
Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Causing Nosocomial Meningitis in the Neurological Intensive Care Unit
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research, Bioethical Requirements, and Patients
2.2. Bacterial Isolates and Growing
2.3. Antimicrobial Susceptibility
2.4. Biofilm Formation
2.5. Whole-Genome Sequencing, Assembly, and Annotation
2.6. Multilocus Sequence Typing, KL-Typing, and OCL-Typing
2.7. Antimicrobial Resistance and Virulence Genes
2.8. Phylogenetic Analysis
2.9. Nucleotide Sequences Accession Numbers
3. Results
3.1. Patients and Clinical Data
3.2. Sequence Types, Capsular Polysaccharide Locus Types, and Lipooligosaccharide Outer Core Locus Types
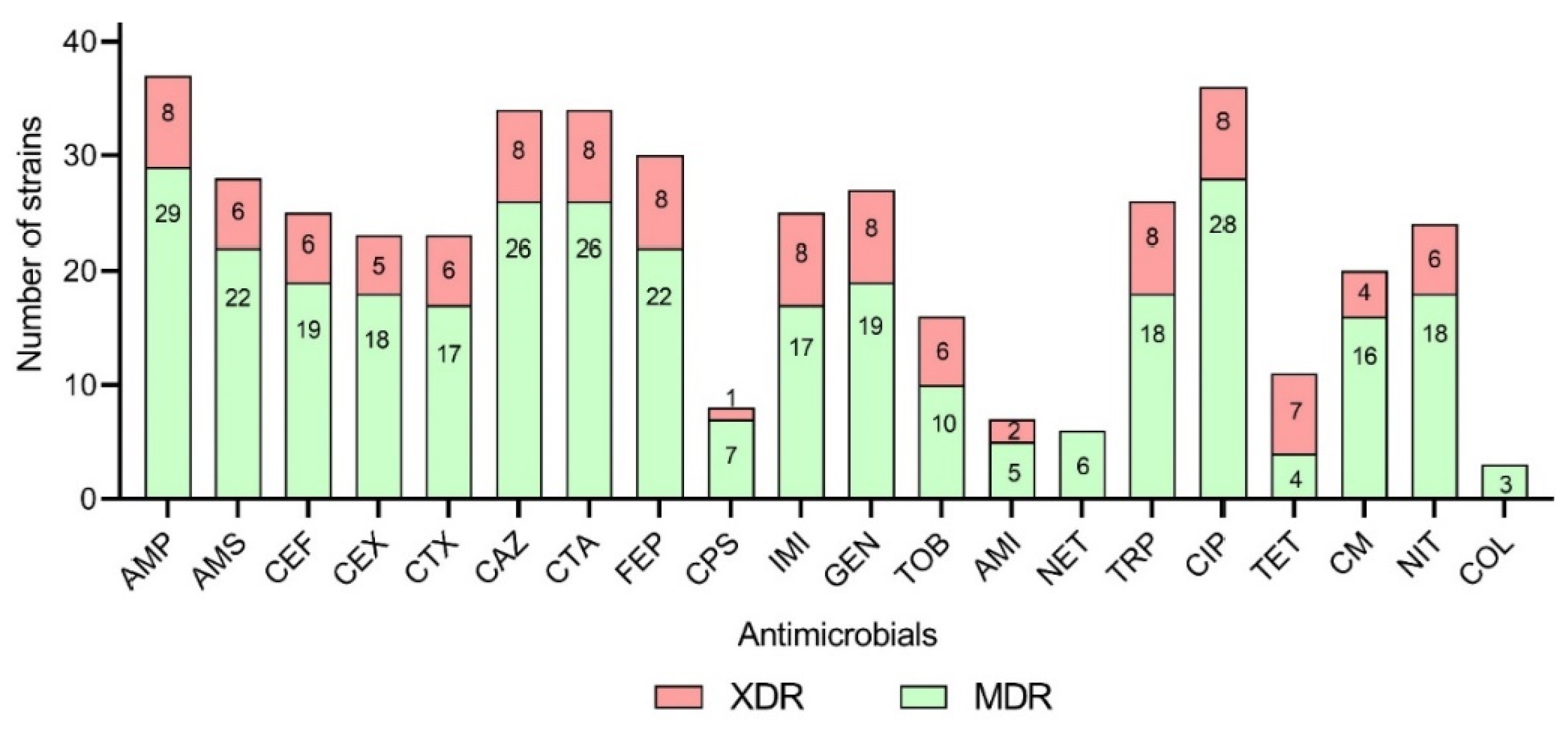
3.3. Antimicrobial Susceptibility
3.4. Biofilm Formation
3.5. Whole-Genome Analysis
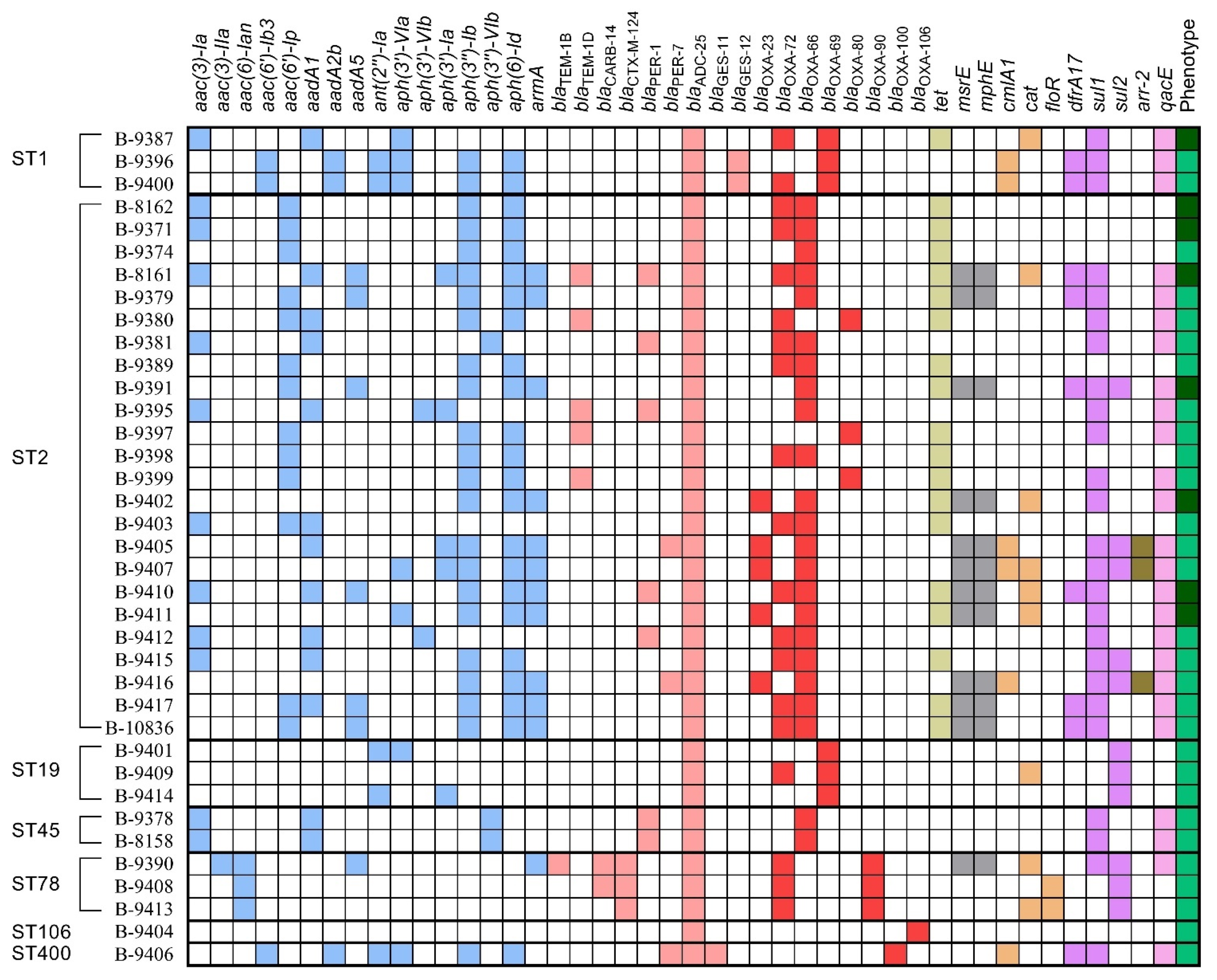
3.6. The Resistome Identification and Analysis
3.7. The Virulome Identification and Analysis
3.8. Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. WHO pathogens priority list working group. discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Lescure, F.X.; Bouadma, L.; Nguyen, D.; Parisey, M.; Wicky, P.H.; Behillil, S.; Gaymard, A.; Bouscambert-Duchamp, M.; Donati, F.; Le Hingrat, Q.; et al. Clinical and virological data of the first cases of COVID-19 in Europe: A case series. Lancet Infect. Dis. 2020, 20, 697–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Fu, Y.; Hua, X.; Xu, Q.; Lan, P.; Jiang, Y.; Yu, Y.; Zhou, Z. Acinetobacter baumannii strains isolated from cerebrospinal fluid (CSF) and bloodstream analysed by cgMLST: The dominance of clonal complex CC92 in CSF infections. Int. J. Antimicrob. Agents. 2021, 58, 106404. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Penteado-Filho, S.R.; Amarante, D.; Andrade, M.A.; Borba, L.A. Mortality rate in patients with nosocomial Acinetobacter meningitis from a Brazilian hospital. Braz. J. Infect. Dis. 2010, 14, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Chusri, S.; Sakarunchai, I.; Kositpantawong, N.; Panthuwong, S.; Santimaleeworagun, W.; Pattharachayakul, S.; Singkhamanan, K.; Doi, Y. Outcomes of adjunctive therapy with intrathecal or intraventricular administration of colistin fo.; post-neurosurgical meningitis and ventriculitis due to carbapenem-resistant Acinetobacter baumannii. Int. J. Antimicrob. Agents 2018, 51, 646–650. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Lopez-Otsoa, F.; Gallego, L.; Towner, K.J.; Tysall, L.; Woodford, N.; Livermore, D.M. Endemic carbapenem resistance associated with OXA-40 carbapenemase among Acinetobacter baumannii isolates from a hospital in northern Spain. J. Clin. Microbiol. 2002, 40, 4741–4743. [Google Scholar] [CrossRef] [Green Version]
