
The World’s First Acne Dysbiosis-like Model of Human 3D Ex Vivo Sebaceous Gland Colonized with Cutibacterium acnes and Staphylococcus epidermidis
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Biological Specimen Collection
2.2. Primary Human Sebocyts Isolation and 2D Monolayer Cell Culture
2.3. Optimization of Experimental Conditions Required for Primary Human Sebocyte Co-Culturing with C. acnes and S. epidermidis
2.3.1. Bacterial Strains Selection and Amplification
2.3.2. Primary Human Sebocyte Co-Culturing with Bacterial Strains
2.3.3. Analysis of the Impact of Sebocytes on the Survival of Bacteria
2.3.4. Analysis of the Impact of Bacteria on the Survival of Sebocytes
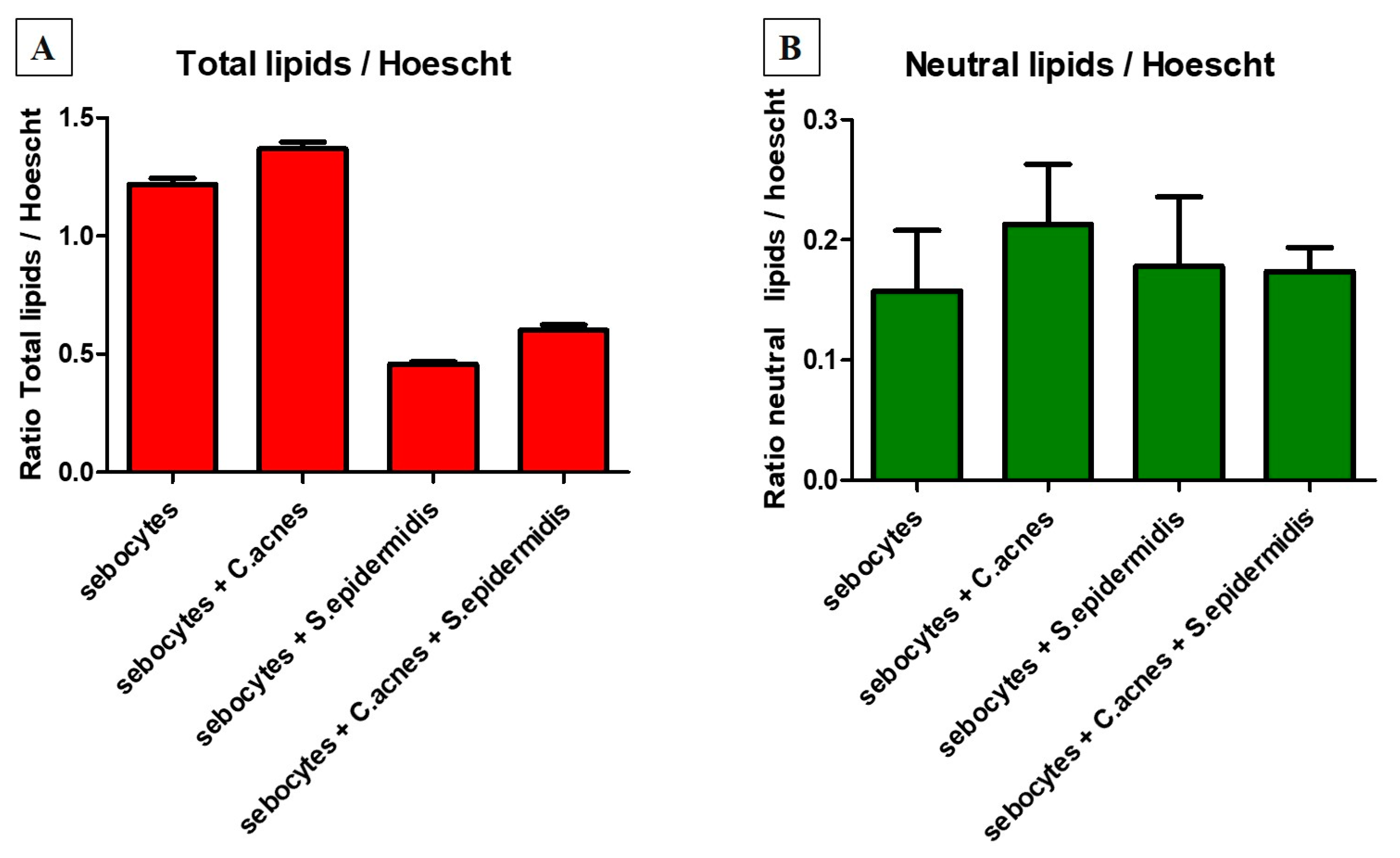
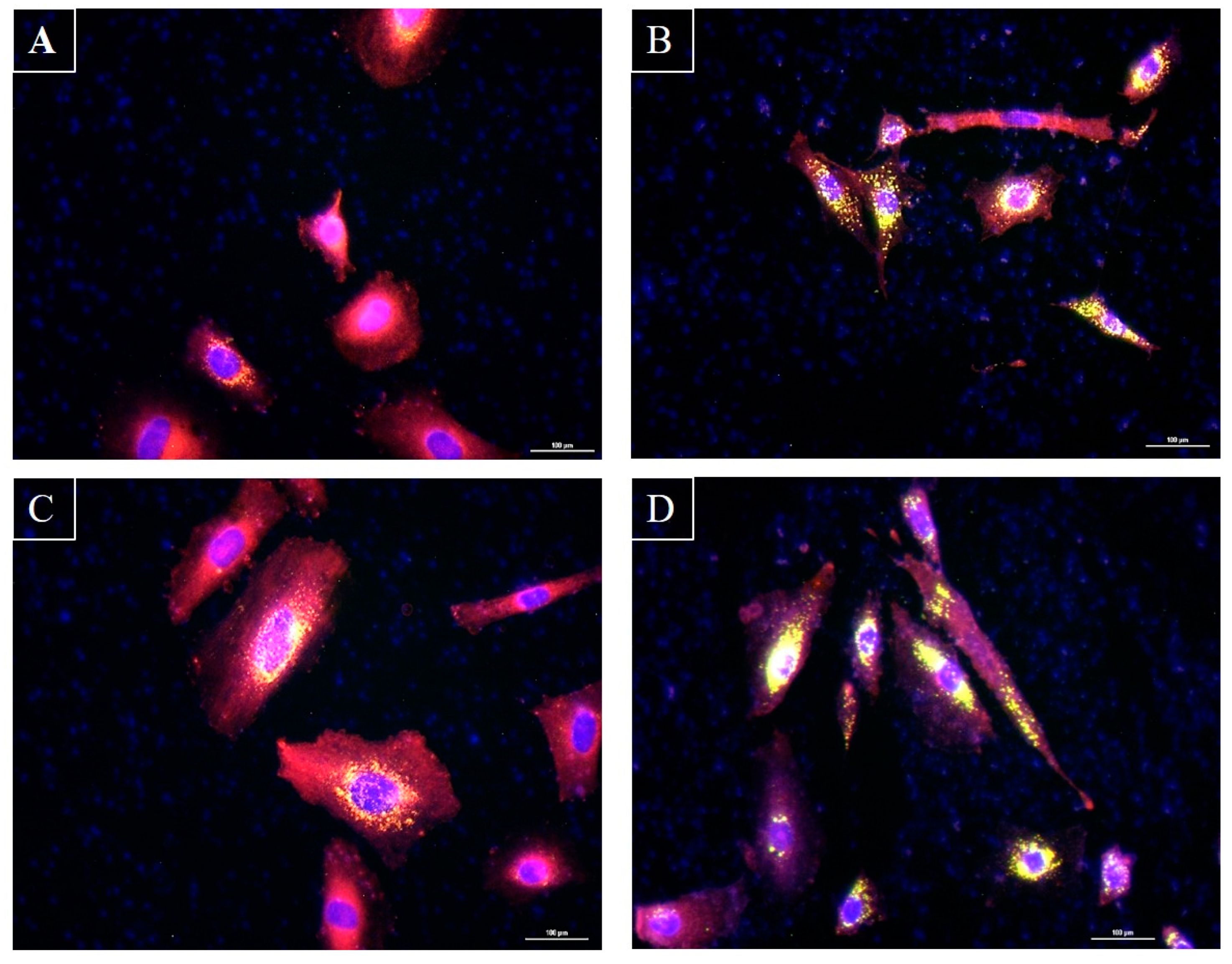
