
Hetero-Pathogenic O181:H4 EAHEC Strain of Sequence Type ST678 Associated with Hemolytic–Uremic Syndrome in Schoolchildren in Russia
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Strains
2.3. Electron Microscopy
2.4. Biochemical Properties
2.5. Mobility, Hemolytic Activity, and Colicin Production
2.6. Biofilm Formation and Susceptibility to Antimicrobials
2.7. Detection of Shiga Toxin 2 Production
2.8. Vero Cell Assay
3. Results
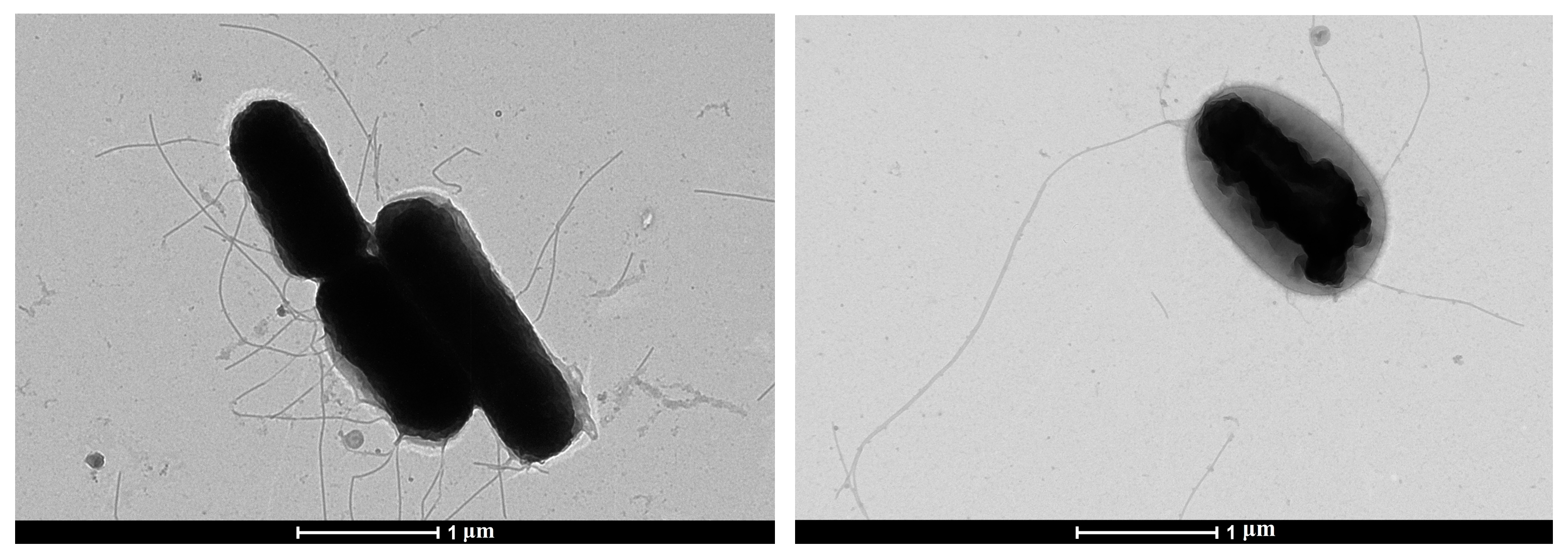
3.1. Electron Microscopy and Cultural Properties
3.2. Biochemical Properties
3.3. Hemolytic and Colicinogenic Activity
3.4. Biofilm Formation and Susceptibility to Antimicrobials
3.5. Detection of Shiga Toxin 2 Production
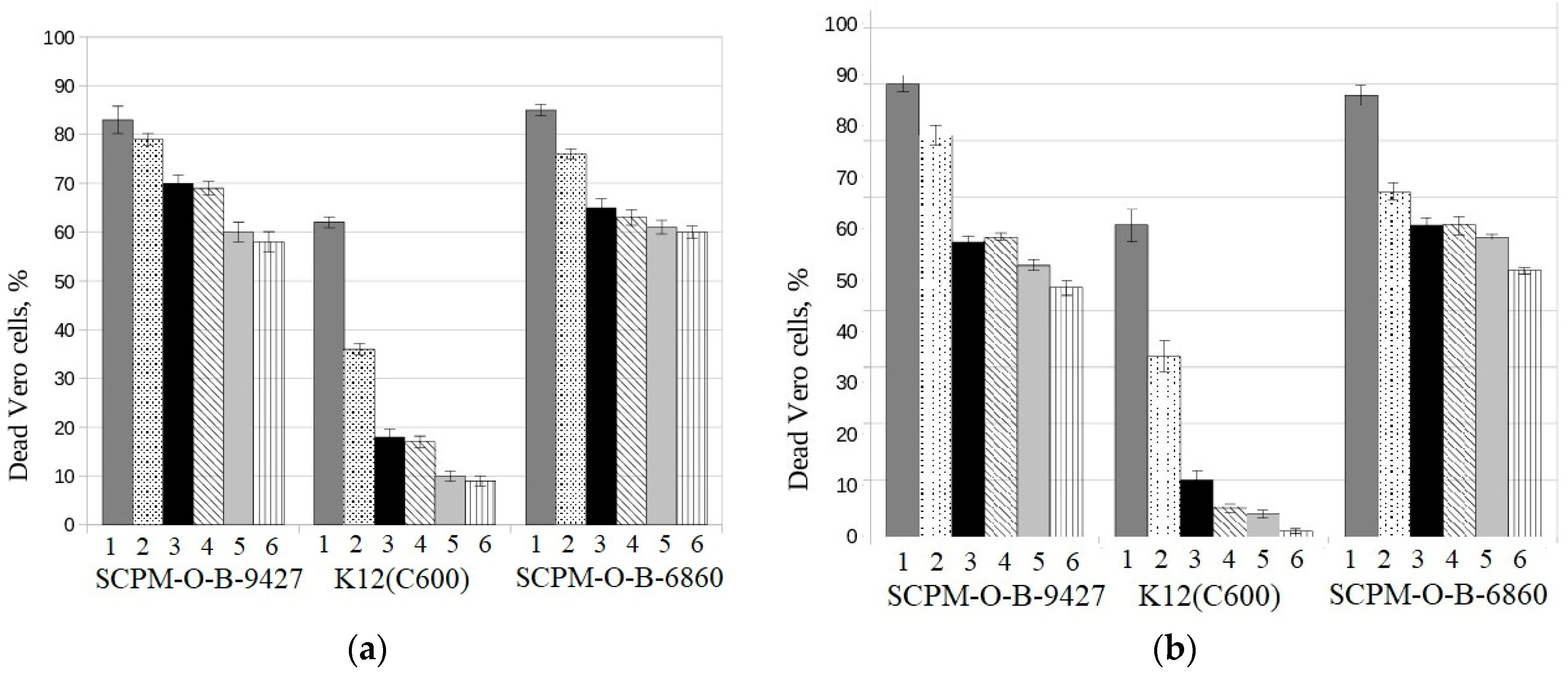
3.6. Cytotoxicity of E. coli Strain SCPM-O-B-9427
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santos, A.C.M.; Santos, F.F.; Silva, R.M.; Gomes, T.A.T. Diversity of Hybrid- and Hetero-Pathogenic Escherichia coli and Their Potential Implication in More Severe Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 339. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.A.; Elias, W.P.; Scaletsky, I.C.; Guth, B.E.; Rodrigues, J.F.; Piazza, R.M.; Ferreira, L.C.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47, 3–30. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Medina, M.; Mora, A.; Blanco, M.; López, C.; Alonso, M.P.; Bonacorsi, S.; Nicolas-Chanoine, M.H.; Darfeuille-Michaud, A.; Garcia-Gil, J.; Blanco, J. Similarity and divergence among adherent-invasive Escherichia coli and extraintestinal pathogenic E. coli strains. J. Clin. Microbiol. 2009, 47, 3968–3979. [Google Scholar] [CrossRef] [Green Version]
- Morabito, S.; Karch, H.; Mariani-Kurkdjian, P.; Schmidt, H.; Minelli, F.; Bingen, E.; Caprioli, A. Enteroaggregative, Shiga toxin-producing Escherichia coli O111:H2 associated with an outbreak of hemolytic-uremic syndrome. J. Clin. Microbiol. 1998, 36, 840–842. [Google Scholar] [CrossRef] [Green Version]
- Buchholz, U.; Bernard, H.; Werber, D.; Böhmer, M.M.; Remschmidt, C.; Wilking, H.; Deleré, Y.; an der Heiden, M.; Adlhoch, C.; Dreesman, J.; et al. German outbreak of Escherichia coli O104:H4 associated with sprouts. N. Engl. J. Med. 2011, 365, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Frank, C.; Werber, D.; Cramer, J.P.; Askar, M.; Faber, M.; an der Heiden, M.; Bernard, H.; Fruth, A.; Prager, R.; Spode, A.; et al. Epidemic profile of Shiga-toxin-producing Escherichia coli O104:H4 outbreak in Germany. N. Engl. J. Med. 2011, 365, 1771–1780. [Google Scholar] [CrossRef] [Green Version]
