Fungal and Bacterial Communities Exhibit Consistent Responses to Reversal of Soil Acidification and Phosphorus Limitation over Time

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site Description
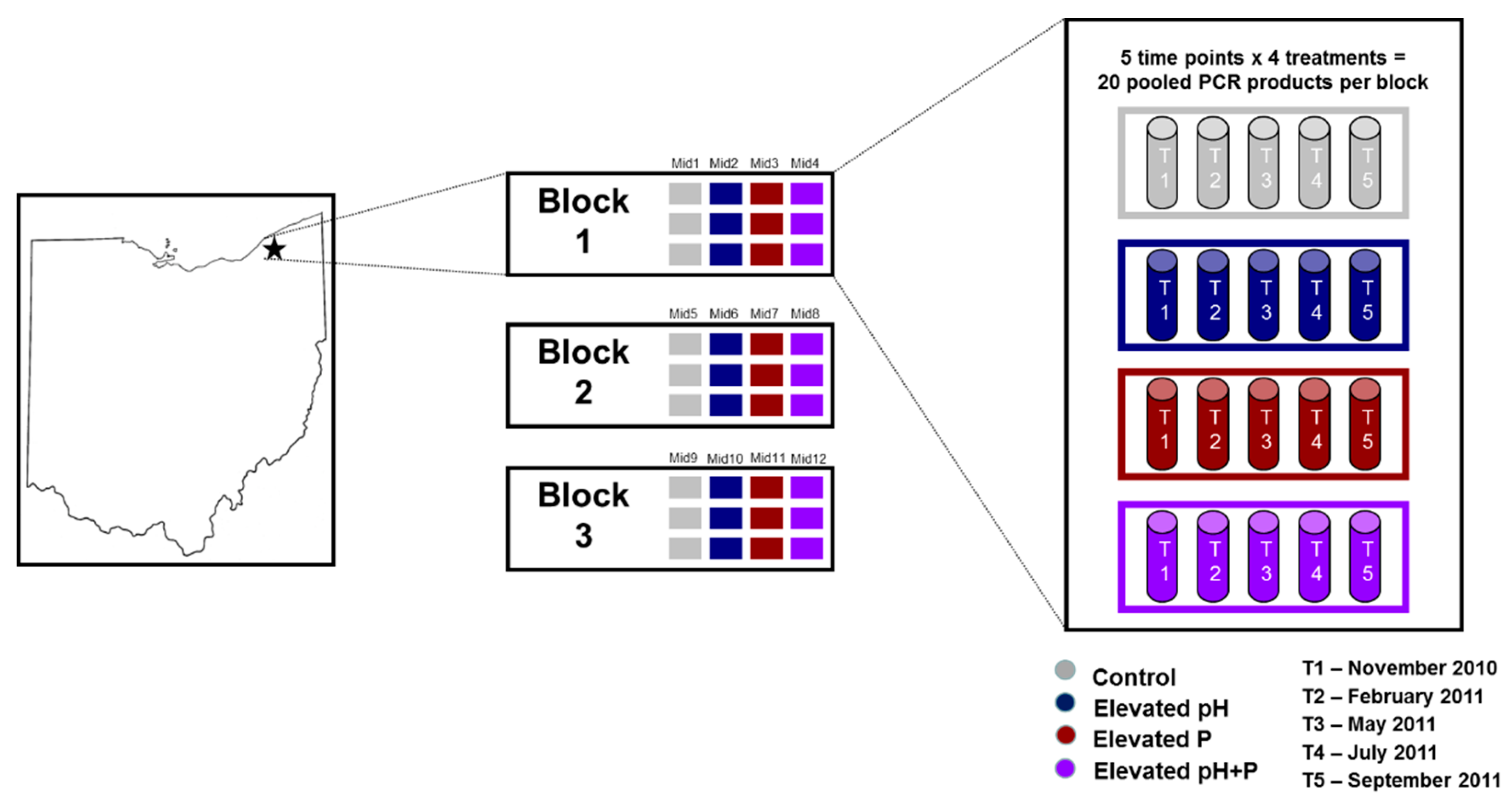
2.2. Experimental Design
2.3. Soil Sampling
2.4. DNA Extraction
2.5. PCR
2.6. 454 Sequencing
2.7. Sequence Analysis
2.8. Statistical Analysis
3. Results
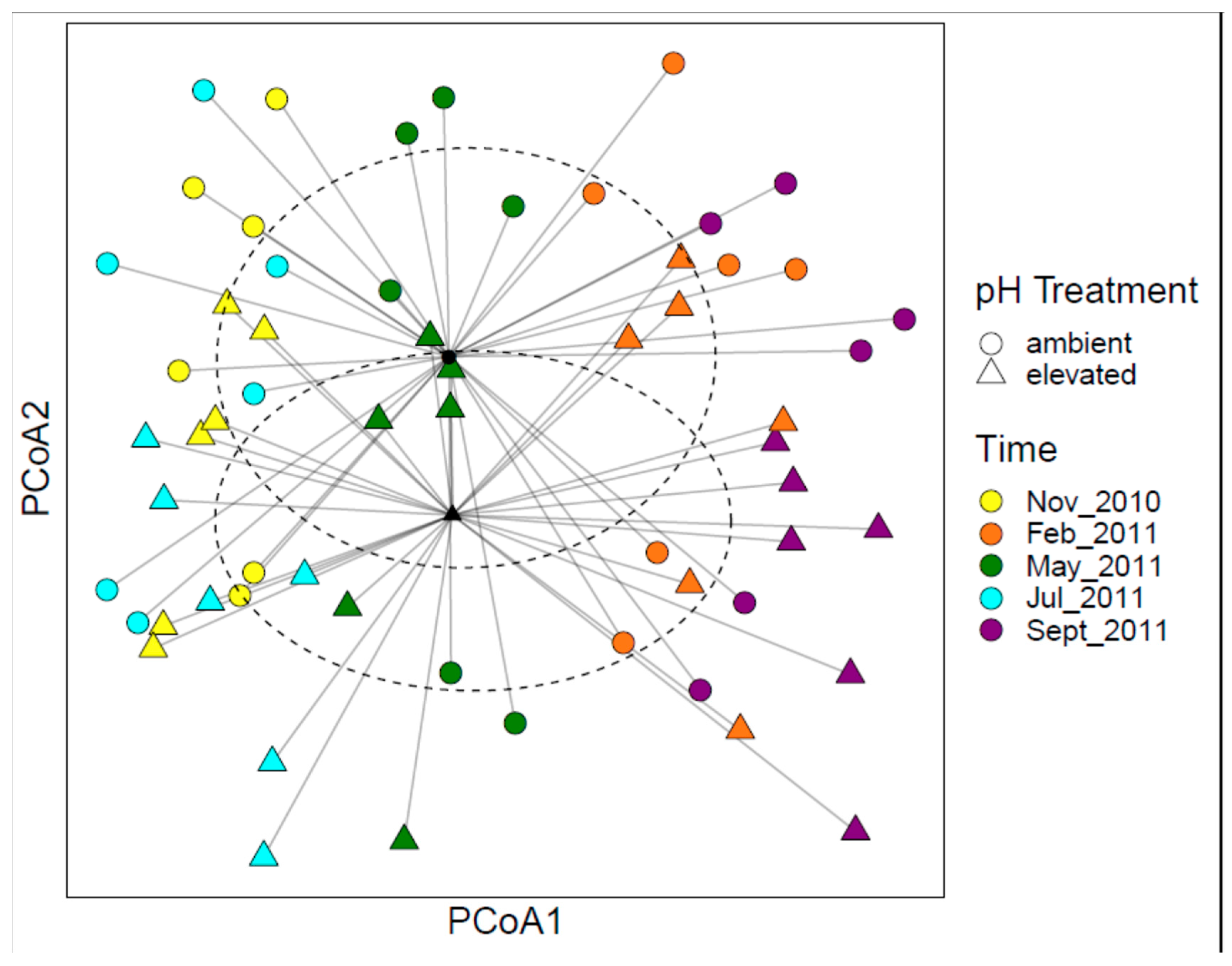
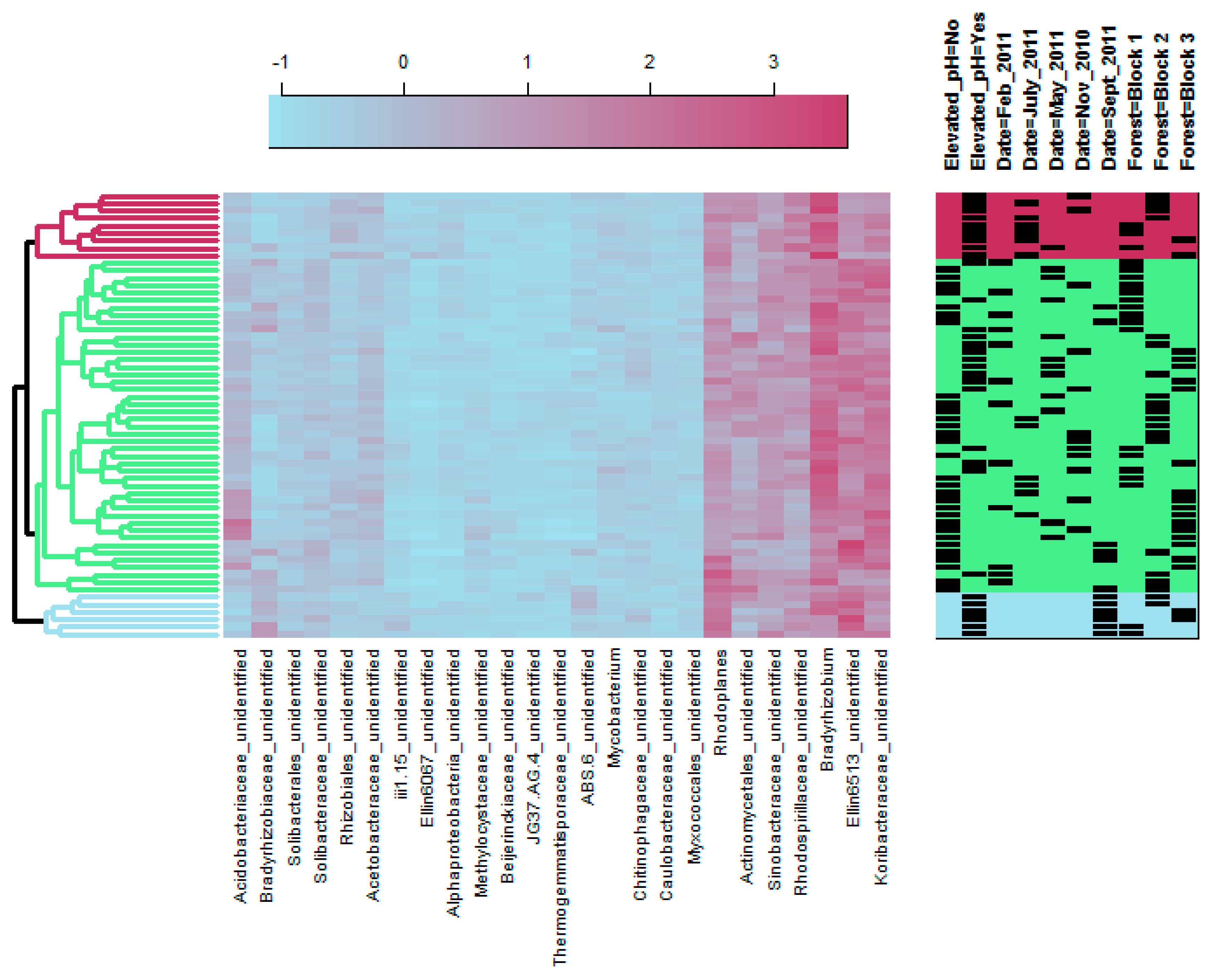
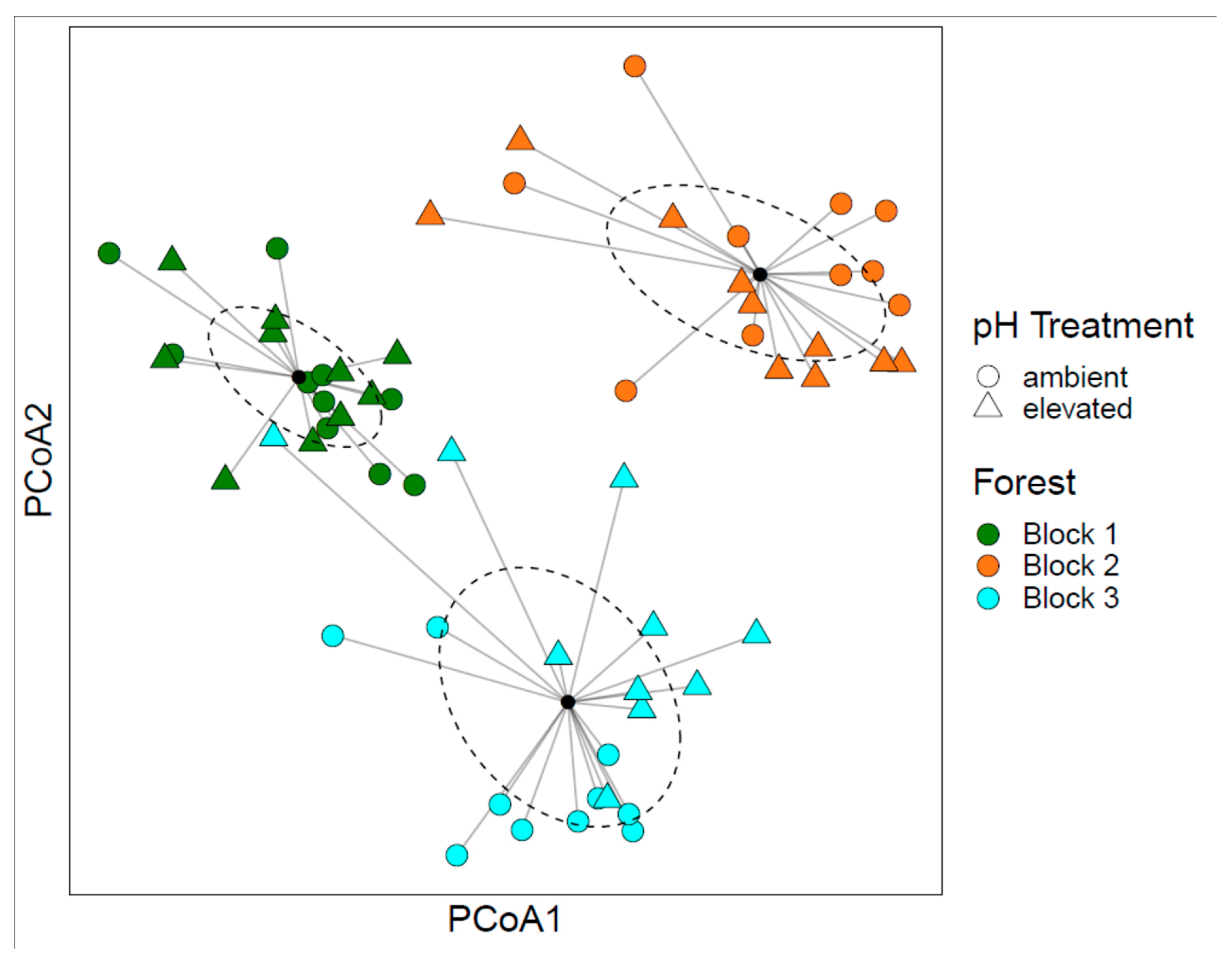
3.1. Bacterial Community Structure
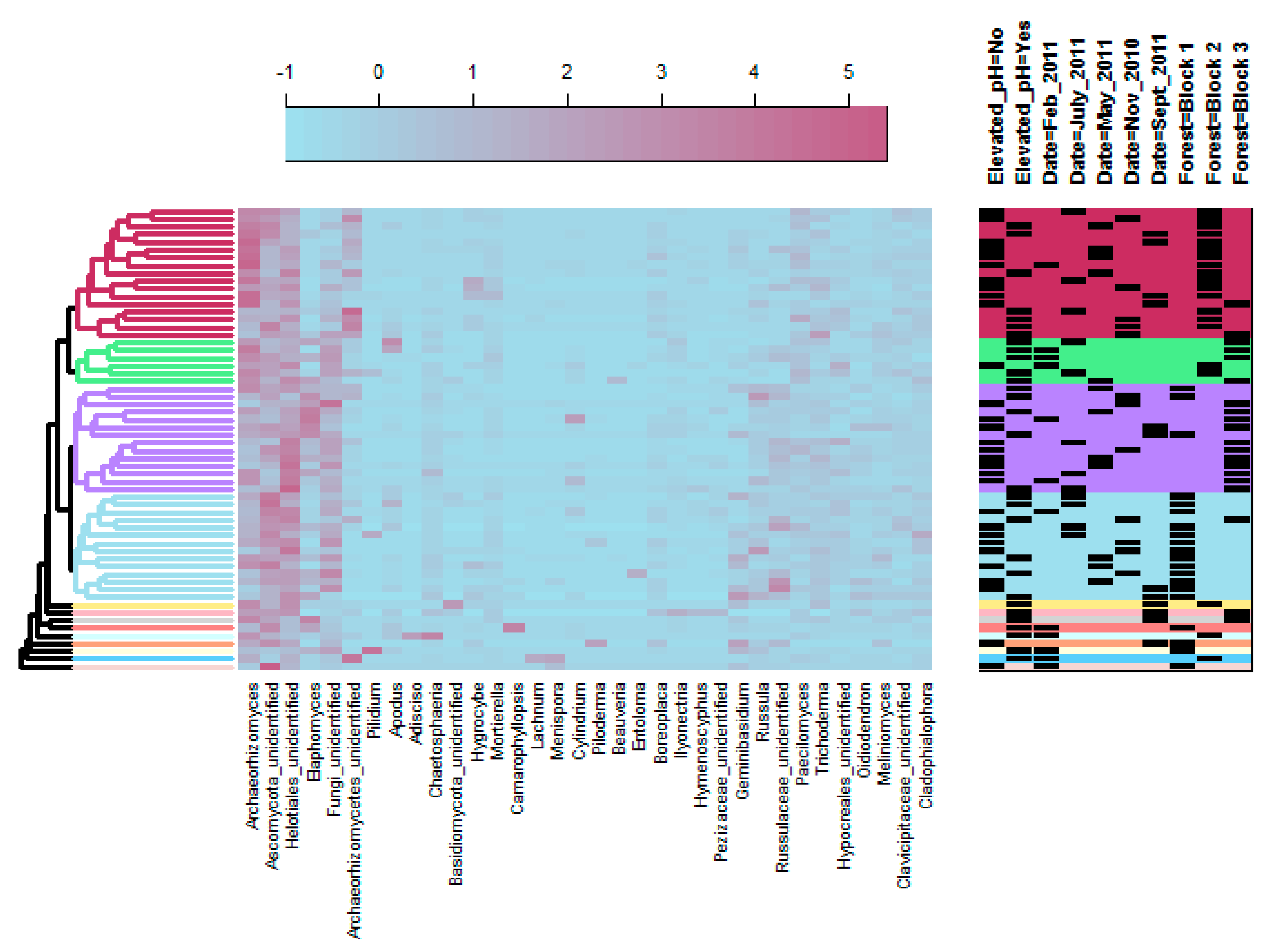
3.2. Fungal Community Structure
4. Discussion
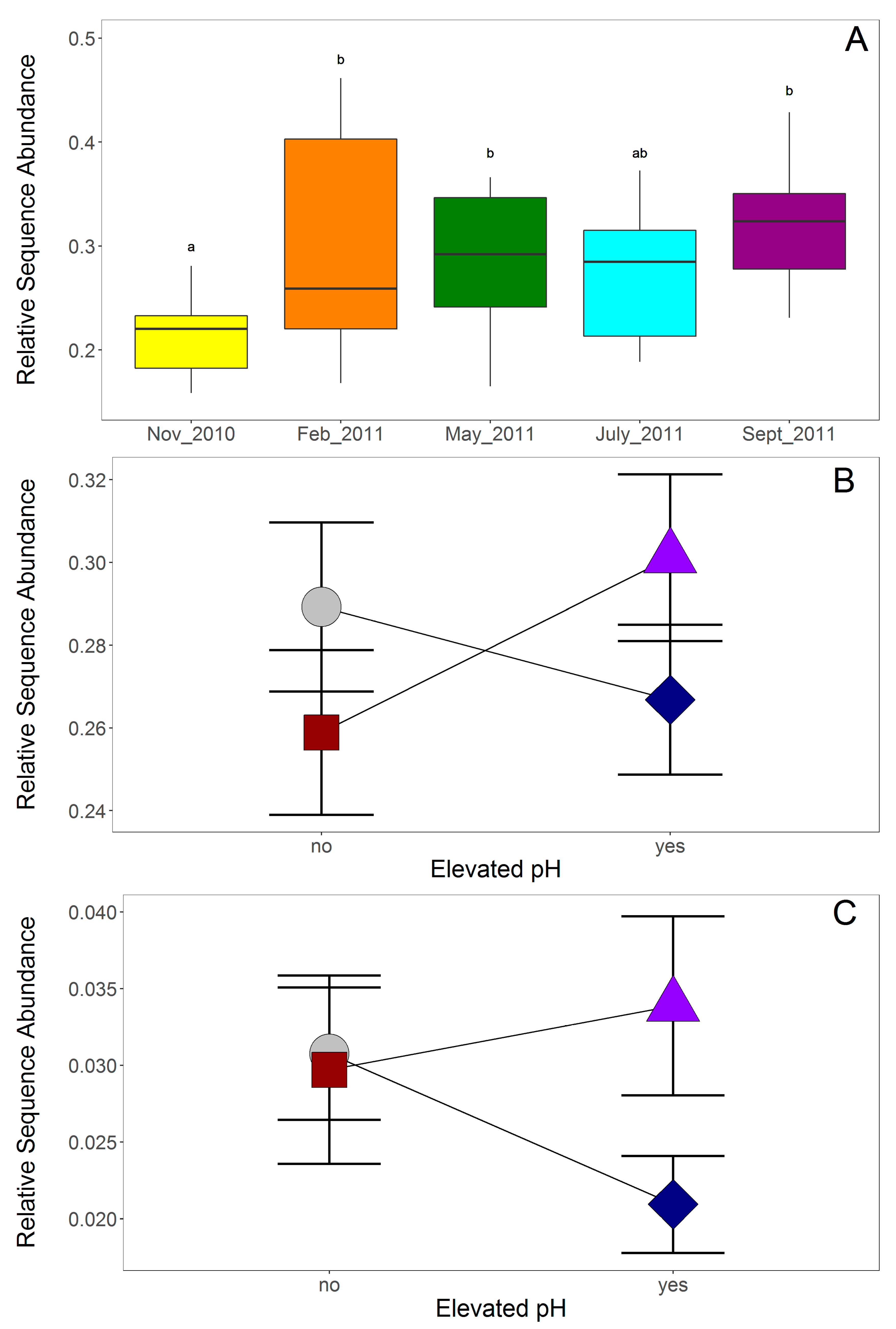
4.1. Fungal Community Response to pH Treatment
4.2. Bacterial Community Response to pH Treatment
4.3. Microbial Community Response to P Treatment
4.4. Summary
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the world’s forests. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar] [CrossRef] [Green Version]
- Bellassen, V.; Luyssaert, S. Carbon sequenstration: Managing forests in uncertain times. Nature 2014, 506, 153–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinley, D.C.; Ryan, M.G.; Birdsey, R.A.; Giardina, C.P.; Harmon, M.E.; Heath, L.S.; Houghton, R.A.; Jackson, R.B.; Morrison, J.F.; Murray, B.C.; et al. A synthesis of current knowledge on forests and carbon storage in the United States. Ecol. Appl. 2011, 21, 1902–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andren, H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review. Oikos 1994, 71, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Dupouey, J.