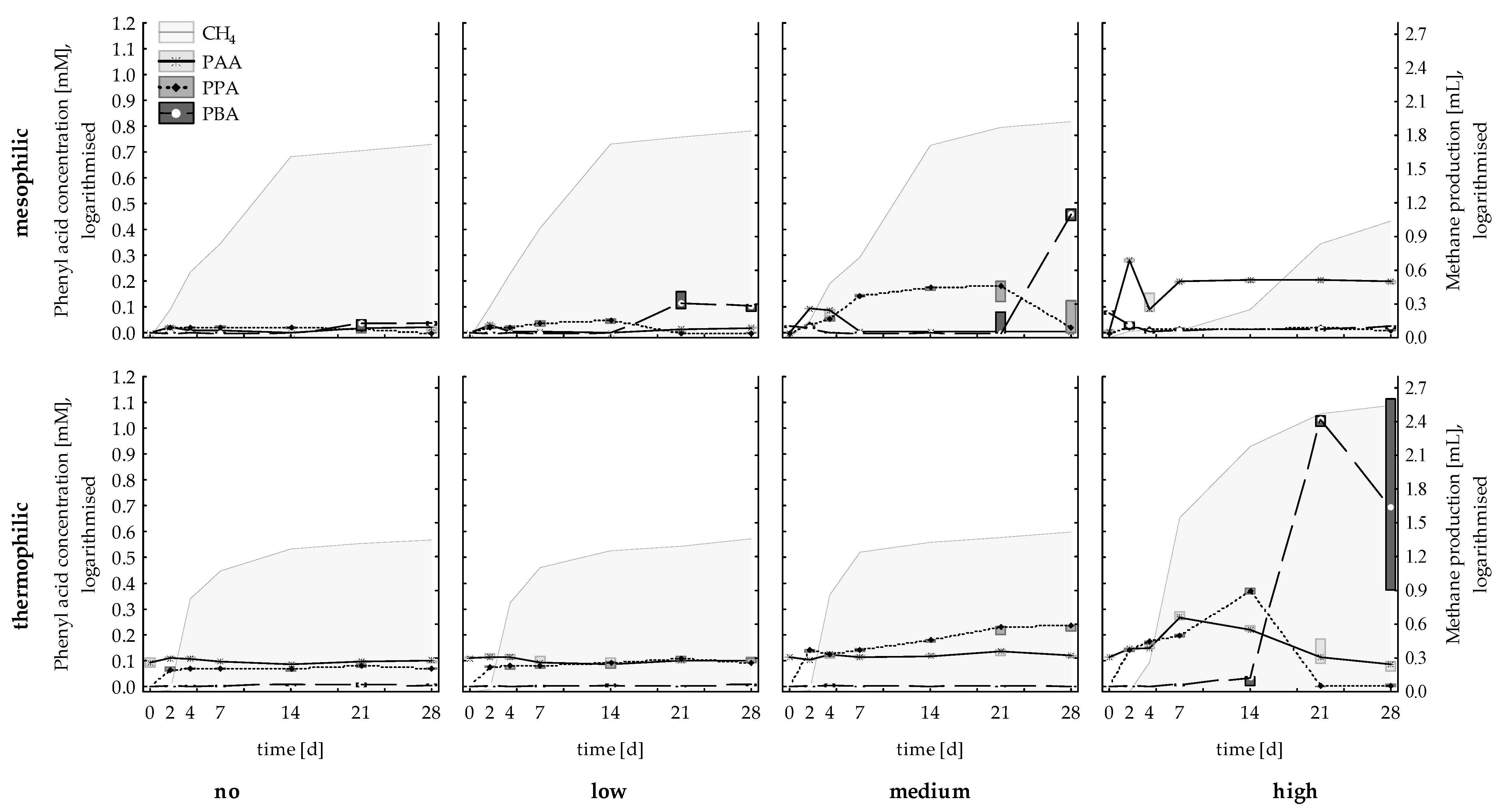
3.1.1. Methane Production, Acetate, and Phenyl Acid Concentrations
3.1.2. Microbial Community Composition
3.2. Thermophilic Communities
3.2.1. Methane Production, Acetate, and Phenyl Acid Concentrations
3.2.2. Microbial Community Composition
3.3. Prediction of Metagenomic Properties—Piphillin Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
Appendix C
References
- Panwar, N.L.; Kaushik, S.C.; Kothari, S. Role of renewable energy sources in environmental protection: A review. Renew. Sustain. Energy Rev. 2011, 15, 1513–1524. [Google Scholar] [CrossRef]
- Zieminski, K.; Romanowska, I.; Kowalska, M. Enzymatic pretreatment of lignocellulosic wastes to improve biogas production. Waste Manag. 2012, 32, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Westerholm, M.; Moestedt, J.; Schnürer, A. Biogas production through syntrophic acetate oxidation and deliberate operating strategies for improved digester performance. Appl. Energy 2016, 179, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.T.; Qiu, Y.L.; Yuan, X.Z.; Shi, X.S.; Xu, X.H.; Guo, R.B. Molecular characterization of bacterial and archaeal communities in a full-scale anaerobic reactor treating corn straw. Bioresour. Technol. 2013, 143, 512–518. [Google Scholar] [CrossRef]
- Rétfalvi, T.; Tukacs-Hájos, A.; Szabó, P. Effects of artificial overdosing of p-cresol and phenylacetic acid on the anaerobic fermentation of sugar beet pulp. Int. Biodeterior. Biodegrad. 2013, 83, 112–118. [Google Scholar] [CrossRef]
- Wagner, A.O.; Lackner, N.; Mutschlechner, M.; Prem, E.M.; Markt, R.; Illmer, P. Biological Pretreatment Strategies for Second-Generation Lignocellulosic Resources to Enhance Biogas Production. Energies 2018, 11, 1797. [Google Scholar] [CrossRef] [Green Version]
- Braguglia, C.M.; Gallipoli, A.; Gianico, A.; Pagliaccia, P. Anaerobic bioconversion of food waste into energy: A critical review. Bioresour. Technol. 2018, 248, 37–56. [Google Scholar] [CrossRef]
- Griehl, C.; Hecht, C.; Streuber, S.; Ganß, Y.; Kremp, H. Bestimmung von Phenylessigsäure zur Früherkennung von Prozessstörungen in Biogasanlagen. Chem. Ing. Tech. 2010, 82, 2223–2229. [Google Scholar] [CrossRef]
- Hecht, C.; Griehl, C. Investigation of the accumulation of aromatic compounds during biogas production from kitchen waste. Bioresour. Technol. 2009, 100, 654–658. [Google Scholar] [CrossRef]
- Moset, V.; Poulsen, M.; Wahid, R.; Højberg, O.; Møller, H.B. Mesophilic versus thermophilic anaerobic digestion of cattle manure: Methane productivity and microbial ecology. Microb. Biotechnol. 2015, 8, 787–800. [Google Scholar] [CrossRef]
