
Global Risk Maps of Climate Change Impacts on the Distribution of Acinetobacter baumannii Using GIS

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
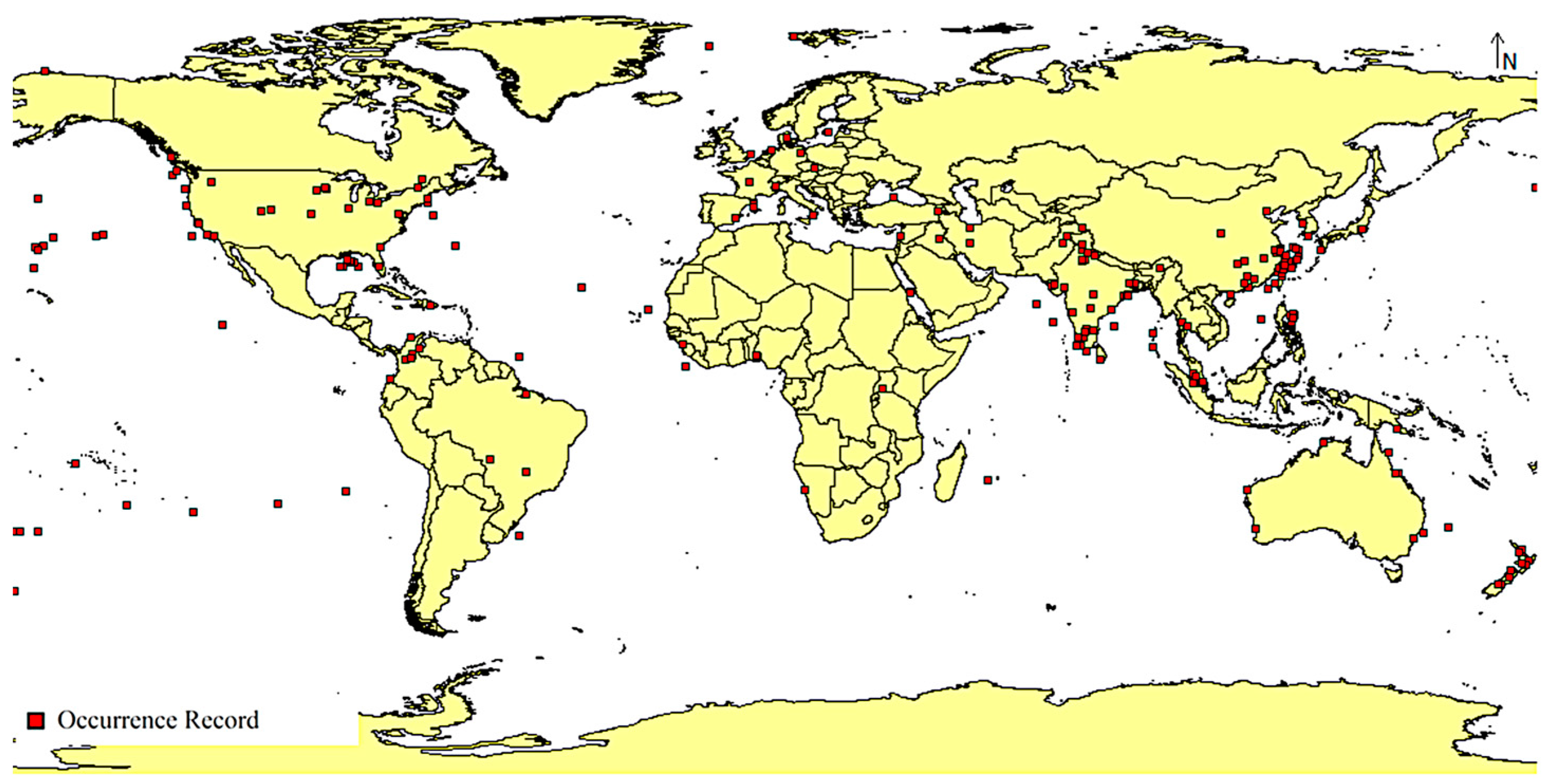
2.1.1. Species Occurrence Records
2.1.2. Climate Data
2.2. Model Construction
2.3. Quality Assessment
2.4. Visualization and Exporting Maps
3. Results
3.1. Model Quality
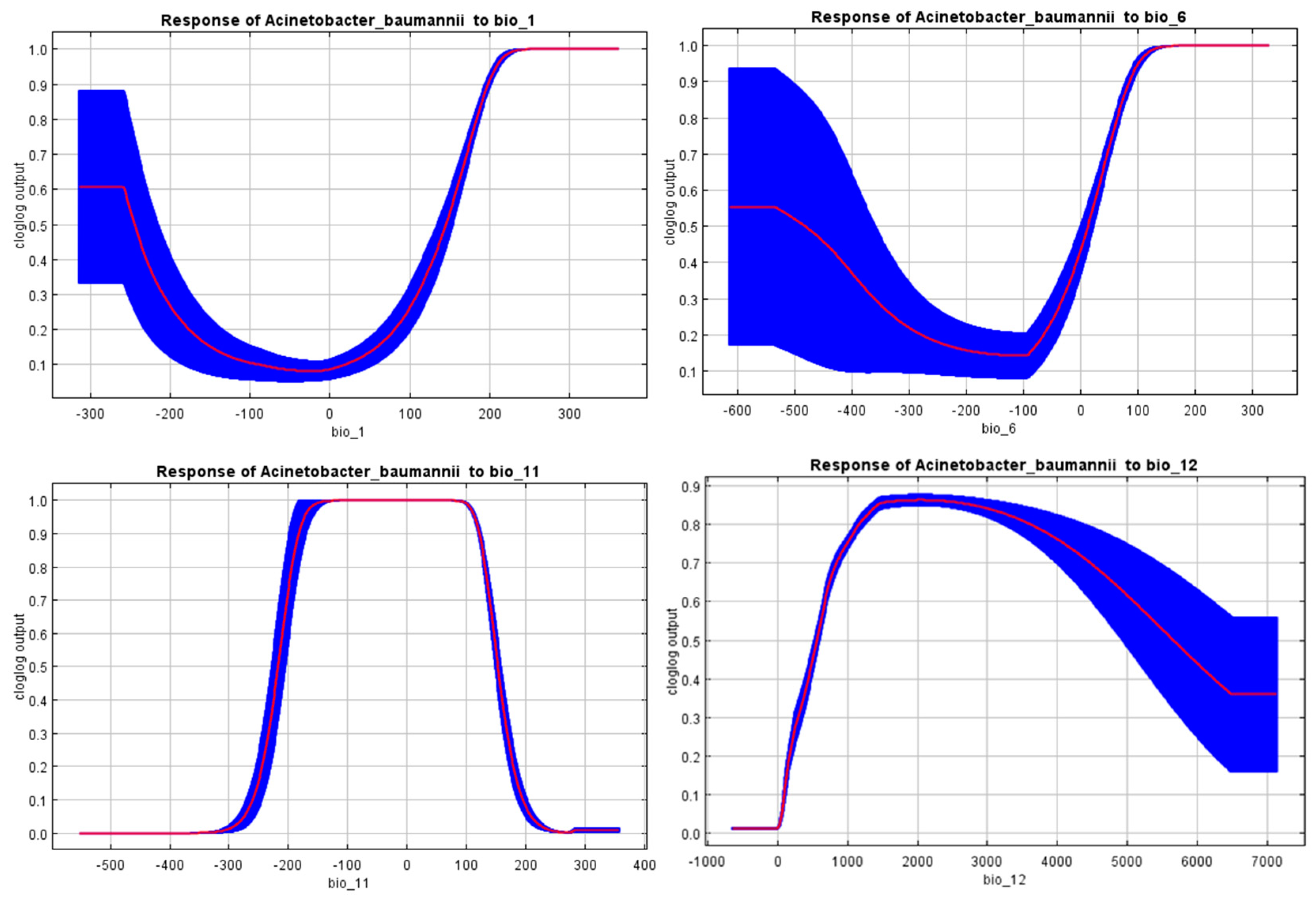
3.2. Significant Environmental Variables
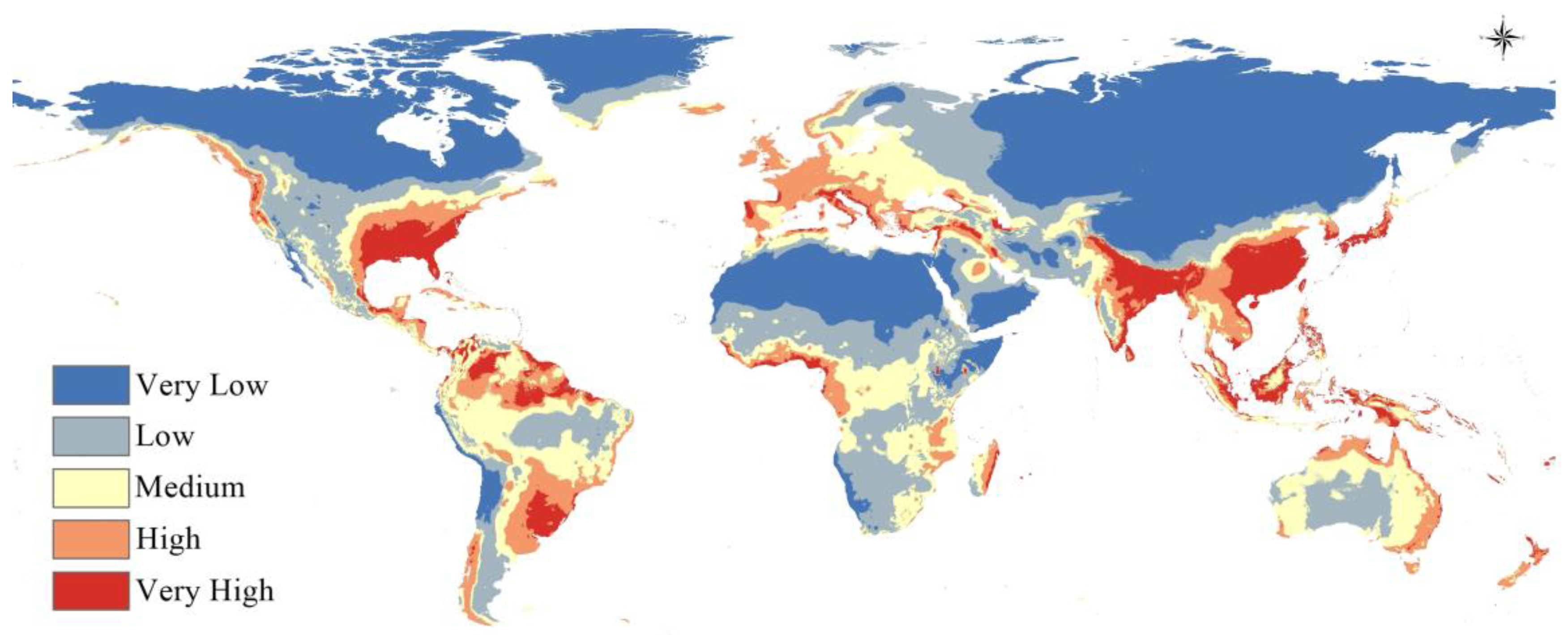
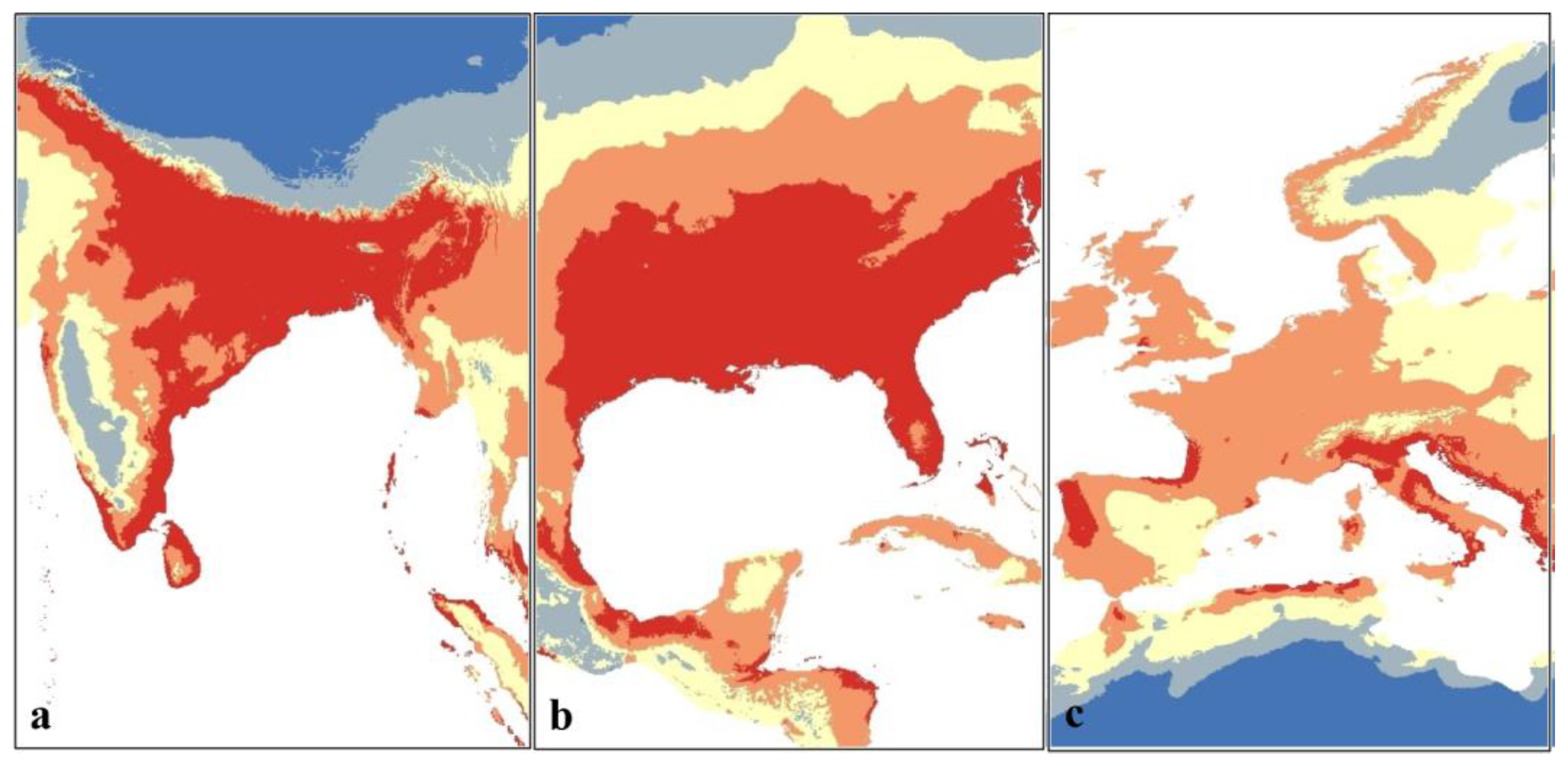
3.3. Potential Current Distribution
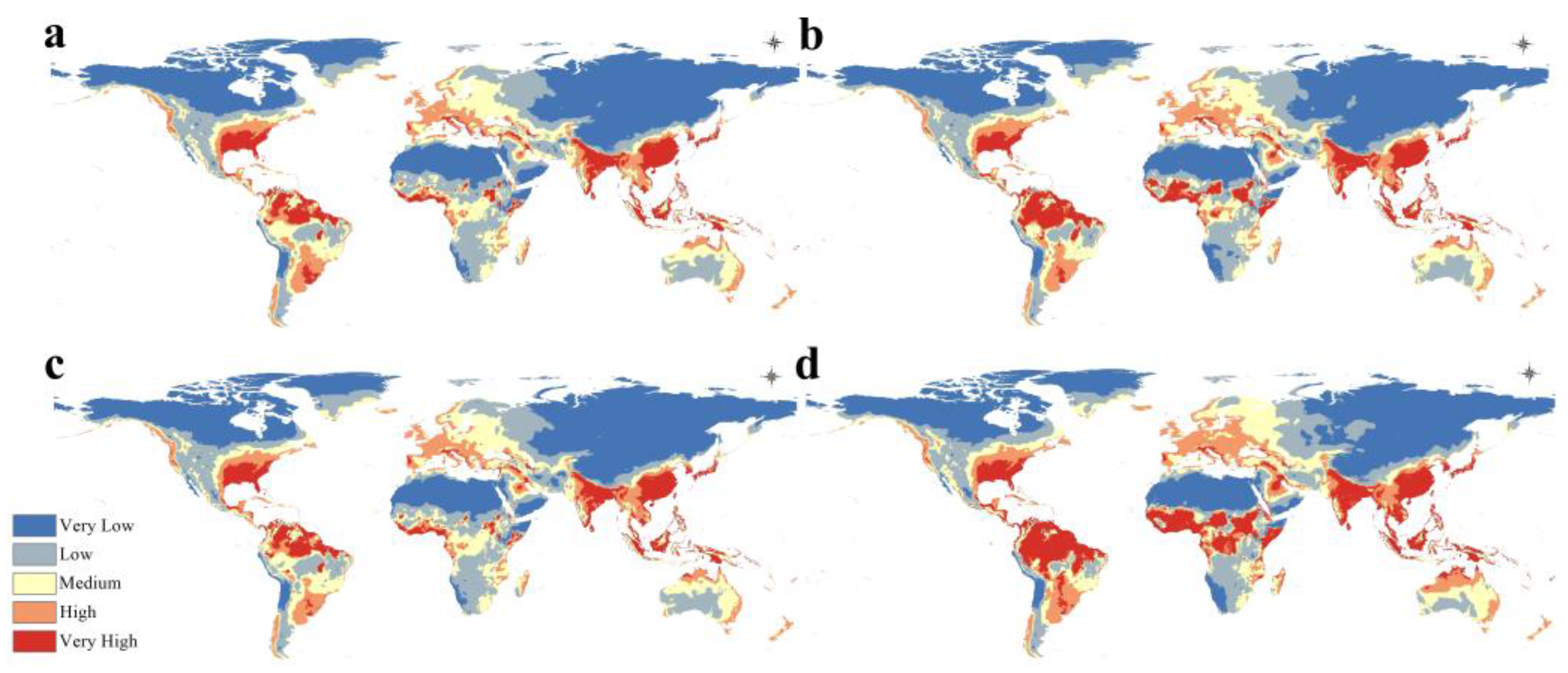
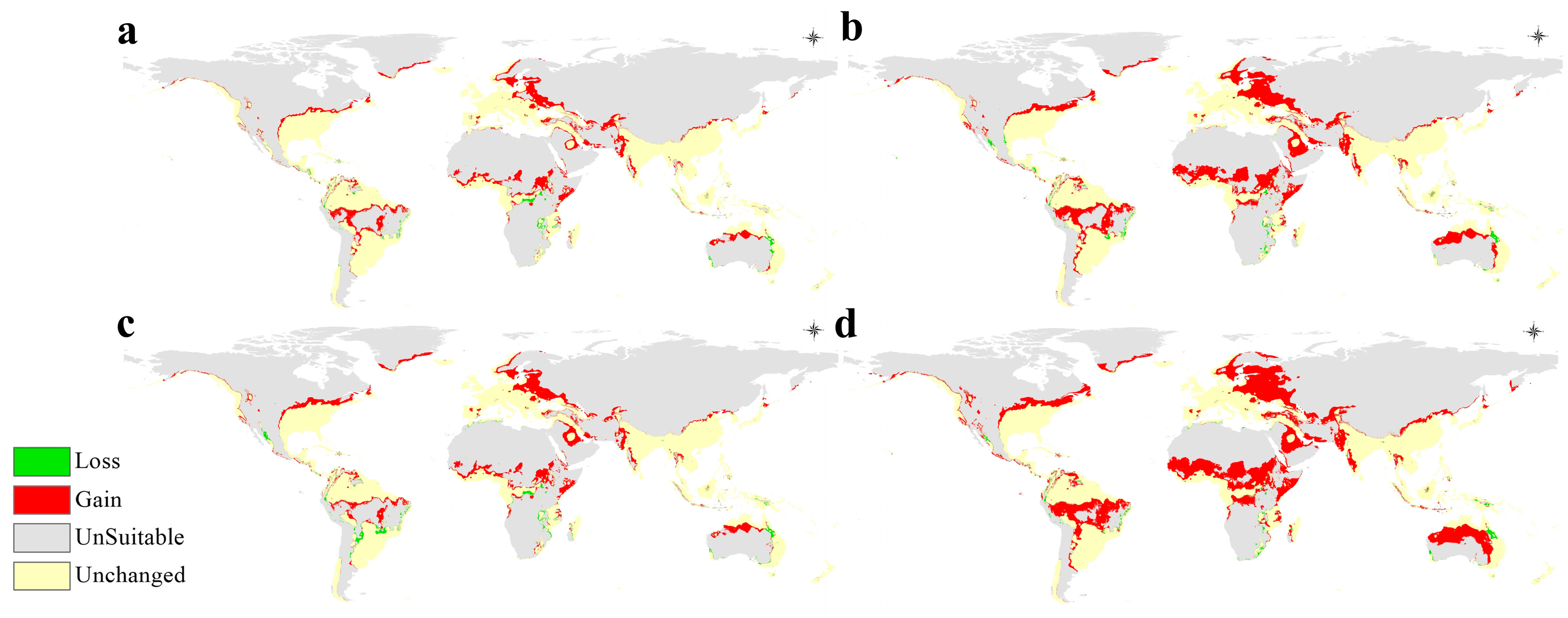
3.4. Potential Future Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hrenovic, J.; Durn, G.; Goic-Barisic, I.; Kovacic, A. Occurrence of an Environmental Acinetobacter baumannii Strain Similar to a Clinical Isolate in Paleosol from Croatia. Appl. Environ. Microbiol. 2014, 80, 2860–2866. [Google Scholar] [CrossRef]
- Bernards, A.T.; Harinck, H.I.J.; Dijkshoorn, L.; van der Reijden, T.J.K.; van den Broek, P.J. Persistent Acinetobacter baumannii? Look Inside Your Medical Equipment. Infect. Control Hosp. Epidemiol. 2004, 25, 1002–1004. [Google Scholar] [CrossRef]
- Garnacho-Montero, J.; Amaya-Villar, R. Multiresistant Acinetobacter baumannii Infections: Epidemiology and Management. Curr. Opin. Infect. Dis. 2010, 23, 332–339. [Google Scholar] [CrossRef]
- Jang, T.-N.; Lee, S.-H.; Huang, C.-H.; Lee, C.-L.; Chen, W.-Y. Risk Factors and Impact of Nosocomial Acinetobacter baumannii Bloodstream Infections in the Adult Intensive Care Unit: A Case–Control Study. J. Hosp. Infect. 2009, 73, 143–150. [Google Scholar] [CrossRef]
- Evans, B.A.; Hamouda, A.; Amyes, S.G.B. The Rise of Carbapenem-Resistant Acinetobacter baumannii. Curr. Pharm. Des. 2013, 19, 223–238. [Google Scholar] [CrossRef]