-L.; Dambrine, E.; Laffite, J.D.; Moares, C. Irreversible impact of past land use on forest soils and biodiversity. Ecology 2002, 83, 2978–2984. [Google Scholar] [CrossRef]
- Johnson, N.M.; Driscoll, C.T.; Eaton, J.S.; Likens, G.E.; McDowell, W.H. ‘Acid-Rain’, dissolved aluminum and chemical weathering at the Hubbard Brook Experimental Forest, New Hampshire. Geochim. Cosmochim. Acta 1981, 45, 1421–1437. [Google Scholar] [CrossRef]
- Lovett, G.M.; Goodale, C.L. A new conceptual model of nitrogen saturation based on experimental nitrogen addition to an oak forest. Ecosystems 2011, 14, 615–631. [Google Scholar] [CrossRef]
- USDA Forest Service. U.S. Forest Facts and Historical Trends (FS-696); U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2001. [Google Scholar]
- Aber, J.D.; Goodale, C.L.; Ollinger, S.V.; Smith, M.-L.; Magill, A.H.; Martin, M.E.; Hallett, R.A.; Stoddard, J.L. Is nitrogen deposition altering the nitrogen status of northeastern forests? BioScience 2003, 53, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Farrington, H. Nutrient limitation and soil development: Experimental test of a biogeochemical theory. Biogeochemistry 1997, 37, 63–75. [Google Scholar] [CrossRef]
- Goldberg, S.; Davis, J.A.; Hem, J.D. The surface chemistry of aluminum oxides and hydroxides. In The Environmental Chemistry of Aluminum; Sposito, G., Ed.; CRC Lewis Publishers: Boca Raton, FL, USA, 1996; pp. 271–331. [Google Scholar]