- Mellmann, A.; Harmsen, D.; Cummings, C.A.; Zentz, E.B.; Leopold, S.R.; Rico, A.; Prior, K.; Szczepanowski, R.; Ji, Y.; Zhang, W.; et al. Prospective genomic characterization of the German enterohemorrhagic Escherichia coli O104:H4 outbreak by rapid next generation sequencing technology. PLoS ONE 2011, 6, e22751. [Google Scholar] [CrossRef]
- Rasko, D.A.; Webster, D.R.; Sahl, J.W.; Bashir, A.; Boisen, N.; Scheutz, F.; Paxinos, E.E.; Sebra, R.; Chin, C.S.; Iliopoulos, D.; et al. Origins of the E. coli strain causing an outbreak of hemolytic-uremic syndrome in Germany. N. Engl. J. Med. 2011, 365, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chemother. 2012, 56, 3277–3282. [Google Scholar] [CrossRef] [Green Version]
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Köck, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyholm, O.; Heinikainen, S.; Pelkonen, S.; Hallanvuo, S.; Haukka, K.; Siitonen, A. Hetero-pathogenics of Shigatoxigenic and Enterotoxigenic Escherichia coli (STEC/ETEC) Among Human and Animal Isolates in Finland. Zoonoses Public Health 2015, 62, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.R.; Mammel, M.K.; Rasko, D.A.; Lacher, D.W. Hetero-pathogenic Shiga Toxin-Producing and Enterotoxigenic Escherichia sp. Cryptic Lineage 1 Strain 7v Harbors a Hetero-pathogenic Plasmid. Appl. Environ. Microbiol. 2016, 82, 4309–4319. [Google Scholar] [CrossRef] [Green Version]
- Prager, R.; Fruth, A.; Busch, U.; Tietze, E. Comparative analysis of virulence genes, genetic diversity, and phylogeny of Shiga toxin 2g and heat-stable enterotoxin STIa encoding Escherichia coli isolates from humans, animals, and environmental sources. Int. J. Med. Microbiol. 2011, 301, 181–191. [Google Scholar] [CrossRef]
- Michelacci, V.; Maugliani, A.; Tozzoli, R.; Corteselli, G.; Chiani, P.; Minelli, F.; Gigliucci, F.; Arancia, S.; Conedera, G.; Targhetta, C.; et al. Characterization of a novel plasmid encoding F4-like fimbriae present in a Shiga-toxin producing enterotoxigenic Escherichia coli isolated during the investigation on a case of hemolytic-uremic syndrome. Int. J. Med. Microbiol. 2018, 308, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Brilhante, M.; Perreten, V.; Donà, V. Multidrug resistance and multivirulence plasmids in enterotoxigenic and hetero-pathogenic Shiga toxin-producing/enterotoxigenic Escherichia coli isolated from diarrheic pigs in Switzerland. Vet. J. 2019, 244, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Zhang, J.; Ambikan, A.; Jernberg, C.; Ehricht, R.; Scheutz, F.; Xiong, Y.; Matussek, A. Molecular Characterization and Comparative Genomics of Clinical Hetero-pathogenic Shiga Toxin-Producing and Enterotoxigenic Escherichia coli (STEC/ETEC) Strains in Sweden. Sci. Rep. 2019, 9, 5619. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [Green Version]
- Gioia-Di Chiacchio, R.M.; Cunha, M.P.V.; de Sá, L.R.M.; Davies, Y.M.; Pereira, C.B.P.; Martins, F.H.; Munhoz, D.D.; Abe, C.M.; Franzolin, M.R.; Dos Santos, L.F.; et al. Novel Hetero-pathogenic of Typical Enteropathogenic Escherichia coli and Shiga-Toxin-Producing E. coli (tEPEC/STEC) Emerging From Pet Birds. Front. Microbiol. 2018, 9, 2975. [Google Scholar] [CrossRef] [Green Version]
- van Hoek, A.H.A.M.; van Veldhuizen, J.N.J.; Friesema, I.; Coipan, C.; Rossen, J.W.A.; Bergval, I.L.; Franz, E. Comparative genomics reveals a lack of evidence for pigeons as a main source of stx2f-carrying Escherichia coli causing disease in humans and the common existence of hetero-pathogenic Shiga toxin-producing and enteropathogenic E. coli pathotypes. BMC Genom. 2019, 20, 271. [Google Scholar] [CrossRef] [Green Version]
- Prager, R.; Lang, C.; Aurass, P.; Fruth, A.; Tietze, E.; Flieger, A. Two novel EHEC/EAEC hetero-pathogenic strains isolated from human infections. PLoS ONE 2014, 9, e95379. [Google Scholar] [CrossRef] [PubMed]
- Brzuszkiewicz, E.; Thürmer, A.; Schuldes, J.; Leimbach, A.; Liesegang, H.; Meyer, F.D.; Boelter, J.; Petersen, H.; Gottschalk, G.; Daniel, R. Genome sequence analyses of two isolates from the recent Escherichia coli outbreak in Germany reveal the emergence of a new pathotype: Entero-Aggregative-Haemorrhagic Escherichia coli (EAHEC). Arch. Microbiol. 2011, 193, 883–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallman, T.; Smith, G.P.; O’Brien, B.; Chattaway, M.A.; Finlay, D.; Grant, K.A.; Jenkins, C. Characterization of a verocytotoxin-producing enteroaggregative Escherichia coli serogroup O111:H21 strain associated with a household outbreak in Northern Ireland. J. Clin. Microbiol. 2012, 50, 4116–4119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chokoshvili, O.; Lomashvili, K.; Malakmadze, N.; Geleishvil, M.; Brant, J.; Imnadze, P.; Chitadze, N.; Tevzadze, L.; Chanturia, G.; Tevdoradze, T.; et al. Investigation of an outbreak of bloody diarrhea complicated with hemolytic uremic syndrome. J. Epidemiol. Glob. Health 2014, 4, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setyarini, W.; Raharjo, D.; Arizandy, R.Y.; Pamoengkas, Z.; Sudarmo, S.M.; Athiyyah, A.F.; Shirakawa, T. Molecular detection of a new pathotype enteroaggregative haemorrhagic Escherichia coli (EAHEC) in Indonesia, 2015. Infect. Dis. Rep. 2020, 12, 8745. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Pazhani, G.P.; Nataro, J.P.; Ramamurthy, T. Heterogenic virulence in a diarrheagenic Escherichia coli: Evidence for an EPEC expressing heat-labile toxin of ETEC. Int. J. Med. Microbiol. 2015, 305, 47–54. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Schiller, R.; Lammers, L.; Bauwens, A.; Fruth, A.; Middendorf, B.; Schmidt, M.A.; Tarr, P.I.; Dobrindt, U.; Karch, H.; et al. Heteropathogenic virulence and phylogeny reveal phased pathogenic metamorphosis in Escherichia coli O2:H6. EMBO Mol. Med. 2014, 6, 347–357. [Google Scholar] [CrossRef]
- Mariani-Kurkdjian, P.; Lemaître, C.; Bidet, P.; Perez, D.; Boggini, L.; Kwon, T.; Bonacorsi, S. Haemolytic-uraemic syndrome with bacteraemia caused by a new hetero-pathogenic Escherichia coli pathotype. New Microbes New Infect. 2014, 2, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Jiménez, D.; García-Meniño, I.; Herrera, A.; García, V.; López-Beceiro, A.M.; Alonso, M.P.; Blanco, J.; Mora, A. Genomic Characterization of Escherichia coli Isolates Belonging to a New Hetero-pathogenic aEPEC/ExPEC Pathotype O153:H10-A-ST10 eae-beta1 Occurred in Meat, Poultry, Wildlife and Human Diarrheagenic Samples. Antibiotics 2020, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Gati, N.S.; Middendorf-Bauchart, B.; Bletz, S.; Dobrindt, U.; Mellmann, A. Origin and Evolution of Hetero-pathogenic Shiga Toxin-Producing and Uropathogenic Escherichia coli Strains of Sequence Type 141. J. Clin. Microbiol. 2019, 58, e01309-19. [Google Scholar] [CrossRef]
- Iyoda, S.; Tamura, K.; Itoh, K.; Izumiya, H.; Ueno, N.; Nagata, K.; Togo, M.; Terajima, J.; Watanabe, H. Inducible stx2 phages are lysogenized in the enteroaggregative and other phenotypic Escherichia coli O86:HNM isolated from patients. FEMS Microbiol. Lett. 2000, 191, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozzoli, R.; Grande, L.; Michelacci, V.; Ranieri, P.; Maugliani, A.; Caprioli, A.; Morabito, S. Shiga toxin-converting phages and the emergence of new pathogenic Escherichia coli: A world in motion. Front. Cell Infect. Microbiol. 