- Dijkshoorn, L.; Aucken, H.; Gerner-Smidt, P.; Janssen, P.; Kaufmann, M.E.; Garaizar, J.; Ursing, J.; Pitt, T.L. Comparison of outbreak and nonoutbreak Acinetobacter baumannii strains by genotypic and phenotypic methods. J. Clin. Microbiol. 1996, 34, 1519–1525. [Google Scholar] [CrossRef]
- Zarrilli, R.; Pournaras, S.; Giannouli, M.; Tsakris, A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int. J. Antimicrob. Agents 2013, 41, 11–19. [Google Scholar] [CrossRef]
- Giannouli, M.; Antunes, L.C.; Marchetti, V.; Triassi, M.; Visca, P.; Zarrilli, R. Virulencerelated traits of epidemic Acinetobacter baumannii strains belonging to the international clonal lineages I–III and to the emerging genotypes ST25 and ST78. BMC Infect. Dis. 2013, 13, 282. [Google Scholar] [CrossRef] [Green Version]
- Mayanskiy, N.; Chebotar, I.; Alyabieva, N.; Kryzhanovskaya, O.; Savinova, T.; Turenok, A.; Bocharova, Y.; Lazareva, A.; Polikarpova, S.; Karaseva, O. Emergence of the uncommon clone ST944/ST78 carrying blaOXA-40-like and blaCTX-M-like genes among carbapenem-nonsusceptible Acinetobacter baumannii in Moscow, Russia. Microb. Drug Resist. 2017, 23, 864–870. [Google Scholar] [CrossRef]
- Cayô, R.; Rodrigues-Costa, F.; Matos, A.P.; Carvalhaes, C.G.; Jové, T.; Gales, A.C. Identification of a new integron harboring bla(IMP-10) in carbapenem-resistant Acinetobacter baumannii clinical isolates. Antimicrob. Agents Chemother. 2015, 59, 3687–3689. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Nordmann, P. Carbapenem resistance in Acinetobacter baumannii: Mechanisms and epidemiology. Clin. Microbiol. Infect. 2006, 12, 826–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrilli, R.; Bagattini, M.; Migliaccio, A.; Esposito, E.P.; Triassi, M. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii in Italy. Ann. Ig. 2021, 33, 401–409. [Google Scholar] [CrossRef]
- Wohlfarth, E.; Kresken, M.; Higgins, P.G.; Stefanik, D.; Wille, J.; Hafner, D.; Körber-Irrgang, B.; Seifert, H.; Study Group “Antimicrobial Resistance” of the Paul-Ehrlich-Society for Infection Therapy. The evolution of carbapenem resistance determinants and major epidemiological lineages among carbapenem-resistant Acinetobacter baumannii isolates in Germany, 2010–2019. Int. J. Antimicrob. Agents 2022, 60, 106689. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii antibiotic resistance mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Gebhardt, M.J.; Shuman, H.A. GigA and GigB are master regulators of antibiotic resistance, stress responses, and virulence in Acinetobacter baumannii. J. Bacteriol. 2017, 199, e00066-17. [Google Scholar] [CrossRef] [Green Version]