- Booth, I.R. Regulation of cytoplasmic pH in bacteria. Microbiol. Rev. 1985, 49, 359–378. [Google Scholar]
- Quivey, R.G., Jr.; Faustoferri, R.; Monahan, K.; Marquis, R. Shifts in membrane fatty acid profiles associated with acid adaptation of Streptococcus mutans. FEMS Microbiol. Lett. 2000, 189, 89–92. [Google Scholar] [CrossRef] [PubMed]
- McGill, W.; Cole, C. Comparative aspects of cycling of organic C, N, S, and P through soil organic matter. Geoderma 1981, 26, 267–286. [Google Scholar] [CrossRef]
- Olander, L.P.; Vitousek, P.M. Biological and geochemical sinks for phosphorus in soil from a wet tropical forest. Ecosystems 2004, 7, 404–419. [Google Scholar] [CrossRef]
- Traina, S.J.; Sposito, G.; Hesterberg, D.; Kafkafi, U. Effects of pH and organic acids on ortho-phosphate solubility in an acidic, montmorillonitic soil. Soil Sci. Soc. Am. J. 1986, 50, 45–52. [Google Scholar] [CrossRef]
- Hartman, W.H.; Richardson, C.J. Differential nutrient limitation of soil microbial biomass and metaboloic quotients (qCO2): Is there a biological stoichiometry of soil microbes? PLoS ONE 2013, 8, e57127. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.J.; López-Gutiérrez, J.C.; Smemo, K.A.; Chan, C.R. Vegetation and soil environment influence the spatial distribution of root-associated fungi in a mature beech-maple forest. Appl. Env. Microbiol. 2009, 75, 7639–7648. [Google Scholar] [CrossRef] [Green Version]