- Gebreeyessus, G.D.; Jenicek, P. Thermophilic versus Mesophilic Anaerobic Digestion of Sewage Sludge: A Comparative Review. Bioengineering 2016, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Glissmann, K.; Hammer, E.; Conrad, R. Production of aromatic compounds during methanogenic degradation of straw in rice field soil. FEMS Microbiol. Ecol. 2005, 52, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, E.L.; Mueller, R.E.; Sievers, D.M.; Georgacakis, D.G.; Gerhardt, K.O. Phenylacetic acid in an anaerobic swine manure digester. J. Ind. Microbiol. 1986, 1, 57–61. [Google Scholar] [CrossRef]
- Cabrol, L.; Urra, J.; Rosenkranz, F.; Kroff, P.A.; Plugge, C.M.; Lesty, Y.; Chamy, R. Influence of phenylacetic acid pulses on anaerobic digestion performance and archaeal community structure in WWTP sewage sludge digesters. Water Sci. Technol. 2015, 71, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Mimura, N.; Shirai, M.; Sato, O. Bond cleavage of lignin model compounds into aromatic monomers using supported metal catalysts in supercritical water. Sci. Rep. 2017, 7, 46172. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds—From one strategy to four. Nat. Rev. Microbiol. 2011, 9, 803–816. [Google Scholar] [CrossRef]
- Boll, M.; Löffler, C.; Morris, B.E.L.; Kung, J.W. Anaerobic degradation of homocyclic aromatic compounds via arylcarboxyl-coenzyme A esters: Organisms, strategies and key enzymes. Environ. Microbiol. 2014, 16, 612–627. [Google Scholar] [CrossRef]
- Kuntze, K.; Shinoda, Y.; Moutakki, H.; McInerney, M.J.; Vogt, C.; Richnow, H.H.; Boll, M. 6-Oxocyclohex-1-ene-1-carbonyl-coenzyme A hydrolases from obligately anaerobic bacteria: Characterization and identification of its gene as a functional marker for aromatic compounds degrading anaerobes. Environ. Microbiol. 2008, 10, 1547–1556. [Google Scholar] [CrossRef]
- Schink, B.; Philipp, B.; Müller, J. Anaerobic Degradation of Phenolic Compounds. Naturwissenschaften 2000, 87, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Carmona, M.; Zamarro, M.T.; Blázquez, B.; Durante-Rodríguez, G.; Juárez, J.F.; Valderrama, J.A.; Barragán, M.J.L.; García, J.L.; Díaz, E. Anaerobic catabolism of aromatic compounds: A genetic and genomic view. Microbiol. Mol. Biol. Rev. 2009, 73, 71–133. [Google Scholar] [CrossRef] [Green Version]
- Harwood, C.S.; Burchhardt, G.; Herrmann, H.; Fuchs, G. Anaerobic metabolism of aromatic compounds via the benzoyl-CoA pathway. FEMS Microbiol. Rev. 1998, 22, 439–458. [Google Scholar] [CrossRef]
- Wagner, A.O.; Prem, E.M.; Markt, R.; Kaufmann, R.; Illmer, P. Formation of phenylacetic acid and phenylpropionic acid under different overload conditions during mesophilic and thermophilic anaerobic digestion. Biotechnol. Biofuels 2019, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Chino, K.; Kamimura, N.; Masai, E.; Yumoto, I.; Kamagata, Y. Methanogenic degradation of lignin-derived monoaromatic compounds by microbial enrichments from rice paddy field soil. Sci. Rep. 2015, 5, 14295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, B.E.; Bhupathiraju, V.K.; Tanner, R.S.; Woese, C.R.; McInerney, M.J. Syntrophus aciditrophicus sp. nov., a new anaerobic bacterium that degrades fatty acids and benzoate in syntrophic association with hydrogen-using microorganisms. Arch. Microbiol. 1999, 171, 107–114. [Google Scholar] [CrossRef]