- Fournier, P.E.; Richet, H.; Weinstein, R.A. The Epidemiology and Control of Acinetobacter baumannii in Health Care Facilities. Clin. Infect. Dis. 2006, 42, 692–699. [Google Scholar] [CrossRef]
- Berndtson, A.E. Increasing Globalization and the Movement of Antimicrobial Resistance between Countries. Surg. Infect. 2020, 21, 579–585. [Google Scholar] [CrossRef]
- Durante-Mangoni, E.; Zarrilli, R. Global Spread of Drug-Resistant Acinetobacter baumannii: Molecular Epidemiology and Management of Antimicrobial Resistance. Future Microbiol. 2011, 6, 407–422. [Google Scholar] [CrossRef]
- Rangel, K.; Chagas, T.P.G.; De-Simone, S.G. Acinetobacter baumannii Infections in Times of COVID-19 Pandemic. Pathogens 2021, 10, 1006. [Google Scholar] [CrossRef]
- Russo, A.; Gavaruzzi, F.; Ceccarelli, G.; Borrazzo, C.; Oliva, A.; Alessandri, F.; Magnanimi, E.; Pugliese, F.; Venditti, M. Multidrug-Resistant Acinetobacter baumannii Infections in COVID-19 Patients Hospitalized in Intensive Care Unit. Infection 2022, 50, 83–92. [Google Scholar] [CrossRef]
- Vázquez-López, R.; Solano-Gálvez, S.G.; Juárez Vignon-Whaley, J.J.; Abello Vaamonde, J.A.; Padró Alonzo, L.A.; Rivera Reséndiz, A.; Muleiro Álvarez, M.; Vega López, E.N.; Franyuti-Kelly, G.; Álvarez-Hernández, D.A.; et al. Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics 2020, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, G.M.; Maragakis, L.L.; Perl, T.M. Acinetobacter baumannii: Epidemiology, Antimicrobial Resistance, and Treatment Options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef]
- Choi, J.Y.; Park, Y.S.; Kim, C.O.; Park, Y.S.; Yoon, H.J.; Shin, S.Y.; Kim, Y.-A.; Song, Y.G.; Yong, D.; Lee, K.; et al. Mortality Risk Factors of Acinetobacter baumannii Bacteraemia. Intern. Med. J. 2005, 35, 599–603. [Google Scholar] [CrossRef]