- DeForest, J.L.; Scott, L.B. Available organic soil phosphorus has in important influence on microbial community composition. Soil Sci. Soc. Am. J. 2010, 74, 2059–2066. [Google Scholar] [CrossRef]
- Kluber, L.A.; Carrino-Kyker, S.R.; Coyle, K.P.; DeForest, J.L.; Hewins, C.R.; Shaw, A.N.; Smemo, K.A.; Burke, D.J. Mycorrhizal response to experimental pH and P manipulation in acidic hardwood forests. PLoS ONE 2012, 7, e48946. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Morris, M.H.; Smith, M.E.; Rizzo, D.M.; Rejmánek, M.; Bledsoe, C.S. Contrasting ectomycorrhizal fungal communities on the roots of co-occurring oaks (Quercus spp.) in a California woodland. New Phytol. 2008, 178, 167–176. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Bååth, E.; Anderson, T.H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrino-Kyker, S.R.; Kluber, L.A.; Petersen, S.M.; Coyle, K.P.; Hewins, C.R.; DeForest, J.L.; Smemo, K.A.; Burke, D.J. Mycorrhizal fungal communities respond to experimental elevation of soil pH and P availability in temperate hardwood forests. FEMS Microbiol. Ecol. 2016, 92, fiw024. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, I.; Fahey, T.J.; Groffman, P.M.; Driscoll, C.T.; Eagar, C.; Siccama, T.G. Initial responses of phosphorus biogeochemistry to calcium addition in a northern hardwood forest ecosystem. Can. J. Res. 2003, 33, 1864–1873. [Google Scholar] [CrossRef]