2.3.5. Analysis of the Impact of Bacteria on Lipid Production by Sebocytes
- Core (Hoechst): excitation = 356 nm and emission = 465 nm;
- Neutral lipids (Nile Red): excitation = 475 nm and emission = 530 nm;
- Total lipids (Nile Red): excitation = 520 nm and emission = 625 nm.
2.4. Optimization of Experimental Conditions for 3D Ex Vivo Sebaceous Glands Using C. acnes and S. epidermidis
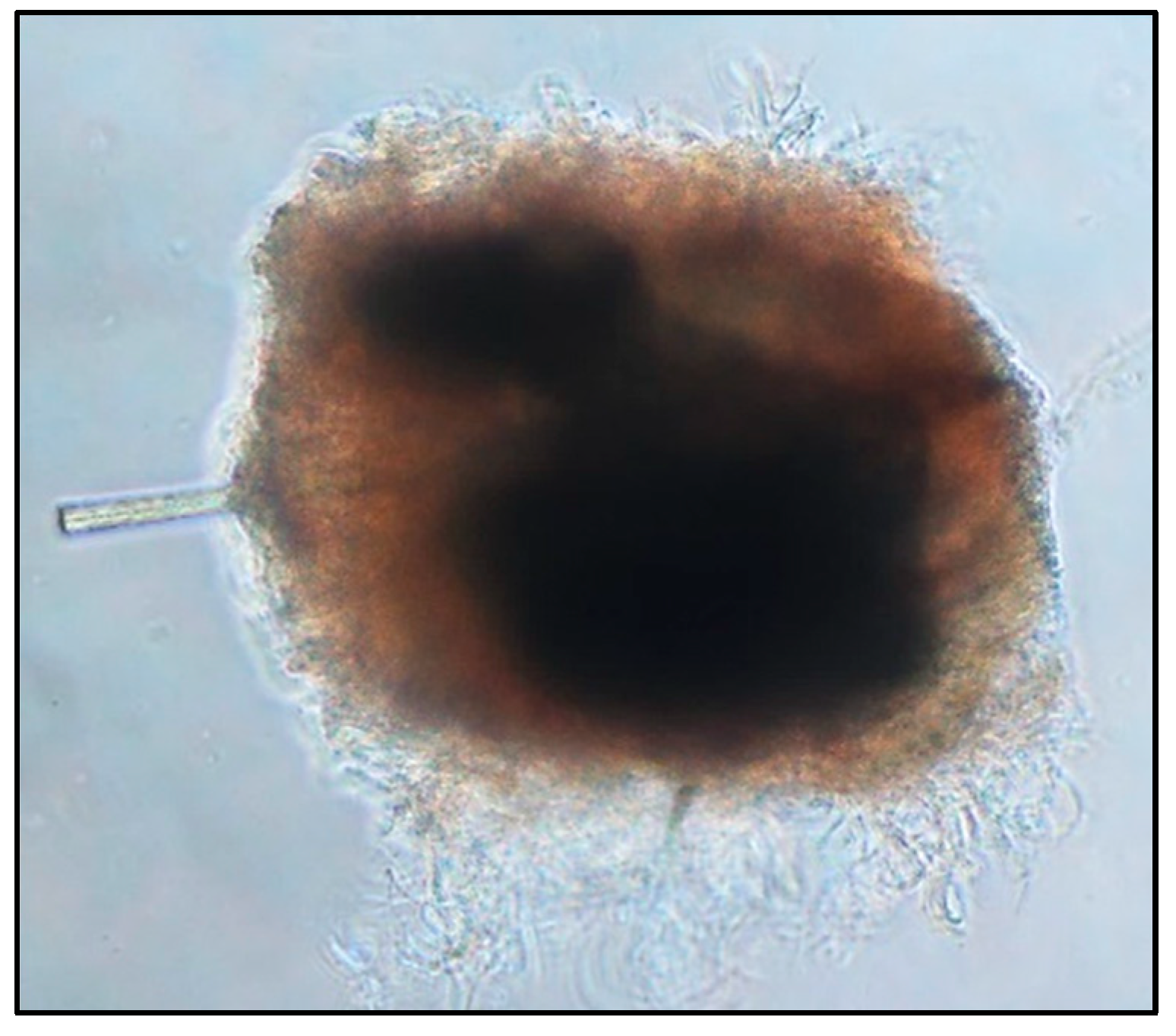
2.4.1. Sebaceous Gland Extraction, Culture and Viability Assessment
2.4.2. Inoculation of Bacteria on the 3D Sebaceous Gland Model
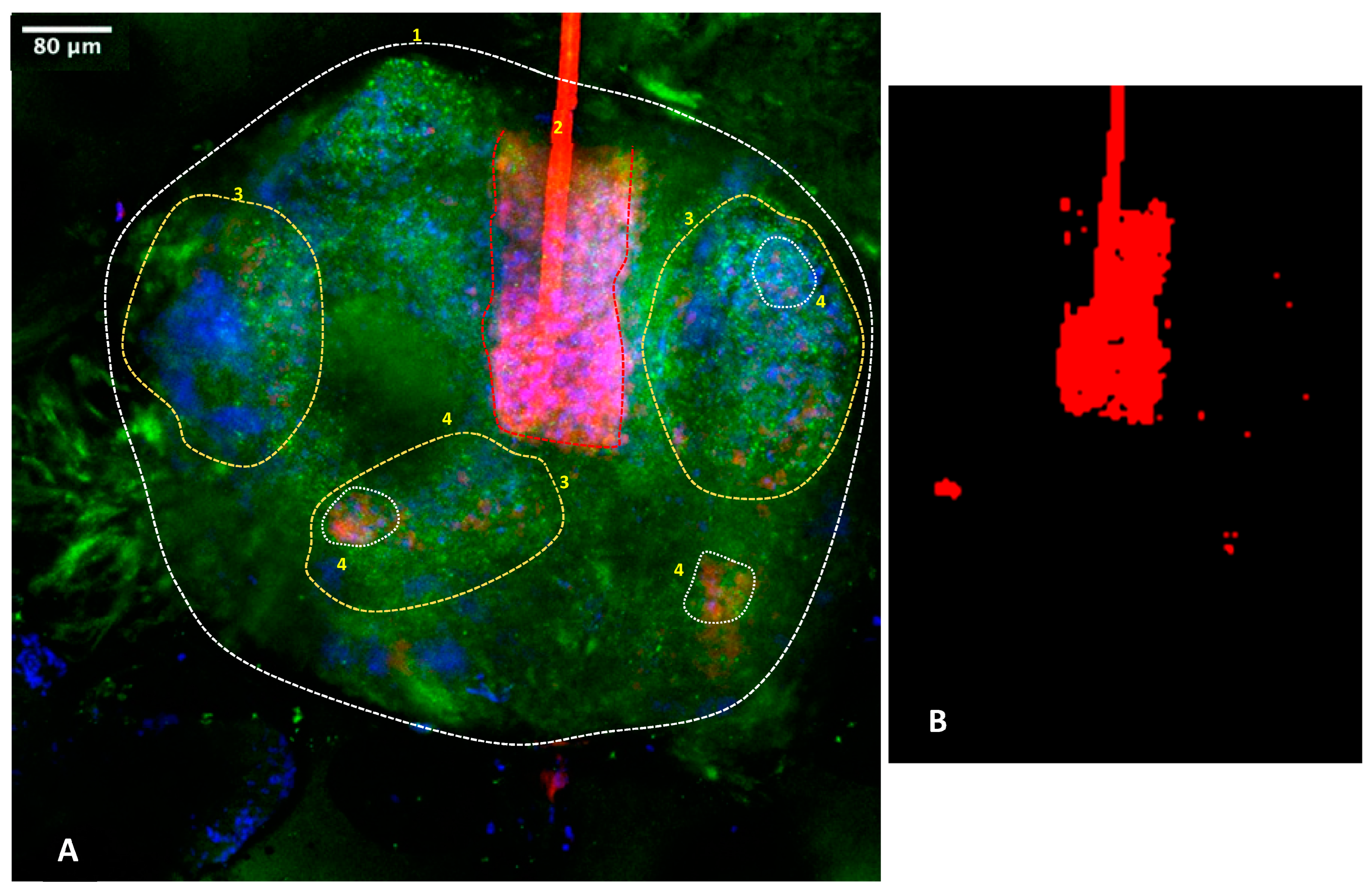
2.4.3. Bio-Imaging of Microbiota on Sebaceous Glands
Immunostaining
Confocal Microscopy Bioimaging
2.5. Statistics
3. Results
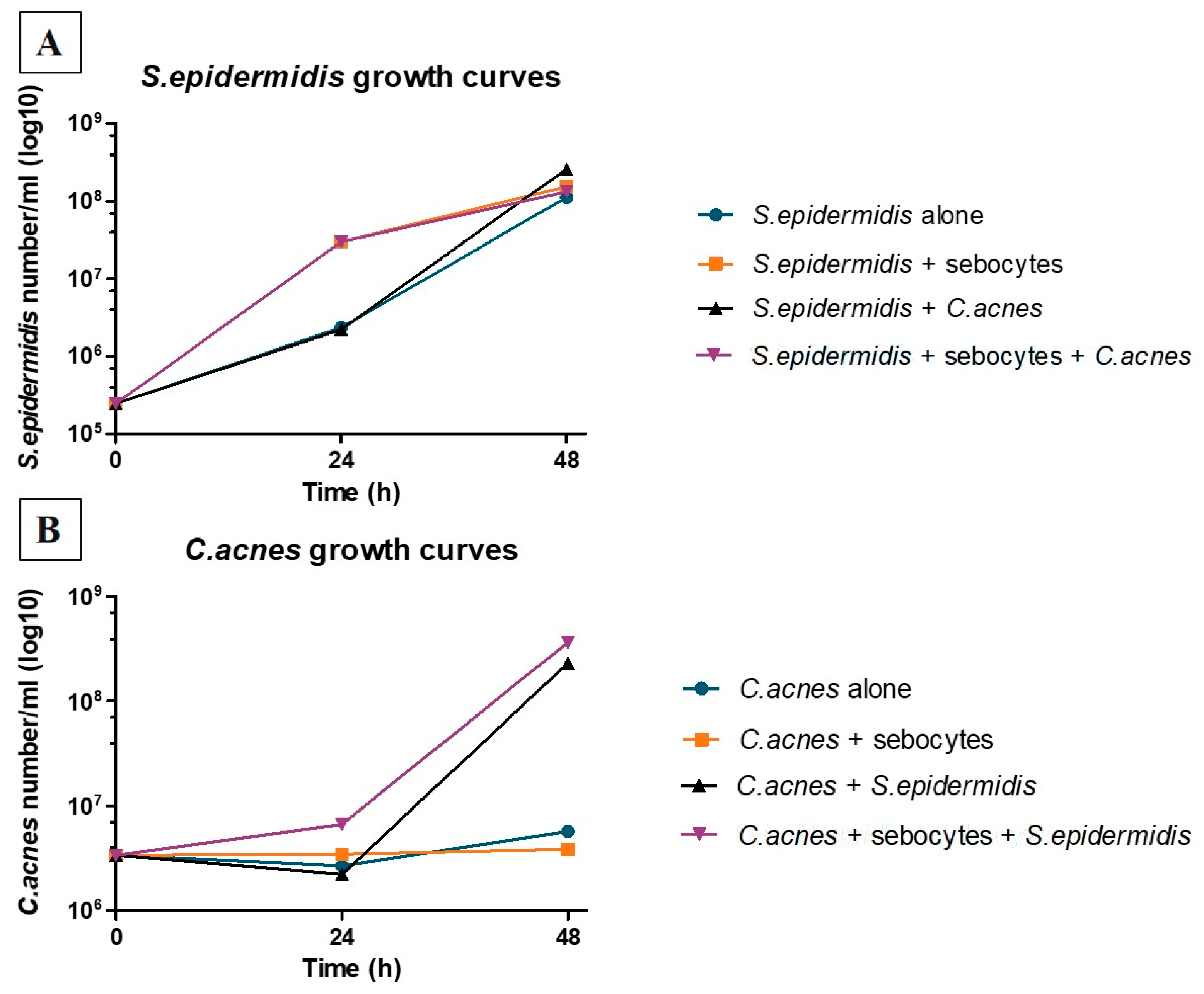