2014, 4, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheutz, F.; Cheasty, T.; Woodward, D.; Smith, H.R. Designation of O174 and O175 to temporary O groups OX3 and OX7, and six new E. coli O groups that include Verocytotoxin-producing E. coli (VTEC): O176, O177, O178, O179, O180 and O181. APMIS 2004, 112, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Eklund, M.; Scheutz, F.; Siitonen, A. Clinical isolates of non-O157 Shiga toxin-producing Escherichia coli: Serotypes, virulence characteristics, and molecular profiles of strains of the same serotype. J. Clin. Microbiol. 2001, 39, 2829–2834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ori, E.L.; Takagi, E.H.; Andrade, T.S.; Miguel, B.T.; Cergole-Novella, M.C.; Guth, B.E.C.; Hernandes, R.T.; Dias, R.C.B.; Pinheiro, S.R.S.; Camargo, C.H.; et al. Diarrhoeagenic Escherichia coli and Escherichia albertii in Brazil: Pathotypes and serotypes over a 6-year period of surveillance. Epidemiol. Infect. 2018, 147, e10. [Google Scholar] [CrossRef] [Green Version]
- García-Aljaro, C.; Muniesa, M.; Blanco, J.E.; Blanco, M.; Blanco, J.; Jofre, J.; Blanch, A.R. Characterization of Shiga toxin-producing Escherichia coli isolated from aquatic environments. FEMS Microbiol. Lett. 2005, 246, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Buvens, G.; Piérard, D. Low prevalence of STEC autotransporter contributing to biofilm formation (Sab) in verocytotoxin-producing Escherichia coli isolates of humans and raw meats. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1463–1465. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Mohamed, K.; The Agama Study Group; Achtman, M. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny and Escherichia core genomic diversity. Genom. Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Kislichkina, A.A.; Kartsev, N.N.; Skryabin, Y.P.; Sizova, A.A.; Kanashenko, M.E.; Teymurazov, M.G.; Kuzina, E.S.; Bogun, A.G.; Fursova, N.K.; Svetoch, E.A.; et al. Genomic Analysis of a Hetero-pathogenic Enteroaggregative Hemorrhagic Escherichia coli O181:H4 Strain Causing Colitis with Hemolytic-Uremic Syndrome. Antibiotics 2022, 11, 1416. [Google Scholar] [CrossRef]
- Fredericq, P. Colicins. Annu. Rev. Microbiol. 1957, 11, 7–22. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- Rodrigues, L.B.; Dos Santos, L.R.; Tagliari, V.Z.; Rizzo, N.N.; Trenhago, G.; de Oliveira, A.P.; Goetz, F.; do Nascimento, V.P. Quantification of biofilm production on polystyrene by Listeria, Escherichia coli and Staphylococcus aureus isolated from a poultry slaughterhouse. Braz. J. Microbiol. 2010, 41, 1082–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 7 March 2022).