- Gress, S.E.; Nichols, T.D.; Northcraft, C.C.; Peterjohn, W.T. Nutrient limitation in soils exhibiting differing nitrogen availabilities: What lies beyond nitrogen saturation? Ecology 2007, 88, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 2005, 20, 634–641. [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchsleuger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Merino, C.; Nannipieri, P.; Matus, F. Soil carbon controlled by plant, microorganisms and mineralogy interactions. J. Soil Sci. Plant Nutr. 2015, 15, 321–332. [Google Scholar] [CrossRef] [Green Version]
- DeForest, J.L.; Smemo, K.A.; Burke, D.J.; Elliott, H.L.; Becker, J.C. Soil microbial responses to elevated phosphorus and pH in acidic temperate deciduous forests. Biogeochemistry 2012, 109, 189–202. [Google Scholar] [CrossRef]
- Maraun, M.; Scheu, S. Changes in microbial biomass, respiration and nutrient status of beech (Fagus sylvatica) leaf litter processed by millipedes (Glomeris marginata). Oecologia 1996, 107, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T. ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Hamady, M.; Lauber, C.L.; Knight, R. The influence of sex, handedness, and washing on the diversity of hand surface bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17994–17999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenberg, D.; Von Kuster, G.; Coraor, N.; Ananda, G.; Lazarus, R.; Mangan, M.; Nekrutenko, A.; Taylor, J. Galaxy: A web-based genome analysis tool for experimentalists. Curr. Protoc. Mol. Biol. 