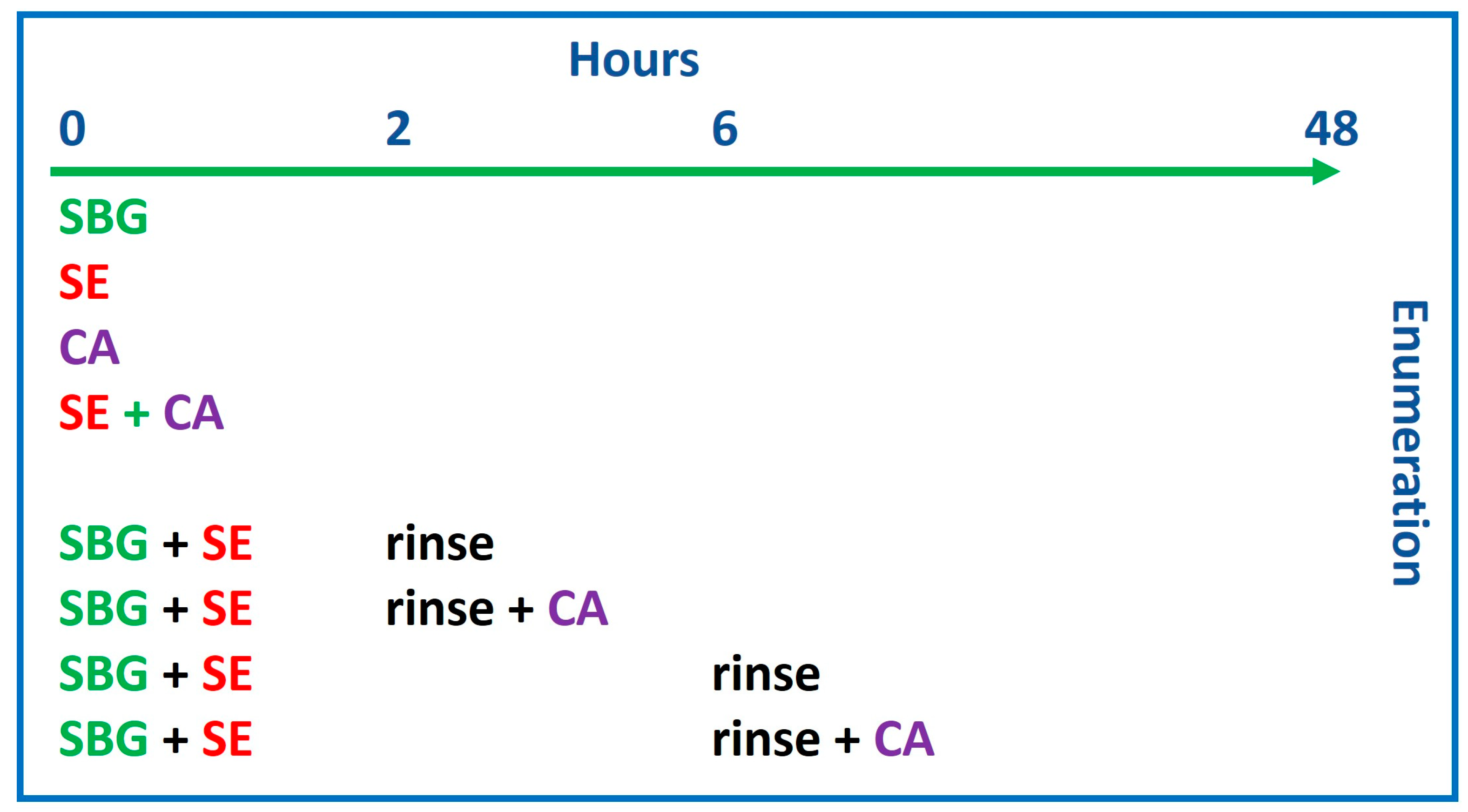
3.1. S. epidermidis Inoculum Must Be Adjusted to Prevent Overproliferation in the Presence of C. acnes and Sebocytes
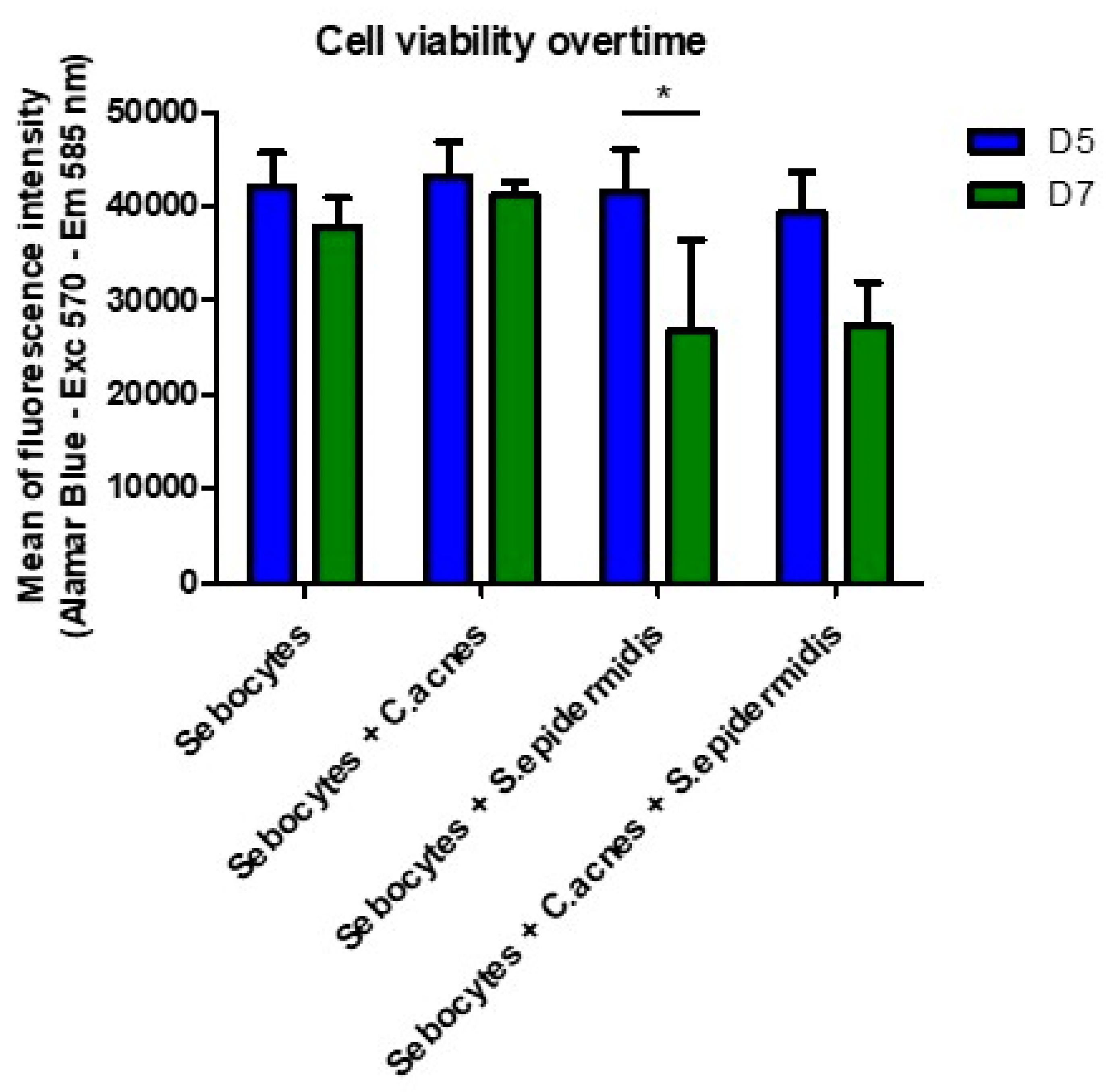
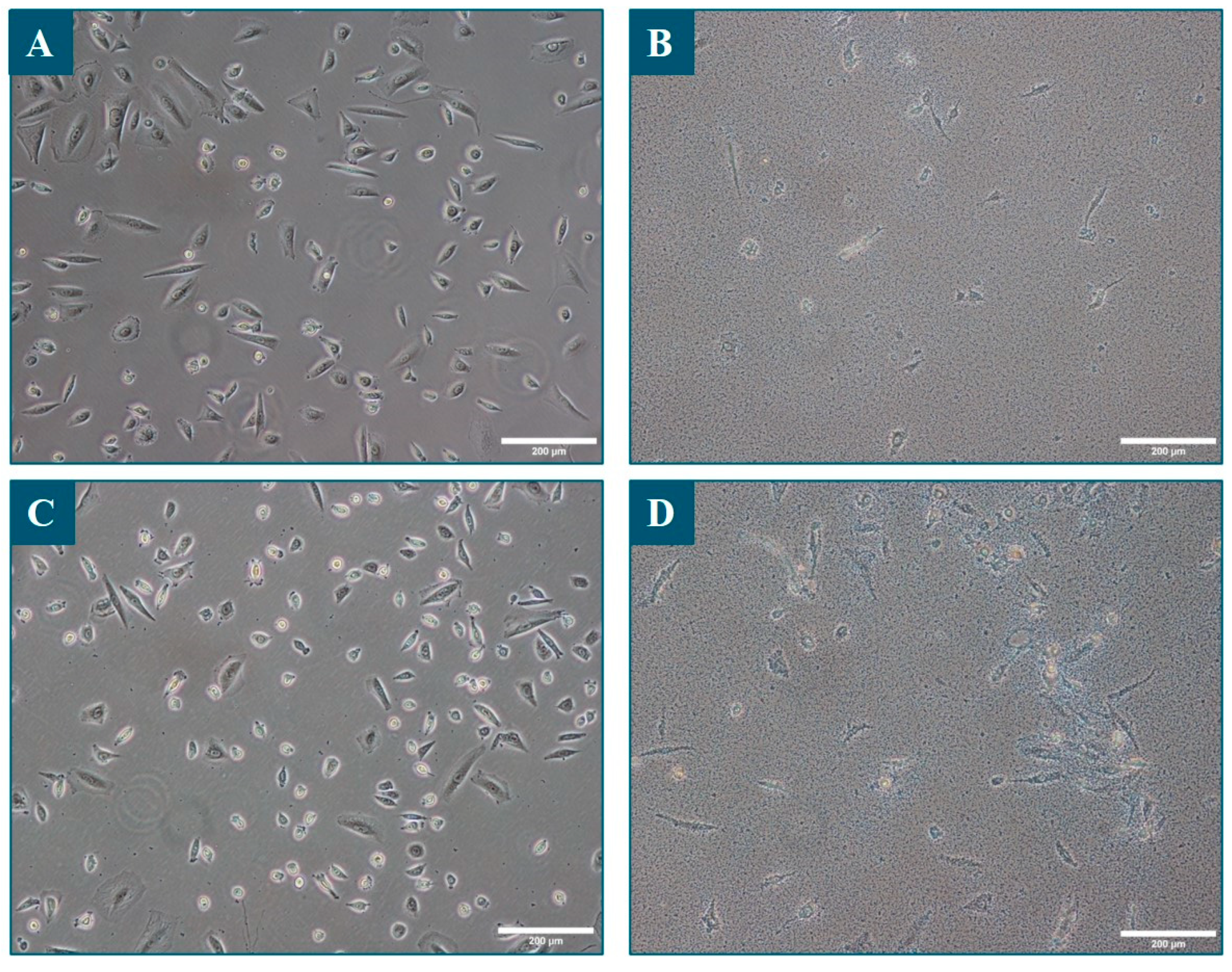
3.2. Human Sebocytes Viability and Metabolism Are Negatively Affected by the Overproliferation of S. epidermidis, but Not by C. acnes