- Falagas, M.E.; Bliziotis, I.A.; Siempos, I.I. Attributable Mortality of Acinetobacter baumannii Infections in Critically Ill Patients: A Systematic Review of Matched Cohort and Case-Control Studies. Crit. Care 2006, 10, R48. [Google Scholar] [CrossRef]
- Serota, D.P.; Sexton, M.E.; Kraft, C.S.; Palacio, F. Severe Community-Acquired Pneumonia Due to Acinetobacter baumannii in North America: Case Report and Review of the Literature. Open Forum Infect. Dis. 2018, 5, ofy044. [Google Scholar] [CrossRef]
- Adamo, N.; Al-Ansari, N.; Sissakian, V. Review of Climate Change Impacts on Human Environment: Past, Present and Future Projections. Engineering 2021, 13, 605–630. [Google Scholar] [CrossRef]
- Wuebbles, D.J.; Jain, A.K. Concerns about Climate Change and the Role of Fossil Fuel Use. Fuel Process. Technol. 2001, 71, 99–119. [Google Scholar] [CrossRef]
- Dale, V.H. The Relationship between Land-Use Change and Climate Change. Ecol. Appl. 1997, 7, 753–769. Available online: https://esajournals.onlinelibrary.wiley.com/doi/full/10.1890/1051-0761%281997%29007%5B0753%3ATRBLUC%5D2.0.CO%3B2 (accessed on 16 July 2023). [CrossRef]
- Du, M.; Peng, C.; Wang, X.; Chen, H.; Wang, M.; Zhu, Q. Quantification of Methane Emissions from Municipal Solid Waste Landfills in China during the Past Decade. Renew. Sustain. Energy Rev. 2017, 78, 272–279. [Google Scholar] [CrossRef]
- Dore, M.H.I. Climate Change and Changes in Global Precipitation Patterns: What Do We Know? Environ. Int. 2005, 31, 1167–1181. [Google Scholar] [CrossRef]
- Hashim, J.H.; Hashim, Z. Climate Change, Extreme Weather Events, and Human Health Implications in the Asia Pacific Region. Asia Pac. J. Public Health 2016, 28 (Suppl. 2), 8S–14S. [Google Scholar] [CrossRef]