2010, 19, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Giardine, B.; Riemer, C.; Hardison, R.C.; Burhans, R.; Elnitski, L.; Shah, P.; Zhang, Y.; Blankenberg, D.; Albert, I.; Taylor, J.; et al. Galaxy: A platform for interactive large-scale genome analysis. Genome Res. 2005, 15, 1451–1455. [Google Scholar] [CrossRef] [Green Version]
- Goecks, J.; Nekrutenko, A.; Taylor, J.; Galaxy Team. Galaxy: A comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010, 11, R86. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.-H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, T.B.K.; Thomas, A.D.; Stamatis, D.; Bertsch, J.; Isbandi, M.; Jansson, J.; Mallajosyula, J.; Pagani, I.; Lobos, E.A.; Kyrpides, N.C. The Genomes OnLine Database (GOLD) v.5: A metadata management system based on a four level (meta)genome project classification. Nucleic Acids Res. 2015, 43, 1099–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Werner, J.; Koren, O.; Hugenholtz, P.; DeSantis, T.Z.; Walters, W.A.; Caporaso, J.G.; Angenent, L.T.; Knight, R.; Ley, R.E. Impact of training sets on classification of high-throughput bacterial 16S rRNA gene surveys. ISME J. 2012, 6, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Dickie, I. Insidious effects of sequencing errors on perceived diversity in molecular surveys. New Phytol. 2010, 188, 916–918. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, v2.5.5. 2013. Available online: http://CRAN.R-project.org/package=vegan.