3.3. C. acnes Induced Total and Neutral Lipid Production by Human Sebocytes
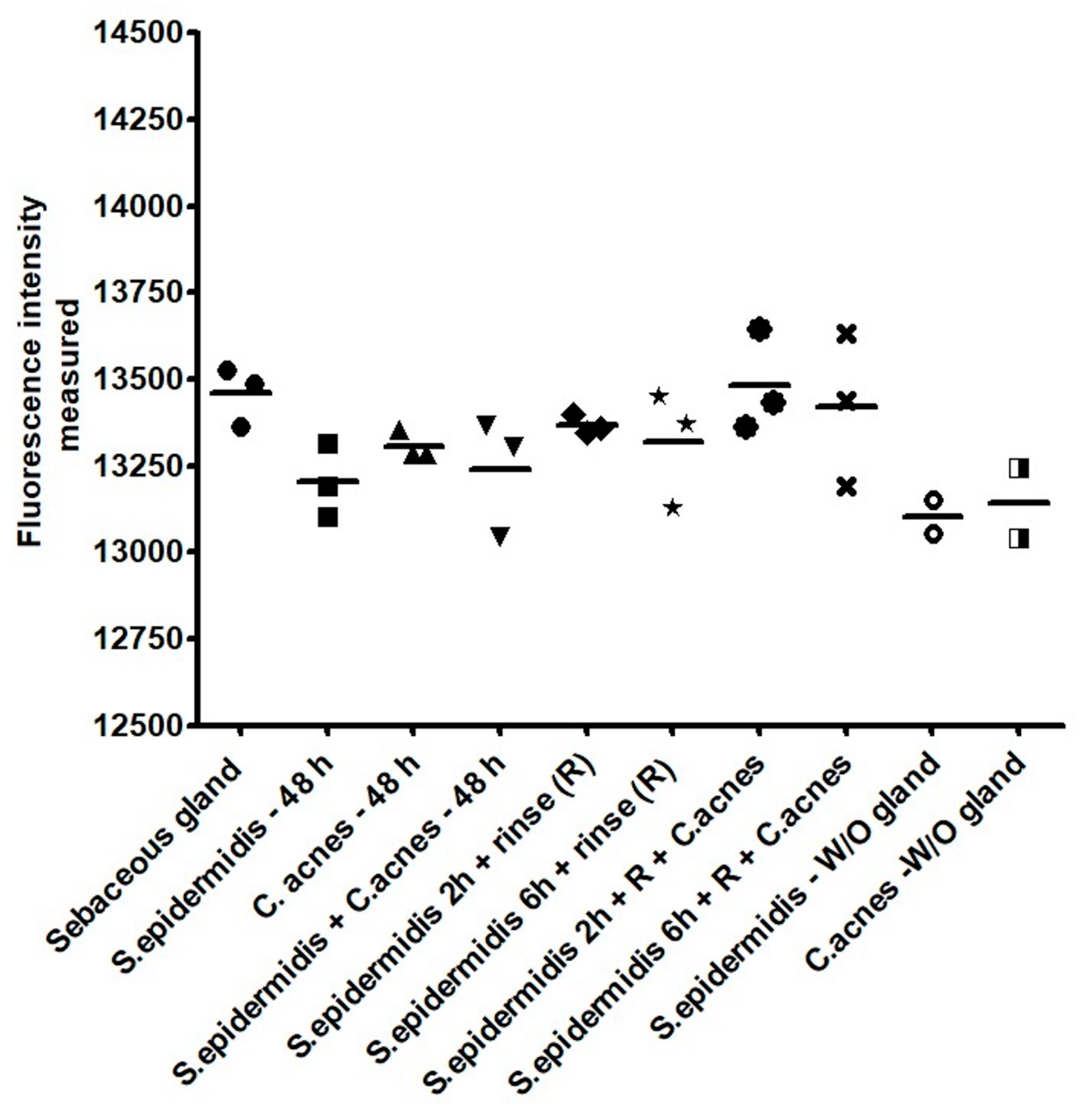
3.4. C. acnes and S. epidermidis Strains and Inoculation Ratio Can Be Optimized for 3D Sebaceous Gland Ex Vivo Culture
- C. acnes: 1.09 × 109;
- S. epidermidis: rinse at 2 h and C. acnes 1 × 105 and after rinse: 2.71 × 108;
- S. epidermidis: rinse at 6 h and C. acnes 1 × 105 and after rinse: 1.79 × 108.