- Berlemann, M.; Steinhardt, M.F. Climate Change, Natural Disasters, and Migration—A Survey of the Empirical Evidence. CESifo Econ. Stud. 2017, 63, 353–385. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Chown, S.L.; Hoffmann, A.A.; Kristensen, T.N.; Angilletta, M., Jr.; Stenseth, N.C.; Pertoldi, C. Adapting to Climate Change: A Perspective from Evolutionary Physiology. Clim. Res. 2010, 43, 3–15. [Google Scholar] [CrossRef]
- HilleRisLambers, J.; Harsch, M.A.; Ettinger, A.K.; Ford, K.R.; Theobald, E.J. How Will Biotic Interactions Influence Climate Change–Induced Range Shifts? Ann. N. Y. Acad. Sci. 2013, 1297, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Moran, E.V.; Alexander, J.M. Evolutionary Responses to Global Change: Lessons from Invasive Species. Ecol. Lett. 2014, 17, 637–649. [Google Scholar] [CrossRef]
- Batten, S. Climate Change and the Macro-Economy: A Critical Review; Bank of England: London, UK, 2018. [Google Scholar] [CrossRef]
- Chang, K.-T. Geographic Information System. In International Encyclopedia of Geography; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 1–9. [Google Scholar] [CrossRef]
- Peuquet, D.J.; Marble, D.F. (Eds.) Introductory Readings. In Geographic Information Systems; CRC Press: London, UK, 2001. [Google Scholar] [CrossRef]
- Hinton, J.C. GIS and Remote Sensing Integration for Environmental Applications. Int. J. Geogr. Inf. Syst. 1996, 10, 877–890. [Google Scholar] [CrossRef]
- Kerr, J.T.; Ostrovsky, M. From Space to Species: Ecological Applications for Remote Sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Foody, G.M. GIS: Biodiversity Applications. Prog. Phys. Geogr. Earth Environ. 2008, 32, 223–235. [Google Scholar] [CrossRef]