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/.
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploner, A. Heatplus: Heatmaps with Row and/or Column Covariates and Colored Clusters, v2.28.0. 2019. Available online: https://github.com/alexploner/Heatplus.
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2015, 20, 241–248. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models, v3.1.137. 2019. Available online: https://CRAN.R-project.org/package=nlme.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- McArthur, J.V. Microbial Ecology: An Evolutionary Approach; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry, Chemical Equilibria and Rates in Natural Waters, 3rd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Walker, L.; Syers, J. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Madigan, M.T.; Martinko, J.M.; Parker, J. (Eds.) Brock Biology of Microorganisms; Pearson Education, Inc.: Upper Saddle River, NJ, USA, 2003. [Google Scholar]
- Erland, S.; Söderström, B. Effects of liming on ectomycorrhizal fungi infecting Pinus sylvestris L. I. mycorrhizal infection in limed humus in the laboratory and isolation of fungi from mycorrhizal roots. New Phytol. 1990, 115, 675–682. [Google Scholar] [CrossRef]
- Lehto, T. Effects of liming and boron fertilization on mycorrhizas of Picea abies. Plant Soil 1994, 163, 65–68. [Google Scholar] [CrossRef]
- Rineau, F.; Maurice, J.-P.; Nys, C.; Voiry, H.; Garbaye, J. Forest liming durably impact the communities of ectomycorrhizas and fungal epigeous fruiting bodies. Ann. Sci. 2010, 67, 110. [Google Scholar] [CrossRef] [Green Version]
- Wubet, T.; Christ, S.; Schöning, I.; Boch, S.; Gawlich, M.; Schnabel, B.; Fischer, M.; Buscot, F. Differences in soil fungal communities between European beech (Fagus sylvatica L.) dominated forests are related to soil and understory vegetation. PLoS ONE 2012, 7, e47500. [Google Scholar] [CrossRef]
- Kottke, I.; Weber, R.; Ritter, T.; Oberwinkler, F. Activity of mycorrhizas and health status of trees on diverse forest stands in West Germany. In Forest Decline in the Atlantic and Pacific Regions; Hüttl, R.F., Mueller-Dombois, D., Eds.; Springer: Berlin, Germany, 1993; pp. 189–201. [Google Scholar]
- Bödeker, I.T.M.; Lindahl, B.D.; Olson, A.; Clemmensen, K.E. Mycorrhizal and saprotrophic fungal guilds compete for the same organic substrates but affect decomposition differently. Funct. Ecol. 2016, 30, 1967–1978. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Boddy, L.; Jones, T.H. Functional and ecological consequences of saprotrophic fungus-grazer interactions. ISME J. 2012, 6, 1992–2001. [Google Scholar] [CrossRef] [Green Version]
- Izzo, A.; Agbowo, J.; Bruns, T.D. Detection of plot-level changes in ectomycorrhizal communities across years in an old-growth mixed-conifer forest. New Phytol. 2005, 166, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Figueroa, E.A.; Seddon, E.; Yashiro, E.; Buri, A.; Niculita-Hirzel, H.; van der Meer, J.R.; Guisan, A. Archaeorhizomycetes spatial distribution in soils along wide elevational and environmental gradients reveal co-abundance patterns with other fungal saprobes and potential weathering capacities. Front. Microbiol. 2019, 10, 656. [Google Scholar] [CrossRef] [PubMed]
- Saitta, A.; Anslan, S.; Bahram, M.; Brocca, L.; Tedersoo, L. Tree species identity and diversity drive fungal richness and community composition along an elevational gradient in a Mediterranean ecosystem. Mycorrhiza 2018, 28, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef] [PubMed]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Zimmerman, N.B.; Carbone, I.; May, G.; Arnold, A.E. Host availability drives distributions of fungal endophytes in the imperiled boreal realm. Nat. Ecol. Evol. 2019, 3, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Suvi, T.; Jairus, T.; Kõljalg, U. Forest microsite effects on community composition of ectomycorrhizal fungi on seedlings of Picea albies and Betula pendula. Environ. Microbiol. 2008, 10, 1189–1201. [Google Scholar] [CrossRef]