- Maharjan, R.P.; Sullivan, G.J.; Adams, F.G.; Shah, B.S.; Hawkey, J.; Delgado, N.; Semenec, L.; Dinh, H.; Li, L.; Short, F.L.; et al. DksA is a conserved master regulator of stress response in Acinetobacter baumannii. Nucleic Acids Res. 2023, 9, gkad341. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Ershova, K.; Savin, I.; Kurdyumova, N.; Wong, D.; Danilov, G.; Shifrin, M.; Alexandrova, I.; Sokolova, E.; Fursova, N.; Zelman, V.; et al. Implementing an infection control and prevention program decreases the incidence of healthcare-associated infections and antibiotic resistance in a Russian neuro-ICU. Antimicrob. Resist. Infect. Control 2018, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babalska, Z.Ł.; Korbecka-Paczkowska, M.; Karpiński, T.M. Wound antiseptics and European Guidelines for antiseptic application in wound treatment. Pharmaceuticals 2021, 14, 1253. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, S.; Rajenderan, S.; Laishram, S.; Anandan, S.; Balaji, V.; Biswas, I. Biofilm formation and motility depend on the nature of the Acinetobacter baumannii clinical isolates. Front. Public. Health. 2016, 4, 105. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 8918–8999. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K. Preparation of genomic DNA from bacteria. Curr. Protoc. Mol. Biol. 2001, 56, 2–4. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Kurdyumova, N.V.; Savin, I.A.; Ershova, O.N.; Aleksandrova, I.A.; Sazykina, S.J.; Gadjieva, O.A.; Danilov, G.V.; Shifrin, M.A.; Fursova, N.K. Nosocomial meningitis caused by Acinetobacter baumannii in ICU patients. Rus. J. Anaesthesiol. Reanimatol. (Anesteziologiya I Reanimatologiya) 2019, 4, 43–49. [Google Scholar] [CrossRef]
- Zheng, G.; Wang, S.; Lv, H.; Zhang, G. Nomogram analysis of clinical characteristics and mortality risk factor of non-fermentative gram-negative bacteria-induced post-neurosurgical meningitis. Infect. Drug Resist. 2022, 15, 6379–6389. [Google Scholar] [CrossRef]
- Panic, H.; Gjurasin, B.; Santini, M.; Kutlesa, M.; Papic, N. Etiology and outcomes of healthcare-associated meningitis and ventriculitis-a single center cohort study. Infect. Dis. Rep. 2022, 14, 420–427. [Google Scholar] [CrossRef]
- Hamidian, M.; Nigro, S.J. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microb. Genom. 2019, 5, e000306. [Google Scholar] [CrossRef]
- Eigenbrod, T.; Reuter, S.; Gross, A.; Kocer, K.; Günther, F.; Zimmermann, S.; Heeg, K.; Mutters, N.T.; Nurjadi, D. Molecular characterization of carbapenem-resistant Acinetobacter baumannii using WGS revealed missed transmission events in Germany from 2012–15. J. Antimicrob. Chemother. 2019, 74, 3473–3480. [Google Scholar] [CrossRef]
- Yang, J.L.; Yang, C.J.; Chuang, Y.C.; Sheng, W.H.; Chen, Y.C.; Chang, S.C. Association of capsu-lar polysaccharide locus 2 with prognosis of Acinetobacter baumannii bacteraemia. Emerg. Microbes Infect. 2022, 11, 83–90. [Google Scholar] [CrossRef]
- Arbatsky, N.P.; Kasimova, A.A.; Shashkov, A.S.; Shneider, M.M.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; Yanushevich, Y.G.; Azizov, I.S.; et al. Structure of the K128 capsular polysaccharide produced by Acinetobacter baumannii. Carbohydr. Res. 2019, 485, 107814. [Google Scholar] [CrossRef]