- Salem, B.B. Application of GIS to Biodiversity Monitoring. J. Arid Environ. 2003, 54, 91–114. [Google Scholar] [CrossRef]
- Samanta, S.; Pal, D.K.; Lohar, D.; Pal, B. Interpolation of Climate Variables and Temperature Modeling. Theor. Appl. Climatol. 2012, 107, 35–45. [Google Scholar] [CrossRef]
- Weart, S. The Development of General Circulation Models of Climate. Stud. Hist. Philos. Sci. Part B Stud. Hist. Philos. Mod. Phys. 2010, 41, 208–217. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive Habitat Distribution Models in Ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species Distribution Models (SDM): Applications, Benefits and Challenges in Invasive Species Management. CABI Rev. 2019, 2019, 1–13. [Google Scholar] [CrossRef]
- GBIF.Org User. Occurrence Download; GBIF: Copenhagen, Denmark, 2023. [Google Scholar] [CrossRef]
- Telenius, A. Biodiversity Information Goes Public: GBIF at Your Service. Nord. J. Bot. 2011, 29, 378–381. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very High Resolution Interpolated Climate Surfaces for Global Land Areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Yukimoto, S.; Adachi, Y.; Hosaka, M.; Sakami, T.; Yoshimura, H.; Hirabara, M.; Tanaka, T.Y.; Shindo, E.; Tsujino, H.; Deushi, M.; et al. A New Global Climate Model of the Meteorological Research Institute: MRI-CGCM3—Model Description and Basic Performance. J. Meteorol. Soc. Jpn. Ser II 2012, 90, 23–64. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The Representative Concentration Pathways: An Overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Booth, T.H. Checking Bioclimatic Variables That Combine Temperature and Precipitation Data before Their Use in Species Distribution Models. Austral Ecol. 2022, 47, 1506–1514. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of Maximum Entropy Modeling in Wildlife Research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Ahmadi, M. Assessing Accuracy Methods of Species Distribution Models: AUC, Specificity, Sensitivity and the True Skill Statistic. Glob. J. Hum.-Soc. Sci. B Geogr. Geo-Sci. Environ. Sci. Disaster Manag. 2018, 18, 1–13. [Google Scholar]
- Lissovsky, A.A.; Dudov, S.V. Species-Distribution Modeling: Advantages and Limitations of Its Application. 2. MaxEnt. Biol. Bull. Rev. 2021, 11, 265–275. [Google Scholar] [CrossRef]
- Liu, L.; Guan, L.; Zhao, H.; Huang, Y.; Mou, Q.; Liu, K.; Chen, T.; Wang, X.; Zhang, Y.; Wei, B.; et al. Modeling Habitat Suitability of Houttuynia cordata thunb (Ceercao) Using MaxEnt under Climate Change in China. Ecol. Inform. 2021, 63, 101324. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the Potential Distribution of Paeonia veitchii (Paeoniaceae) in China by Incorporating Climate Change into a Maxent Model. Forests 2019, 10, 190. [Google Scholar] [CrossRef]
- Smith, A.B.; Santos, M.J. Testing the Ability of Species Distribution Models to Infer Variable Importance. Ecography 2020, 43, 1801–1813. [Google Scholar] [CrossRef]
- Inweregbu, K.; Dave, J.; Pittard, A. Nosocomial Infections. Contin. Educ. Anaesth. Crit. Care Pain 2005, 5, 14–17. [Google Scholar] [CrossRef]
- Chim, H.; Tan, B.H.; Song, C. Five-Year Review of Infections in a Burn Intensive Care Unit: High Incidence of Acinetobacter baumannii in a Tropical Climate. Burns 2007, 33, 1008–1014. [Google Scholar] [CrossRef]
- Noel, H.R.; Petrey, J.R.; Palmer, L.D. Mobile Genetic Elements in Acinetobacter Antibiotic-Resistance Acquisition and Dissemination. Ann. N. Y. Acad. Sci. 2022, 1518, 166–182. [Google Scholar] [CrossRef]