- Detusheva, E.V.; Rodin, V.B.; Slukin, P.V.; Ershova, O.N.; Aleksandrova, I.A.; Kurdyumova, N.V.; Sazykina, S.Y.; Dyatlov, I.A.; Fursova, N.K. Susceptibility of nosocomial K. pneumoniae, P. aeruginosa, A. baumannii, and P. mirabilis strains to a chlorhexidine-based antiseptic preparation. Klin. Mikrobiol. Antimikrob. Himioter. 2015, 17, 57–66. (In Russian) [Google Scholar]
- Parma, Y.R.; Chacana, P.A.; Rogé, A.; Kahl, A.; Cangelosi, A.; Geoghegan, P.; Lucchesi, P.M.; Fernández-Miyakawa, M.E. Antibodies anti-Shiga toxin 2 B subunit from chicken egg yolk: Isolation, purification and neutralization efficacy. Toxicon 2011, 58, 380–388. [Google Scholar] [CrossRef]
- Gentry, M.K.; Dalrymple, J.M. Quantitative microtiter cytotoxicity assay for Shigella toxin. J. Clin. Microbiol. 1980, 12, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, T.; Fujii, J.; Yoshida, S.; Yutsudo, T. Reconstitution of active recombinant Shiga toxin (Stx)1 from recombinant Stx1-A and Stx1-B subunits independently produced by E. coli clones. FEMS Microbiol. Lett. 1999, 178, 13–18. [Google Scholar] [CrossRef]
- al-Jaufy, A.Y.; King, S.R.; Jackson, M.P. Purification and characterization of a Shiga toxin A subunit-CD4 fusion protein cytotoxic to human immunodeficiency virus-infected cells. Infect. Immun. 1995, 63, 3073–3078. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Božić, D.D.; Pavlović, B.; Milovanović, J.; Jotić, A.; Čolović, J.; Ćirković, I. Antibiofilm effects of amoxicillin-clavulanic acid and levofloxacin in patients with chronic rhinosinusitis with nasal polyposis. Eur. Arch. Otorhinolaryngol. 2018, 275, 2051–2059. [Google Scholar] [CrossRef]
- Soysal, N.; Mariani-Kurkdjian, P.; Smail, Y.; Liguori, S.; Gouali, M.; Loukiadis, E.; Fach, P.; Bruyand, M.; Blanco, J.; Bidet, P.; et al. Enterohemorrhagic Escherichia coli Hetero-pathogenic Pathotype O80:H2 as a New Therapeutic Challenge. Emerg. Infect. Dis. 2016, 22, 1604–1612. [Google Scholar] [CrossRef] [Green Version]
- Cointe, A.; Birgy, A.; Mariani-Kurkdjian, P.; Liguori, S.; Courroux, C.; Blanco, J.; Delannoy, S.; Fach, P.; Loukiadis, E.; Bidet, P.; et al. Emerging Multidrug-Resistant Hetero-pathogenic Pathotype Shiga Toxin-Producing Escherichia coli O80 and Related Strains of Clonal Complex 165, Europe. Emerg. Infect. Dis. 2018, 24, 2262–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Mayack, L.A.; Soracco, R.J.; Wilde, E.W.; Pope, D.H. Comparative effectiveness of chlorine and chlorine dioxide biocide regimes for biofouling control. Water Res. 1984, 18, 593–599. [Google Scholar] [CrossRef]
- Petrenko, N.F.; Mokienko, A.V. Chlorine dioxide as a means of eliminating biofilms. Bull. Odessa St. Acad. Civil. Engin. Architec. 2005, 19, 58–63. (In Russian) [Google Scholar]
- Exner, M.; Kramer, A.; Lajoie, L.; Gebel, J.; Engelhart, S.; Hartemann, P. Prevention and control of health care-associated waterborne infections in health care facilities. Am. J. Infect. Control 2005, 33, S26–S40. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Test | Description | SCPM-O-B-9427 O181:H4 | SCPM-O-B-6860 O104:H4 |
|---|---|---|---|
| BGAL | Beta-galactosidase | + | + |
| dGLU | d-glucose | + | + |
| OFF | Fermentation of glucose | + | + |
| dMAL | d-maltose | + | + |
| dMAN | d-mannitol | + | + |
| dMNE | d-mannose | + | + |
| Tyra | Tyrosinarylamidase | + | + |
| dSOR | d-sorbitol | + | + |