- Walter, A.; Probst, M.; Franke-Whittle, I.H.; Ebner, C.; Podmirseg, S.M.; Etemadi-Shalamzari, M.; Hupfauf, S.; Insam, H. Microbiota in anaerobic digestion of sewage sludge with and without co-substrates. Water Environ. J. 2019, 33, 214–222. [Google Scholar] [CrossRef]
- Illmer, P.; Schwarzenauer, T.; Malin, C.; Wagner, A.O.; Miller, L.M.; Gstraunthaler, G. Process parameters within a 750,000 litre anaerobic digester during a year of disturbed fermenter performance. Waste Manag. 2009, 29, 1838–1843. [Google Scholar] [CrossRef]
- Wagner, A.O.; Markt, R.; Mutschlechner, M.; Lackner, N.; Prem, E.M.; Praeg, N.; Illmer, P. Medium Preparation for the Cultivation of Microorganisms under Strictly Anaerobic/Anoxic Conditions. JoVE 2019. [Google Scholar] [CrossRef]
- Wagner, A.O.; Hohlbrugger, P.; Lins, P.; Illmer, P. Effects of different nitrogen sources on the biogas production—A lab-scale investigation. Microbiol. Res. 2012, 167, 630–636. [Google Scholar] [CrossRef]
- Lins, P.; Malin, C.; Wagner, A.O.; Illmer, P. Reduction of accumulated volatile fatty acids by an acetate-degrading enrichment culture. FEMS Microbiol. Ecol. 2010, 71, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.O.; Lins, P.; Malin, C.; Reitschuler, C.; Illmer, P. Impact of protein-, lipid- and cellulose-containing complex substrates on biogas production and microbial communities in batch experiments. Sci. Total Environ. 2013, 458–460, 256–266. [Google Scholar] [CrossRef]
- Reitschuler, C.; Lins, P.; Wagner, A.O.; Illmer, P. Cultivation of moonmilk-born non-extremophilic Thaum and Euryarchaeota in mixed culture. Anaerobe 2014, 29, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.O.; Markt, R.; Puempel, T.; Illmer, P.; Insam, H.; Ebner, C. Sample preparation, preservation, and storage for volatile fatty acid quantification in biogas plants. Eng. Life Sci. 2017, 17, 132–139. [Google Scholar] [CrossRef]
- Steinberg, L.M.; Regan, J.M. Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl. Environ. Microbiol. 2008, 74, 6663–6671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, L.M.; Regan, J.M. mcrA-targeted real-time quantitative PCR method to examine methanogen communities. Appl. Environ. Microbiol. 2009, 75, 4435–4442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Jansson, J.K.; Knight, R. The Earth Microbiome project: Successes and aspirations. BMC Biol. 2014, 12, 69. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.O.; Praeg, N.; Reitschuler, C.; Illmer, P. Effect of DNA extraction procedure, repeated extraction and ethidium monoazide (EMA)/propidium monoazide (PMA) treatment on overall DNA yield and impact on microbial fingerprints for bacteria, fungi and archaea in a reference soil. Appl. Soil Ecol. 2015, 93, 56–64. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [Green Version]