- Sorbello, B.M.; Cahill, S.M.; Kenyon, J.J. Identification of further variation at the lipooligosac-charide outer core locus in Acinetobacter baumannii genomes and extension of the OCL reference sequence database for Kaptive. Microb. Genom. 2023, 9, 001042. [Google Scholar] [CrossRef]
- Stratev, A.; Tanova, R.; Dimov, S.; Mitov, I.; Strateva, T. Clonal spread of carbapenem-resistant Acinetobacter baumannii isolates among Bulgarian critically ill patients undergoing renal replacement therapy (2016–2018). Infect. Dis. 2020, 52, 430–433. [Google Scholar] [CrossRef]
- Strateva, T.V.; Sirakov, I.; Stoeva, T.J.; Stratev, A.; Peykov, S. Phenotypic and molecular characteristics of carbapenem-resistant Acinetobacter baumannii isolates from Bulgarian Intensive Care Unit patients. Microorganisms 2023, 11, 875. [Google Scholar] [CrossRef]
- Munari, M.; Franzoi, F.; Sergi, M.; De Cassai, A.; Geraldini, F.; Grandis, M.; Caravello, M.; Boscolo, A.; Navalesi, P. Extensively drug-resistant and multidrug-resistant gram-negative pathogens in the neurocritical intensive care unit. Acta Neurochir. 2022, 164, 859–865. [Google Scholar] [CrossRef]
- Ayobami, O.; Willrich, N.; Harder, T.; Okeke, I.N.; Eckmanns, T.; Markwart, R. The incidence and prevalence of hospital-acquired (carbapenem-resistant) Acinetobacter baumannii in Europe, Eastern Mediterranean and Africa: A systematic review and meta-analysis. Emerg. Microbes Infect. 2019, 8, 1747–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, S.M.; Hussein, A.F.A.; Al-Agamy, M.H.; Radwan, H.H.; Zafer, M.M. Genetic configuration of genomic resistance islands in Acinetobacter baumannii clinical isolates from Egypt. Front. Microbiol. 2022, 13, 878912. [Google Scholar] [CrossRef] [PubMed]
- Farzana, R.; Swedberg, G.; Giske, C.G.; Hasan, B. Molecular and genetic characterization of emerging carbapenemase-producing Acinetobacter baumannii strains from patients and hospital environments in Bangladesh. Infect. Prev. Pract. 2022, 4, 100215. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Luan, G.; Wang, Y.; Chang, Y.; Zhang, C.; Yang, J.; Deng, S.; Ling, B.; Jia, X. Coexistence of blaOXA-23 with armA in quinolone-resistant Acinetobacter baumannii from a Chinese university hospital. Diagn. Microbiol. Infect. Dis. 2016, 84, 230–231. [Google Scholar] [CrossRef]
- Chukamnerd, A.; Singkhamanan, K.; Chongsuvivatwong, V.; Palittapongarnpim, P.; Doi, Y.; Pomwised, R.; Sakunrang, C.; Jeenkeawpiam, K.; Yingkajorn, M.; Chusri, S.; et al. Whole-genome analysis of carbapenem-resistant Acinetobacter baumannii from clinical isolates in Southern Thailand. Comput. Struct. Biotechnol. J. 2022, 20, 545–558. [Google Scholar] [CrossRef]
- Sheck, E.A.; Edelstein, M.V.; Sukhorukova, M.V.; Ivanchik, N.V.; Skleenova, E.Y.; Dekhnich, A.V.; Azizov, I.S.; Kozlov, R.S. Epidemiology and genetic diversity of colistin nonsusceptible nosocomial Acinetobacter baumannii strains from Russia for 2013–2014. Can. J. Infect. Dis. Med. Microbiol. 2017, 2017, 1839190. [Google Scholar] [CrossRef] [Green Version]