- Burke, D.J. Effects of annual and inter-annual environmental variability on soil fungi associated with an old-growth, temperate hardwood forest. FEMS Microbiol. Ecol. 2015, 91, fiv053. [Google Scholar] [CrossRef] [Green Version]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Hackl, E.; Zechmeister-Boltenstern, S.; Bodrossy, L.; Sessitsch, A. Comparison of diversities and compositions of bacterial populations inhabiting natural forest soils. Appl. Environ. Microbiol. 2004, 70, 5057–5065. [Google Scholar] [CrossRef] [Green Version]
- Bartram, A.K.; Jiang, X.; Lynch, M.D.J.; Masella, A.P.; Nicol, G.W.; Dushoff, J.; Neufeld, J.D. Exploring links between pH and bacterial community composition in soils from the Craibstone Experimental Farm. FEMS Microbiol. Ecol. 2013, 87, 403–415. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Groffman, P.M.; Fisk, M.C. Phosphate additions have no effect on microbial biomass and activity in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 2441–2449. [Google Scholar] [CrossRef]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil micorbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Martino, E.; Morin, E.; Greler, G.-A.; Kuo, A.; Kohler, A.; Daghino, S.; Barry, K.W.; Cichocki, N.; Clum, A.; Dockter, R.B.; et al. Comparative genomics and transcriptomics depict ericoid mycorrhizal fungi as versatile saprotrophs and plant mutualists. New Phytol. 2018, 217, 1213–1229. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| df† | SS† | Mean Sqs† | F-Value | R2 | p-Value * | |
|---|---|---|---|---|---|---|
| pH | 1 | 0.51 | 0.51 | 3.99 | 0.048 | <0.001 |
| P | 1 | 0.13 | 0.13 | 0.98 | 0.012 | 0.26 |
| Time | 4 | 3.46 | 0.87 | 6.78 | 0.33 | <0.001 |
| pH:P | 1 | 0.15 | 0.15 | 1.17 | 0.014 | 0.14 |
| pH:Time | 4 | 0.56 | 0.14 | 1.10 | 0.053 | 0.098 |
| P:Time | 4 | 0.33 | 0.083 | 0.65 | 0.032 | 0.97 |
| pH:P:Time | 4 | 0.34 | 0.085 | 0.67 | 0.032 | 0.96 |
| Residuals | 80 | 5.11 | 0.13 | 0.48 | ||
| Total | 119 | 10.60 | 1 |
| df† | SS† | Mean Sqs† | F-Value | R2 | p-Value * | |
|---|---|---|---|---|---|---|
| pH | 1 | 0.49 | 0.49 | 1.53 | 0.025 | <0.001 |
| P | 1 | 0.28 | 0.28 | 0.87 | 0.014 | 0.27 |
| Time | 4 | 3.32 | 0.83 | 2.59 | 0.17 | <0.001 |
| pH:P | 1 | 0.24 | 0.24 | 0.76 | 0.012 | 0.67 |
| pH:Time | 4 | 0.94 | 0.23 | 0.73 | 0.047 | 0.94 |
| P:Time | 4 | 0.86 | 0.21 | 0.67 | 0.043 | 1.00 |
| pH:P:Time | 4 | 0.82 | 0.20 | 0.64 | 0.041 | 0.99 |
| Residuals | 40 | 12.82 | 0.32 | 0.65 | ||
| Total | 59 | 19.77 | 1 |
| numDF† | denDF† | F-Value | p-Value * | |
|---|---|---|---|---|
| Saprotroph | ||||
| pH | 1 | 38 | 0.47 | 0.50 |
| P | 1 | 38 | 0.019 | 0.89 |
| Time | 4 | 38 | 6.57 | <0.01 |
| pH:P | 1 | 38 | 5.02 | 0.031 |
| pH:Time | 4 | 38 | 2.56 | 0.054 |
| P:Time | 4 | 38 | 0.096 | 0.98 |
| pH:P:Time | 4 | 38 | 1.38 | 0.26 |
| Ericoid Mycorrhizal Fungi | ||||
| pH | 1 | 37 | 0.012 | 0.91 |
| P | 1 | 37 | 0.70 | 0.41 |
| Time | 4 | 37 | 0.75 | 0.56 |
| pH:P | 1 | 37 | 4.71 | 0.036 |
| pH:Time | 4 | 37 | 0.32 | 0.86 |
| P:Time | 4 | 37 | 0.26 | 0.90 |
| pH:P:Time | 4 | 37 | 0.90 | 0.47 |
| Ectomycorrhizal Fungi | ||||
| pH | 1 | 38 | 0.23 | 0.64 |
| P | 1 | 38 | 0.80 | 0.38 |
| Time | 4 | 38 | 1.46 | 0.23 |
| pH:P | 1 | 38 | 0.026 | 0.87 |
| pH:Time | 4 | 38 | 0.15 | 0.96 |
| P:Time | 4 | 38 | 1.41 | 0.25 |
| pH:P:Time | 4 | 38 | 0.67 | 0.62 |
| Pathogen | ||||
| pH | 1 | 36 | 0.0013 | 0.97 |
| P | 1 | 36 | 0.85 | 0.36 |
| Time | 4 | 36 | 1.49 | 0.23 |
| pH:P | 1 | 36 | 2.24 | 0.14 |
| pH:Time | 4 | 36 | 0.41 | 0.80 |
| P:Time | 4 | 36 | 2.21 | 0.087 |
| pH:P:Time | 4 | 36 | 0.77 | 0.55 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
R. Carrino-Kyker, S.; P. Coyle, K.; A. Kluber, L.; J. Burke, D. Fungal and Bacterial Communities Exhibit Consistent Responses to Reversal of Soil Acidification and Phosphorus Limitation over Time. Microorganisms 2020, 8, 1. https://doi.org/10.3390/microorganisms8010001
R. Carrino-Kyker S, P. Coyle K, A. Kluber L, J. Burke D. Fungal and Bacterial Communities Exhibit Consistent Responses to Reversal of Soil Acidification and Phosphorus Limitation over Time. Microorganisms. 2020; 8(1):1. https://doi.org/10.3390/microorganisms8010001
Chicago/Turabian StyleR. Carrino-Kyker, Sarah, Kaitlin P. Coyle, Laurel A. Kluber, and David J. Burke. 2020. "Fungal and Bacterial Communities Exhibit Consistent Responses to Reversal of Soil Acidification and Phosphorus Limitation over Time" Microorganisms 8, no. 1: 1. https://doi.org/10.3390/microorganisms8010001