- Iwai, S.; Weinmaier, T.; Schmidt, B.L.; Albertson, D.G.; Poloso, N.J.; Dabbagh, K.; DeSantis, T.Z. Piphillin: Improved Prediction of Metagenomic Content by Direct Inference from Human Microbiomes. PLoS ONE 2016, 11, e0166104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Sekiguchi, Y.; Kamagata, Y.; Syutsubo, K.; Ohashi, A.; Harada, H.; Nakamura, K. Phylogenetic diversity of mesophilic and thermophilic granular sludges determined by 16S rRNA gene analysis. Microbiology (Reading, Engl.) 1998, 144 Pt 9, 2655–2665. [Google Scholar] [CrossRef] [Green Version]
- De Vrieze, J.; Christiaens, M.E.R.; Walraedt, D.; Devooght, A.; Ijaz, U.Z.; Boon, N. Microbial community redundancy in anaerobic digestion drives process recovery after salinity exposure. Water Res. 2017, 111, 109–117. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the Genetic Basis of Fibrolytic Specialization by Lachnospiraceae and Ruminococcaceae in Diverse Gut Communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Cornick, N.A.; Jensen, N.S.; Stahl, D.A.; Hartman, P.A.; Allison, M.J. Lachnospira pectinoschiza sp. nov., an anaerobic pectinophile from the pig intestine. Int. J. Syst. Bacteriol. 1994, 44, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsyurbenko, O.R.; Friedrich, M.W.; Simankova, M.V.; Nozhevnikova, A.N.; Golyshin, P.N.; Timmis, K.N.; Conrad, R. Shift from acetoclastic to H2-dependent methanogenesis in a west Siberian peat bog at low pH values and isolation of an acidophilic Methanobacterium strain. Appl. Environ. Microbiol. 2007, 73, 2344–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vrieze, J.; Hennebel, T.; Boon, N.; Verstraete, W. Methanosarcina: The rediscovered methanogen for heavy duty biomethanation. Bioresour. Technol. 2012, 112, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Illmer, P.; Reitschuler, C.; Wagner, A.O.; Schwarzenauer, T.; Lins, P. Microbial succession during thermophilic digestion: The potential of Methanosarcina sp. PLoS ONE 2014, 9, e86967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahnke, S.; Langer, T.; Koeck, D.E.; Klocke, M. Description of Proteiniphilum saccharofermentans sp. nov., Petrimonas mucosa sp. nov. and Fermentimonas caenicola gen. nov., sp. nov., isolated from mesophilic laboratory-scale biogas reactors, and emended description of the genus Proteiniphilum. Int. J. Syst. Evol. Microbiol. 2016, 66, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Dong, X. Acetanaerobacterium elongatum gen. nov., sp. nov., from paper mill waste water. Int. J. Syst. Evol. Microbiol. 2004, 54, 2257–2262. [Google Scholar] [CrossRef] [Green Version]