- Nor A’shimi, M.H.; Alattraqchi, A.G.; Mohd Rani, F.; Rahman, N.I.A.; Ismail, S.; Abdullah, F.H.; Othman, N.; Cleary, D.W.; Clarke, S.C.; Yeo, C.C. Biocide susceptibilities and biofilm-forming capacities of Acinetobacter baumannii clinical isolates from Malaysia. J. Infect. Dev. Ctries 2019, 13, 626–633. [Google Scholar] [CrossRef]
- Schmitt, B.L.; Leal, B.F.; Leyser, M.; de Barros, M.P.; Trentin, D.S.; Ferreira, C.A.S.; de Oliveira, S.D. Increased ompW and ompA expression and higher virulence of Acinetobacter baumannii persister cells. BMC Microbiol. 2023, 23, 157. [Google Scholar] [CrossRef]
- Jones, C.L.; Clancy, M.; Honnold, C.; Singh, S.; Snesrud, E.; Onmus-Leone, F.; McGann, P.; Ong, A.C.; Kwak, Y.; Waterman, P.; et al. Fatal outbreak of an emerging clone of extensively drug-resistant Acinetobacter baumannii with enhanced virulence. Clin. Infect. Dis. 2015, 61, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Sitthisak, S. Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter baumannii clinical isolates. Int. Microbiol. 2016, 19, 121–129. [Google Scholar] [CrossRef]
- Gheorghe, I.; Barbu, I.C.; Surleac, M.; Sârbu, I.; Popa, L.I.; Paraschiv, S.; Feng, Y.; Lazăr, V.; Chifiriuc, M.C.; Oţelea, D.; et al. Subtypes, resistance and virulence platforms in extended-drug resistant Acinetobacter baumannii Romanian isolates. Sci. Rep. 2021, 11, 13288. [Google Scholar] [CrossRef]
- Law, S.K.K.; Tan, H.S. The role of quorum sensing, biofilm formation, and iron acquisition as key virulence mechanisms in Acinetobacter baumannii and the corresponding anti-virulence strategies. Microbiol. Res. 2022, 260, 127032. [Google Scholar] [CrossRef]
- Harrison, A.; Fernando, D.M.; Mason, K.M.; Santana, E.; Loewen, P.C.; Kumar, A.; Liu, Y. KatG and KatE confer Acinetobacter resistance to hydrogen peroxide but sensitize bacteria to killing by phagocytic respiratory burst. Life Sci. 2016, 148, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Cook-Libin, S.; Sykes, E.M.E.; Kornelsen, V.; Kumar, A. Iron acquisition mechanisms and their role in the virulence of Acinetobacter baumannii. Infect. Immun. 2022, 90, e0022322. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Gender, Age | Date | Patient’s Diagnosis | Infections | Stay in the ICU, Days | Outcome |
|---|---|---|---|---|---|---|
| #1 | M, 16 | 15 January 2013 | Medulloblastoma of the fourth ventricle | MG, RTI, SSI, GD | 186 | Survival |
| #2 | M, 46 | 27 February 2013 | Anaplastic astrocytoma | MG, RTI, UTI | 24 | Died |
| #3 | M, 50 | 4 February 2014 | Cholesteatoma | MG, BI, GD | 184 | Survival |
| #4 | F, 65 | 25 June 2014 | Suprasellar craniopharyngioma | MG | 31 | Died |
| #5 | F, 24 | 5 November 2014 | Anterior cerebral artery aneurysm | MG | 62 | Survival |