| SAC | sucrose | + | + |
| dTRE | d-trehalose | + | + |
| 5KG | 5-keto-d-gluconate | + | − |
| ILATk | l-lactate | − | + |
| AGAL | Alpha-galactosidase | + | − |
| ODC | Ornithine decarboxylase | + | + |
| LDC | Lysine decarboxylase | + | + |
| CMT | Kumarat | + | + |
| BGUR | Beta-glucuronidase | + | + |
| O129R | Resistance to vibriostatic agent O/129 | − | + |
| ELLM | Elman | + | + |
| Disinfectant Group | Active Ingredients | MBC, % | |||
|---|---|---|---|---|---|
| E. coli ATCC 25922 | E. coli SCPM-O-B-9427 | ||||
| P | B | P | B | ||
| Dezin | Chlorhexidine bigluconate 20% | 0.001 | 0.02 | 0.004 | 0.02 |
| Triosept-Oxy | Hydrogen peroxide 25 ± 7%, didecyldimethylammonium chloride 5 ± 1.5%, auxiliary component—phosphoric acid 1.5% | 0.005 | 0.03 | 0.03 | 0.06 |
| Neobak-Oxy | QAS-3%, polyhexamethylene guanidine hydrochloride-3% | 0.008 | 0.06 | 0.02 | 0.08 |
| SAT-22 | QAS complex 15%, N, N-bis (3-aminopropyl-dodecylamine) 4%, polyhexamethylene guanidine hydrochloride 5% | 0.008 | 0.1 | 0.01 | 0.1 |
| Biodez-Extra DVU | QAS complex 24%, Glutaraldehyde 7%, glyoxal 6% | 0.002 | 0.01 | 0.008 | 0.02 |
| Microbak Forte | Benzalkonium chloride 20%, dodecylbispropylenetriamine 5% | 0.01 | 0.04 | 0.01 | 0.04 |
| Biodez-Optima | Alkyldimethylbenzylammonium chloride 25% | 0.001 | 0.06 | 0.004 | 0.13 |
| Tristel-Fuse | Base: citric acid 5% Activator: sodium chlorite 0.5% | 0.5 | 0.13 | 1 | 0.25 |
| Antibiotics | MIC, mg/L (Interpretation) | |||
|---|---|---|---|---|
| E. coli ATCC 25922 | E. coli B-9427 | |||
| P | B | P | B | |
| Amoxicillin–clavulanic acid | 0.008 (S) | <0.0005 (S) | 0.008 (S) | <0.0005 (S) |
| Cefoperazone–sulbactam | <0.0005 (S) | <0.0005 (S) | <0.0005 (S) | <0.0005 (S) |
| Aztreonam | 0.001 (S) | 0.13 (S) | <0.0005 (S) | 0.13 (S) |
| Meropenem | <0.0005 (S) | <0.0005 (S) | <0.0005 (S) | <0.0005 (S) |
| Amikacin | 0.002 (S) | 0.002 (S) | 0.002 (S) | 0.002 (S) |
| Ciprofloxacin | 0.001 (S) | 0.004 (S) | <0.0005 (S) | 0.004 (S) |
| Fosfomycin | 0.01 (S) | 0.03 (S) | 0.02 (S) | 0.03 (S) |
| Trimethoprim–sulfamethoxazole | <0.0005 (S) | 0.002 (S) | <0.0005 (S) | 0.002 (S) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kartsev, N.N.; Detusheva, E.V.; Kalmantaeva, O.V.; Korobova, O.V.; Gerasimov, V.N.; Kombarova, T.I.; Borzilov, A.I.; Fursova, N.K.; Vereshchagin, A.N.; Svetoch, E.A. Hetero-Pathogenic O181:H4 EAHEC Strain of Sequence Type ST678 Associated with Hemolytic–Uremic Syndrome in Schoolchildren in Russia. Microorganisms 2023, 11, 1771. https://doi.org/10.3390/microorganisms11071771
Kartsev NN, Detusheva EV, Kalmantaeva OV, Korobova OV, Gerasimov VN, Kombarova TI, Borzilov AI, Fursova NK, Vereshchagin AN, Svetoch EA. Hetero-Pathogenic O181:H4 EAHEC Strain of Sequence Type ST678 Associated with Hemolytic–Uremic Syndrome in Schoolchildren in Russia. Microorganisms. 2023; 11(7):1771. https://doi.org/10.3390/microorganisms11071771
Chicago/Turabian StyleKartsev, Nikolay N., Elena V. Detusheva, Olga V. Kalmantaeva, Olga V. Korobova, Vladimir N. Gerasimov, Tatiana I. Kombarova, Aleksander I. Borzilov, Nadezhda K. Fursova, Anatoly N. Vereshchagin, and Edward A. Svetoch. 2023. "Hetero-Pathogenic O181:H4 EAHEC Strain of Sequence Type ST678 Associated with Hemolytic–Uremic Syndrome in Schoolchildren in Russia" Microorganisms 11, no. 7: 1771. https://doi.org/10.3390/microorganisms11071771