- Defnoun, S.; Labat, M.; Ambrosio, M.; Garcia, J.L.; Patel, B.K. Papillibacter cinnamivorans gen. nov., sp. nov., a cinnamate-transforming bacterium from a shea cake digester. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 3, 1221–1228. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qiao, J.T.; Yuan, X.Z.; Guo, R.B.; Qiu, Y.L. Hydrogenispora ethanolica gen. nov., sp. nov., an anaerobic carbohydrate-fermenting bacterium from anaerobic sludge. Int. J. Syst. Evol. Microbiol. 2014, 64, 1756–1762. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Ma, C.; Gao, C.; Li, F.; Qin, J.; Zhang, H.; Wang, K.; Xu, P. Efficient conversion of phenylpyruvic acid to phenyllactic acid by using whole cells of Bacillus coagulans SDM. PLoS ONE 2011, 6, e19030. [Google Scholar] [CrossRef] [Green Version]
- Atashgahi, S.; Hornung, B.; van der Waals, M.J.; da Rocha, U.N.; Hugenholtz, F.; Nijsse, B.; Molenaar, D.; van Spanning, R.; Stams, A.J.M.; Gerritse, J.; et al. A benzene-degrading nitrate-reducing microbial consortium displays aerobic and anaerobic benzene degradation pathways. Sci. Rep. 2018, 8, 4490. [Google Scholar] [CrossRef]
- Meng, L.W.; Li, X.K.; Wang, S.T.; Liu, L.L.; Ma, K.L.; Zhang, J. The long-term impact of cefalexin on organic substrate degradation and microbial community structure in EGSB system. Chemosphere 2017, 184, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Westram, R.; Bader, K.; Prüsse, E.; Kumar, Y.; Meier, H.; Glöckner, F.O.; Ludwig, W. ARB: A software environment for sequence data. In Handbook of Molecular Microbial Ecology I: Metagenomics and Complementary Approaches; De Bruijn, F.J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 399–406. ISBN 9780470644799. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C Load | Mesophilic Core Microbiome | Thermophilic Core Microbiome | |
|---|---|---|---|
| control | Methanosarcina Macellibacteroides Anaerolineaceae (uncultured genus) * Paludibacteraceae (uncultured genus) | Caldicoprobacter Hydrogenispora Ruminiclostridium * Clostridia DTU014 genus Methanoculleus * Proteiniphilum * Clostridiales vadinBB60_group genus | Tepidimicrobium * Tepidanaerobacter Clostridium sensu stricto 1 Defluviitoga Christensenellaceae (uncultured genus) Firmicutes (uncultured genus) Clostridia MBA03 genus |
| LCL | Methanosarcina Macellibacteroides Clostridia DTU014 genus Anaerolineaceae (uncultured genus) * Proteiniphilum Lachnospiraceae (uncultured genus) Paludibacteraceae (uncultured genus) Ruminiclostridium 1 Bacteroides Christensenellaceae R-7_group Petrimonas | Caldicoprobacter Hydrogenispora Ruminiclostridium * Clostridia DTU014 genus Methanoculleus * Proteiniphilum * Clostridiales vadinBB60_group genus | Herbinix Tepidimicrobium * Tepidanaerobacter Defluviitoga Christensenellaceae (uncultured genus) Firmicutes (uncultured genus) Clostridia MBA03 genus |