| #6 | F, 52 | 19 January 2015 | Craniofacial tumor | MG, SSI, UTI | 34 | Died |
| #7 | M, 48 | 4 February 2015 | Pilocytic astrocytoma | MG, RTI | 43 | Survival |
| #8 | M, 33 | 29 September 2015 | Traumatic brain injury | MG, RTI, SSI | 45 | Survival |
| #9 | F, 2 | 13 November 2015 | Cerebellar astrocytoma | MG, RTI, SSI | 62 | Survival |
| #10 | M, 57 | 7 December 2015 | Giant cell tumor of the middle cranial fosses | MG | 15 | Survival |
| #11 | M, 44 | 9 December 2015 | Anterior cerebral artery aneurysm | MG, RTI | 8 | Died |
| #12 | M, 43 | 18 May 2016 | Traumatic brain injury | MG | 15 | Survival |
| #13 | M, 43 | 15 June 2016 | Cerebellum tumor | MG, RTI, SSI | 63 | Died |
| #14 | M, 12 | 16 June 2016 | Giant craniopharyngioma | MG | 62 | Survival |
| #15 | M, 28 | 1 August 2016 | Atypical parasagittal meningioma | MG, SSI | 49 | Survival |
| #16 | F, 60 | 2 August 2016 | Giant olfactory meningioma | MG | 229 | Died |
| #17 | M, 75 | 16 August 2016 | Ependymoma | MG, RTI, UTI | 136 | Died |
| #18 | M, 35 | 12 September 2016 | Glioblastoma | MG | 98 | Died |
| #19 | F, 57 | 10 October 2016 | Traumatic brain injury | MG, RTI, UTI | 14 | Survival |
| #20 | M, 67 | 29 November 2016 | Sub-ependymoma | MG | 31 | Survival |
| #21 | M, 61 | 4 September 2017 | Craniopharyngioma | MG | 133 | Survival |
| #22 | F, 61 | 12 December 2017 | Hemangiopericytoma | MG | 24 | Survival |
| #23 | F, 36 | 15 January 2018 | Giant meningioma | MG, RTI, SSI | 65 | Died |
| #24 | M, 55 | 21 February 2018 | Hemangioblastoma | MG | 46 | Survival |
| #25 | M, 57 | 20 June 2018 | Gunshot wound head trauma | MG, RTI | 181 | Survival |
| #26 | F, 58 | 19 September 2018 | Glioma | MG | 52 | Died |
| #27 | M, 43 | 1 October 2018 | Hemangioblastoma of the cerebellum | MG, RTI | 53 | Survival |
| #28 | M, 53 | 23 October 2018 | Traumatic brain injury | MG, RTI | 58 | Survival |
| #29 | M, 53 | 12 November 2018 | Traumatic brain injury | MG, RTI, UTI, GD | 103 | Survival |
| #30 | M, 61 | 3 December 2018 | Post-hemorrhagic hydrocephalus | MG, RTI, GD | 19 | Died |
| #31 | M, 64 | 15 May 2019 | Skull base chordoma | MG, RTI | 25 | Died |
| #32 | F, 25 | 20 May 2019 | Traumatic brain injury | MG, RTI | 52 | Survival |
| #33 | M, 26 | 18 July 2019 | Traumatic brain injury | MG, RTI | 45 | Survival |
| #34 | M, 51 | 6 August 2019 | Ruptured middle cerebral artery aneurysms | MG | 26 | Survival |
| #35 | M, 47 | 27 January 2020 | Giant skull base chordoma | MG | 89 | Died |
| #36 | F, 52 | 28 August 2020 | Syringomyelia of the cervical spinal cord | MG, RTI | 71 | Died |
| #37 | F, 45 | 8 September 2020 | Hemangiopericytoma | MG, GD | 69 | Survival |
| Case | Strain | ST | KL | OCL | AMR Phenotype | AM gr. | ARP |
|---|---|---|---|---|---|---|---|
| #1 | B-8162 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, TET, NIT | 9 | XDR |