- Joly-Guillou, M.-L. Clinical Impact and Pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Kumar, P.; Ciraj, A.M.; Shivananda, P.G. Acinetobacter baumannii—An Emerging Nosocomial Pathogen in the Burns Unit Manipal, India. Burns 2001, 27, 140–144. [Google Scholar] [CrossRef]
- Rodríguez, C.H.; Balderrama Yarhui, N.; Nastro, M.; Nuñez Quezada, T.; Castro Cañarte, G.; Magne Ventura, R.; Ugarte Cuba, T.; Valenzuela, N.; Roach, F.; Mota, M.I.; et al. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter baumannii in South America. J. Med. Microbiol. 2016, 65, 1088–1091. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Paulus, T.; Lugenheim, M.; Stefanik, D.; Higgins, P.G.; Edmond, M.B.; Wenzel, R.P.; Seifert, H. Nosocomial Bloodstream Infections Due to Acinetobacter baumannii, Acinetobacter Pittii and Acinetobacter Nosocomialis in the United States. J. Infect. 2012, 64, 282–290. [Google Scholar] [CrossRef]
- Ayobami, O.; Willrich, N.; Harder, T.; Okeke, I.N.; Eckmanns, T.; Markwart, R. The Incidence and Prevalence of Hospital-Acquired (Carbapenem-Resistant) Acinetobacter baumannii in Europe, Eastern Mediterranean and Africa: A Systematic Review and Meta-Analysis. Emerg. Microbes Infect. 2019, 8, 1747–1759. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; McClean, S. Mapping Global Prevalence of Acinetobacter baumannii and Recent Vaccine Development to Tackle It. Vaccines 2021, 9, 570. [Google Scholar] [CrossRef]
- Segagni Lusignani, L.; Starzengruber, P.; Dosch, V.; Assadian, O.; Presterl, E.; Diab-Elschahawi, M. Molecular Epidemiology of Multidrug-Resistant Clinical Isolates of Acinetobacter baumannii. Wien. Klin. Wochenschr. 2017, 129, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Revathi, G.; Siu, L.K.; Lu, P.-L.; Huang, L.-Y. First Report of NDM-1-Producing Acinetobacter baumannii in East Africa. Int. J. Infect. Dis. 2013, 17, e1255–e1258. [Google Scholar] [CrossRef]