| MCL | Methanosarcina Macellibacteroides Caproiciproducens Anaerolineaceae (uncultured genus) * Proteiniphilum Bacteroides Petrimonas Acetanaerobacterium | Methanosarcina Caldicoprobacter Caproiciproducens Hydrogenispora Ruminiclostridium * Clostridia DTU014 genus Methanoculleus * Proteiniphilum * Clostridiales vadinBB60_group genus Lachnospiraceae (uncultured genus) Herbinix Ruminiclostridium 1 | Tepidimicrobium * Tepidanaerobacter Clostridium sensu stricto 1 Defluviitoga Christensenellaceae (uncultured genus) Firmicutes (uncultured genus) Ruminococcaceae UCG-010 Izimaplasmatales genus Clostridia MBA03 genus Oxobacter Anaerocolumna Limnochordales (uncultured genus) |
| HCL | Caproiciproducens Clostridia DTU014 genus Anaerolineaceae (uncultured genus) * Clostridium sensu stricto 1 Clostridia D8A-2 genus Ruminococcaceae genus Lachnospira Candidatus Caldatribacterium Methanobacterium Clostridium sensu stricto 12 | Methanosarcina Caproiciproducens Ruminiclostridium * | Methanoculleus * Proteiniphilum * Tepidimicrobium * |
| Temperature | C-load (Class) | Sample Size | LEfSe Biomarkers (LDA Score ≥ 4.0) |
|---|---|---|---|
| mesophilic | control | 9 | Anaerolineaceae (uncultured genus) Clostridia DTU014 genus Bacteroidetes vadinHA17 genus |
| LCL | 6 | Macellibacteroides Paludibacteraceae (uncultured genus) Ruminiclostridium 1 Christensenellaceae R-7 group Lachnospiraceae (uncultured genus) Proteiniphilum Ruminiclostridium Dysgonomonadaceae (uncultured genus) Mobilitalea | |
| MCL | 6 | Bacteroides Petrimonas | |
| HCL | 6 | Caproiciproducens Ruminococcaceae genus Lachnospira Clostridium sensu stricto 1 Methanobacterium Clostridia D8A-2 genus | |
| thermophilic | control | 9 | Defluviitoga Syntrophaceticus Clostridia MBA03 genus Lactobacillus |
| LCL | 6 | Caldicoprobacter Methanoculleus | |
| MCL | 6 | Ruminiclostridium Lachnospiraceae (uncultured genus) Herbinix Izimaplasmatales genus Anaerocolumna | |
| HCL | 6 | Hydrogenispora Methanosarcina Ruminococcaceae UCG-010 Caproiciproducens Proteiniphilum |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prem, E.M.; Markt, R.; Lackner, N.; Illmer, P.; Wagner, A.O. Microbial and Phenyl Acid Dynamics during the Start-up Phase of Anaerobic Straw Degradation in Meso- and Thermophilic Batch Reactors. Microorganisms 2019, 7, 657. https://doi.org/10.3390/microorganisms7120657
Prem EM, Markt R, Lackner N, Illmer P, Wagner AO. Microbial and Phenyl Acid Dynamics during the Start-up Phase of Anaerobic Straw Degradation in Meso- and Thermophilic Batch Reactors. Microorganisms. 2019; 7(12):657. https://doi.org/10.3390/microorganisms7120657
Chicago/Turabian StylePrem, Eva Maria, Rudolf Markt, Nina Lackner, Paul Illmer, and Andreas Otto Wagner. 2019. "Microbial and Phenyl Acid Dynamics during the Start-up Phase of Anaerobic Straw Degradation in Meso- and Thermophilic Batch Reactors" Microorganisms 7, no. 12: 657. https://doi.org/10.3390/microorganisms7120657