| #2 | B-9371 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, NIT | 8 | XDR |
| #3 | B-9374 | 2 | 9 | 1 | AG, CAR, FQN, CEF, PEN, TET, CM, NIT | 8 | MDR |
| #4 | B-8161 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, TET, CM, NIT | 10 | XDR |
| #5 | B-9378 | 45 | 77 | 1 | AG, CAR, FQN, CEF, PEN, PEN-IN, CM, NIT | 8 | MDR |
| #6 | B-9379 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, TET, CM, NIT | 9 | MDR |
| #7 | B-9380 | 2 | 2 | 1 | AG, FQN, CEF, PEN, PEN-IN, TET, CM, NIT | 8 | MDR |
| #8 | B-9381 | 2 | 9 | 1 | AG, CAR, FQN, CEF, PEN, PEN-IN, CM, NIT | 8 | MDR |
| #9 | B-9387 | 1 | 17 | 1 | AG, CAR, FQN, CEF, PEN, TET, CM, NIT | 8 | XDR |
| #10 | B-9389 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, CM, NIT | 8 | MDR |
| #11 | B-10836 | 2 | 9 | 1 | AG, CAR, FQN, CEF, PEN, PEN-IN, CM, NIT | 8 | MDR |
| #12 | B-9390 | 78 | 49 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, CM, NIT | 9 | MDR |
| #13 | B-9391 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, TET, CM, NIT | 9 | XDR |
| #14 | B-9395 | 2 | 165 | 1 | AG, FQN, CEF, PEN, PEN-IN, CM, NIT | 7 | MDR |
| #15 | B-9397 | 2 | 213 | 1 | AG, FQN, CEF, PEN, PEN-IN, POL, NIT | 7 | MDR |
| #16 | B-9398 | 2 | 2 | 1 | AG, FQN, CEF, SUL, PEN, PEN-IN, POL, NIT | 8 | MDR |
| #17 | B-8158 | 45 | 9 | 1 | AG, FQN, CEF, PEN, POL, CM | 6 | MDR |
| #18 | B-9399 | 2 | 213 | 1 | AG, FQN, CEF, PEN, PEN-IN, TET, CM, NIT | 8 | MDR |
| #19 | B-9396 | 1 | 4 | 1 | AG, FQN, CEF, SUL, PEN, PEN-IN, CM, NIT | 8 | MDR |
| #20 | B-9400 | 1 | 4 | 1 | AG, FQN, CEF, SUL, PEN, PEN-IN, CM, NIT | 8 | MDR |
| #21 | B-9401 | 19 | 91 | 5 | AG, FQN, CEF, SUL, PEN, PEN-IN, CM, NIT | 8 | MDR |
| #22 | B-9402 | 2 | 3 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, TET, CM, NIT | 10 | XDR |
| #23 | B-9403 | 2 | 9 | 1 | AG, CAR, FQN, CEF, PEN, CM, NIT | 7 | MDR |
| #24 | B-9404 | 106 | 104 | 6 | CEF, SUL, PEN, PEN-IN, CM, NIT | 6 | MDR |
| #25 | B-9405 | 2 | 235 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN | 7 | MDR |
| #26 | B-9406 | 400 | 15 | 1 | AG, FQN, CEF, SUL, PEN-IN | 6 | MDR |
| #27 | B-9407 | 2 | 235 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN | 7 | MDR |
| #28 | B-9408 | 78 | 3 | 1 | CAR, FQN, CEF, SUL, PEN, PEN-IN | 6 | MDR |
| #29 | B-9409 | 19 | 91 | 5 | CAR, FQN, CEF, SUL, PEN, PEN-IN | 6 | MDR |
| #30 | B-9410 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, TET | 8 | XDR |
| #31 | B-9411 | 2 | 3 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN, TET | 8 | XDR |
| #32 | B-9412 | 2 | 9 | 1 | CAR, FQN, CEF, PEN, PEN-IN, NIT | 6 | MDR |
| #33 | B-9413 | 78 | 15 | 1 | CAR, FQN, CEF, SUL, PEN, PEN-IN | 6 | MDR |
| #34 | B-9414 | 19 | 91 | 5 | AG, FQN, CEF, SUL, PEN, PEN-IN | 6 | MDR |
| #35 | B-9415 | 2 | 2 | 1 | AG, CAR, FQN, CEF, SUL PEN, | 6 | MDR |
| #36 | B-9416 | 2 | 49 | 1 | AG, CAR, FQN, CEF, SUL, PEN, POL | 7 | MDR |
| #37 | B-9417 | 2 | 9 | 1 | AG, CAR, FQN, CEF, SUL, PEN, PEN-IN | 7 | MDR |