3.5. C. acnes Bacterial Load Can Be Imaged via 3D Confocal Microscopy of Sebaceous Glands
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kealey, T.; Philpott, M.; Guy, R. The regulatory biology of the human pilosebaceous unit. Baillieres Clin. Obstet. Gynaecol. 1997, 11, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, M.; Flori, E.; Mastrofrancesco, A.; Briganti, S.; Lora, V.; Capitanio, B.; Zouboulis, C.C.; Picardo, M. Sebocyte differentiation as a new target for acne therapy: An in vivo experience. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1803–1814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-F.; Cai, X.-L.; Jing, K.-P.; Pi, X.-X.; Liao, P.-Y.; Li, S.-J.; Li, W.; Cai, C.-C.; Quan, J.-H.; Fan, Y.-M.; et al. Differentiation Model Establishment and Differentiation-Related Protein Screening in Primary Cultured Human Sebocytes. BioMed Res. Int. 2018, 2018, 7174561. [Google Scholar] [CrossRef] [PubMed]
- Philpott, M.P. Culture of the human pilosebaceous unit, hair follicle and sebaceous gland. Exp. Dermatol. 2018, 27, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, N.; Hernandez, L.E.; Martin, M.R.; Does, A.V.; Nouri, K. Acne treatment review and future perspectives. Dermatol. Ther. 2022, 35, e15719. [Google Scholar] [CrossRef]
- Platsidaki, E.; Dessinioti, C. Recent advances in understanding Propionibacterium acnes (Cutibacterium acnes) in acne. F1000Research 2018, 7, 1953. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- McLaughlin, J.; Watterson, S.; Layton, A.M.; Bjourson, A.J.; Barnard, E.; McDowell, A. Propionibacterium acnes and Acne Vulgaris: New Insights from the Integration of Population Genetic, Multi-Omic, Biochemical and Host-Microbe Studies. Microorganisms 2019, 7, 128. [Google Scholar] [CrossRef]
- de Bengy, A.; Forraz, N.; Danoux, L.; Berthelemy, N.; Cadau, S.; Degoul, O.; Andre, V.; Pain, S.; McGuckin, C. Development of new 3D human ex vivo models to study sebaceous gland lipid metabolism and modulations. Cell Prolif. 2018, 52, e12524. [Google Scholar] [CrossRef]
- Carmona-Cruz, S.; Orozco-Covarrubias, L.; Sáez-De-Ocariz, M. The Human Skin Microbiome in Selected Cutaneous Diseases. Front. Cell. Infect. Microbiol. 2022, 12, 834135. [Google Scholar] [CrossRef]
- Muñoz-Egea, M.-C.; García-Pedrazuela, M.; Mahillo, I.; García, M.J.; Esteban, J. Autofluorescence as a Tool for Structural Analysis of Biofilms Formed by Nonpigmented Rapidly Growing Mycobacteria. Appl. Environ. Microbiol. 2013, 79, 1065–1067. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pellicer, P.; Navarro-Moratalla, L.; Núñez-Delegido, E.; Ruzafa-Costas, B.; Agüera-Santos, J.; Navarro-López, V. Acne, Microbiome, and Probiotics: The Gut–Skin Axis. Microorganisms 2022, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Bagatin, E.; Freitas, T.H.P.; Rivitti-Machado, M.C.; Machado, M.C.R.; Ribeiro, B.M.; Nunes, S.; Rocha, M.A.D.D. Adult female acne: A guide to clinical practice. An Bras Dermatol. 2019, 94, 62–75, Erratum in: An Bras Dermatol. 2019, 94, 255. [Google Scholar] [CrossRef] [PubMed]
- Araviiskaia, E.; Estebaranz, J.L.L.; Pincelli, C. Dermocosmetics: Beneficial adjuncts in the treatment of acne vulgaris. J. Dermatol. Treat. 2019, 32, 3–10. [Google Scholar] [CrossRef]
- Conforti, C.; Giuffrida, R.; Fadda, S.; Fai, A.; Romita, P.; Zalaudek, I.; Dianzani, C. Topical dermocosmetics and acne vulgaris. Dermatol. Ther. 2020, 34, e14436. [Google Scholar] [CrossRef]
- Goodarzi, A.; Mozafarpoor, S.; Bodaghabadi, M.; Mohamadi, M. The potential of probiotics for treating acne vulgaris: A review of literature on acne and microbiota. Dermatol. Ther. 2020, 33, e13279. [Google Scholar] [CrossRef]
- Fitz-Gibbon, S.; Tomida, S.; Chiu, B.-H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Propionibacterium acnes Strain Populations in the Human Skin Microbiome Associated with Acne. J. Investig. Dermatol. 2013, 133, 2152–2160. [Google Scholar] [CrossRef]
- Barnard, E.; Shi, B.; Kang, D.; Craft, N.; Li, H. The balance of metagenomic elements shapes the skin microbiome in acne and health. Sci. Rep. 2016, 6, 39491. [Google Scholar] [CrossRef]
- Dréno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: A brief look at the latest updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32 (Suppl. S2), 5–14. [Google Scholar] [CrossRef]
- Ahle, C.M.; Feidenhansl, C.; Brüggemann, H. Cutibacterium acnes. Trends Microbiol. 2023, 31, 419–420. [Google Scholar] [CrossRef]
- Dréno, B.; Dagnelie, M.A.; Khammari, A.; Corvec, S. The Skin Microbiome: A New Actor in Inflammatory Acne. Am. J. Clin. Dermatol. 2020, 21 (Suppl. 1), 18–24. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, P.; Falahzadeh, K.; Nematizadeh, M.; Farazandeh, P.; Payab, M.; Larijani, B.; Beik, A.T.; Arjmand, B. Tissue Engineered Skin Substitutes. Adv. Exp. Med. Biol. 2018, 1107, 143–188. [Google Scholar] [CrossRef]
- Portugal-Cohen, M.; Cohen, D.; Kohen, R.; Oron, M. Exploitation of alternative skin models from academia to industry: Proposed functional categories to answer needs and regulation demands. Front. Physiol. 2023, 14, 1215266. [Google Scholar] [CrossRef] [PubMed]