- Gellings, P.S.; Wilkins, A.A.; Morici, L.A. Recent Advances in the Pursuit of an Effective Acinetobacter baumannii Vaccine. Pathogens 2020, 9, 1066. [Google Scholar] [CrossRef]
- McConnell, M.J.; Pachón, J. Active and Passive Immunization against Acinetobacter baumannii Using an Inactivated Whole Cell Vaccine. Vaccine 2010, 29, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.C.; Lahiri, C. Promising Acinetobacter baumannii Vaccine Candidates and Drug Targets in Recent Years. Front. Immunol. 2022, 13, 900509. [Google Scholar] [CrossRef] [PubMed]
- Dekic, S.; Hrenovic, J.; Durn, G.; Venter, C. Survival of Extensively- and Pandrug-Resistant Isolates of Acinetobacter baumannii in Soils. Appl. Soil Ecol. 2020, 147, 103396. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Percent Contribution | Permutation Importance |
|---|---|---|
| Annual mean temperature (bio_1) | 9.5% | 26.1% |
| Minimum temperature of the coldest month (bio_6) | 12.6% | 26.1% |
| Mean temperature of the coldest quarter (bio_11) | 37.8% | 32.9% |
| Annual precipitation (bio_12) | 40.2% | 14.9% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabour, A. Global Risk Maps of Climate Change Impacts on the Distribution of Acinetobacter baumannii Using GIS. Microorganisms 2023, 11, 2174. https://doi.org/10.3390/microorganisms11092174
Sabour A. Global Risk Maps of Climate Change Impacts on the Distribution of Acinetobacter baumannii Using GIS. Microorganisms. 2023; 11(9):2174. https://doi.org/10.3390/microorganisms11092174
Chicago/Turabian StyleSabour, Amal. 2023. "Global Risk Maps of Climate Change Impacts on the Distribution of Acinetobacter baumannii Using GIS" Microorganisms 11, no. 9: 2174. https://doi.org/10.3390/microorganisms11092174