- Podrini, C.; Schramm, L.; Marianantoni, G.; Apolinarska, J.; McGuckin, C.; Forraz, N.; Milet, C.; Desroches, A.-L.; Payen, P.; D’Aguanno, M.; et al. Topical Administration of Lactiplantibacillus plantarum (SkinDuoTM) Serum Improves Anti-Acne Properties. Microorganisms 2023, 11, 417. [Google Scholar] [CrossRef] [PubMed]
- Mayslich, C.; Grange, P.A.; Castela, M.; Marcelin, A.G.; Calvez, V.; Dupin, N. Characterization of a Cutibacterium acnes Camp Factor 1-Related Peptide as a New TLR-2 Modulator in In Vitro and Ex Vivo Models of Inflammation. Int. J. Mol. Sci. 2022, 23, 5065. [Google Scholar] [CrossRef] [PubMed]
- Laclaverie, M.; Rouaud-Tinguely, P.; Grimaldi, C.; Jugé, R.; Marchand, L.; Aymard, E.; Closs, B. Development and characterization of a 3D in vitro model mimicking acneic skin. Exp. Dermatol. 2020, 30, 347–357. [Google Scholar] [CrossRef]
- Nagy, I.; Pivarcsi, A.; Kis, K.; Koreck, A.; Bodai, L.; McDowell, A.; Seltmann, H.; Patrick, S.; Zouboulis, C.C.; Kemény, L. Propionibacterium acnes and lipopolysaccharide induce the expression of antimicrobial peptides and proinflammatory cytokines/chemokines in human sebocytes. Microbes Infect. 2006, 8, 2195–2205. [Google Scholar] [CrossRef]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut–Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Baglama, Š.Š.; Trčko, K. Skin and gut microbiota dysbiosis in autoimmune and inflammatory skin diseases. Acta Derm. Alp Pannonica Adriat. 2022, 31, 105–109. [Google Scholar]
- Kuehnast, T.; Cakar, F.; Weinhäupl, T.; Pilz, A.; Selak, S.; Schmidt, M.A.; Rüter, C.; Schild, S. Comparative analyses of biofilm formation among different Cutibacterium acnes isolates. Int. J. Med. Microbiol. 2018, 308, 1027–1035. [Google Scholar] [CrossRef]
- Mack, D.; Davies, A.P.; Harris, L.G.; Rohde, H.; Horstkotte, M.A.; Knobloch, J.K.-M. Microbial interactions in Staphylococcus epidermidis biofilms. Anal. Bioanal. Chem. 2006, 387, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, I.A.; Thipe, V.C.; Katti, K.V.; Mandiwana, V.; Kalombo, M.L.; Ray, S.S.; Rikhotso, R.; van Vuuren, A.J.; Esmear, T.; Lall, N. Targeting Acne Bacteria and Wound Healing In Vitro Using Plectranthus aliciae, Rosmarinic Acid, and Tetracycline Gold Nanoparticles. Pharmaceuticals 2022, 15, 933. [Google Scholar] [CrossRef] [PubMed]
- Taleb, M.H.; Abdeltawab, N.F.; Shamma, R.N.; Abdelgayed, S.S.; Mohamed, S.S.; Farag, M.A.; Ramadan, M.A. Origanum vulgare L. Essential Oil as a Potential Anti-Acne Topical Nanoemulsion-In Vitro and In Vivo Study. Molecules 2018, 23, 2164. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STRAIN | MEDIA | GROWTH | INCUBATIONS | CELL NUMBERS | COUNT POINT |
|---|---|---|---|---|---|
| Staphylococcus epidermidis CIP28.61 | TSA agar | Aerobic | 37 °C for 24/48 h * 5% CO2 | 1.0 × 107 | 2 h/6 h/24 h/48 h |
| Cutibacterium acnes DSM 1897-Type IA | COS agar | Anaerobic | 37 °C for 48/72 h * 5% CO2 | 1.0 × 107 | 2 h/6 h/24 h/48 h |
| Condition | TSA (UFC/mL) | Average TSA | COS (UFC/mL) | Average COS |
|---|---|---|---|---|
| S.epidermidis + C.acnes 1E5 | 3.20 × 105 | 1.87 × 108 | 5.50 × 105 | 4.37 × 108 |
| 1.90 × 108 | 6.20 × 108 | |||
| 3.0 × 108 | 6.90 × 108 | |||
| S.epidermidis + rinse at 2 h | 1.99 × 108 | 1.46 × 108 | ||
| 2.03 × 108 | ||||
| 3.70 × 107 | ||||
| S.epidermidis + rinse at 2 h + C.acnes 1E5 after rinse | 2.06 × 108 | 1.72 × 108 | 3.70 × 108 | 2.71 × 108 |
| 2.59 × 108 | 3.90 × 108 | |||
| 5.10 × 107 | 6.90 × 108 | |||
| S.epidermidis + rinse at 6 h | 5.90 × 108 | 4.57 × 108 | ||
| 5.40 × 108 | ||||
| 2.41 × 108 | ||||
| S.epidermidis + rinse at 6 h + C.acnes 1E5 after rinse | 2.30 × 108 | 7.76 × 107 | 5.30 × 108 | 1.79 × 108 |
| 1.10 × 106 | 3.50 × 106 | |||
| 1.78 × 106 | 2.60 × 106 | |||
| S.epidermidis 1E4 | 8.50 × 108 | 7.10 × 108 | ||
| 5.40 × 108 | ||||
| 7.40 × 108 | ||||
| C.acnes 1E5 | 1.30 × 109 | 1.09 × 109 | ||
| 1.38 × 109 | ||||
| 5.80 × 108 | ||||
| No IC | >3.00 × 104 | >3.00 × 104 | >3.00 × 104 | >3.00 × 104 |
| >3.00 × 104 | >3.00 × 104 | |||
| >3.00 × 104 | >3.00 × 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forraz, N.; Bize, C.; Desroches, A.-L.; Milet, C.; Payen, P.; Chanut, P.; Kern, C.; Garcia, C.; McGuckin, C. The World’s First Acne Dysbiosis-like Model of Human 3D Ex Vivo Sebaceous Gland Colonized with Cutibacterium acnes and Staphylococcus epidermidis. Microorganisms 2023, 11, 2183. https://doi.org/10.3390/microorganisms11092183
Forraz N, Bize C, Desroches A-L, Milet C, Payen P, Chanut P, Kern C, Garcia C, McGuckin C. The World’s First Acne Dysbiosis-like Model of Human 3D Ex Vivo Sebaceous Gland Colonized with Cutibacterium acnes and Staphylococcus epidermidis. Microorganisms. 2023; 11(9):2183. https://doi.org/10.3390/microorganisms11092183
Chicago/Turabian StyleForraz, Nico, Cécile Bize, Anne-Laure Desroches, Clément Milet, Pauline Payen, Pauline Chanut, Catherine Kern, Christine Garcia, and Colin McGuckin. 2023. "The World’s First Acne Dysbiosis-like Model of Human 3D Ex Vivo Sebaceous Gland Colonized with Cutibacterium acnes and Staphylococcus epidermidis" Microorganisms 11, no. 9: 2183. https://doi.org/10.3390/microorganisms11092183