| Strain | MIC, mg/L | Strain | MIC, mg/L | ||
|---|---|---|---|---|---|
| CHD | BZK | CHD | BZK | ||
| B-9387 | 32 | 8 | B-9389 | 32 | 8 |
| B-9396 | 64 | 8 | B-9395 | 32 | 8 |
| B-9400 | 64 | 8 | B-9397 | 32 | 8 |
| B-9371 | 32 | 8 | B-9402 | 32 | 4 |
| B-8161 | 32 | 8 | B-9405 | 16 | 4 |
| B-9379 | 32 | 8 | B-9407 | 32 | 8 |
| B-9391 | 32 | 8 | B-9412 | 32 | 8 |
| B-9398 | 32 | 8 | B-9417 | 32 | 8 |
| B-9399 | 32 | 8 | B-9401 | 32 | 8 |
| B-9403 | 32 | 8 | B-9409 | 16 | 8 |
| B-9410 | 32 | 8 | B-9414 | 16 | 4 |
| B-9411 | 16 | 4 | B-8158 | 16 | 8 |
| B-9415 | 8 | 4 | B-9378 | 16 | 8 |
| B-9416 | 32 | 8 | B-9390 | 16 | 4 |
| B-10836 | 32 | 8 | B-9408 | 32 | 8 |
| B-8162 | 32 | 8 | B-9413 | 16 | 4 |
| B-9374 | 32 | 8 | B-9404 | 16 | 4 |
| B-9380 | 32 | 8 | B-9406 | 32 | 8 |
| B-9381 | 32 | 8 | |||
| Strain | GenBank ID | GC Content, % | Genes (Total) | CDSs (Total) | Genes (Coding) | Genes (RNA) | rRNA Genes (5S, 16S, 23S) | tRNAs | ncRNAs | Pseudo Genes |
|---|---|---|---|---|---|---|---|---|---|---|
| B-8162 | JAROBQ000000000 | 39.8 | 3929 | 3858 | 3801 | 71 | 1, 1, 1 | 64 | 4 | 57 |
| B-9371 | JASKJB000000000 | 42.4 | 3722 | 3650 | 3600 | 72 | 0, 2, 3 | 63 | 4 | 50 |
| B-9374 | JASKJA000000000 | 42.4 | 3873 | 3802 | 3745 | 71 | 0, 2, 1 | 64 | 4 | 57 |
| B-8161 | JAROBM000000000 | 39.2 | 3848 | 3778 | 3723 | 70 | 1, 1, 1 | 63 | 4 | 55 |
| B-9378 | JAROBU000000000 | 39.6 | 3707 | 3636 | 3576 | 71 | 1, 1, 1 | 64 | 4 | 60 |
| B-9379 | JASKIZ000000000 | 42.4 | 3704 | 3632 | 3580 | 72 | 1, 2, 2 | 63 | 4 | 52 |
| B-9380 | JAROBS000000000 | 39.9 | 3951 | 3880 | 3829 | 71 | 1, 1, 1 | 64 | 4 | 51 |
| B-9381 | JAROBO000000000 | 39.3 | 3867 | 3797 | 3745 | 70 | 1, 1, 1 | 63 | 4 | 52 |
| B-9387 | JAROBG000000000 | 39.2 | 3877 | 3807 | 3737 | 70 | 1, 1, 1 | 63 | 4 | 70 |
| B-9389 | JASKIY000000000 | 42.2 | 3596 | 3524 | 3475 | 72 | 0, 2, 3 | 63 | 4 | 49 |
| B-9390 | JASKIX000000000 | 42.4 | 3890 | 3820 | 3764 | 70 | 1, 1, 1 | 63 | 4 | 56 |
| B-9391 | JASKIW000000000 | 42.1 | 3679 | 3608 | 3555 | 71 | 0, 2, 2 | 63 | 4 | 53 |
| B-9395 | JASKIV000000000 | 42.9 | 3887 | 3816 | 3758 | 71 | 0, 2, 1 | 64 | 4 | 58 |
| B-9397 | JASKIU000000000 | 42.2 | 3795 | 3725 | 3676 | 70 | 0, 1, 1 | 64 | 4 | 49 |
| B-9398 | JAROBR000000000 | 39.8 | 3965 | 3894 | 3840 | 71 | 1, 1, 1 | 64 | 4 | 54 |
| B-8158 | JAROBT000000000 | 40.3 | 3735 | 3664 | 3605 | 71 | 1, 1, 1 | 64 | 4 | 59 |
| B-9399 | JASKIT000000000 | 42.0 | 3810 | 3741 | 3692 | 69 | 0, 1, 1 | 63 | 4 | 49 |
| B-9396 | JAROBB000000000 | 40.1 | 3855 | 3785 | 3720 | 70 | 1, 1, 1 | 63 | 4 | 65 |
| B-9400 | JAROBI000000000 | 39.8 | 3879 | 3809 | 3744 | 70 | 1, 1, 1 | 63 | 4 | 65 |
| B-9401 | JASKIR000000000 | 40.6 | 3624 | 3553 | 3496 | 71 | 1, 1, 1 | 63 | 5 | 57 |
| B-9402 | JASKIQ000000000 | 39.3 | 3797 | 3726 | 3664 | 71 | 1, 1, 1 | 64 | 5 | 62 |
| B-9403 | JASKIP000000000 | 39.2 | 4019 | 3949 | 3891 | 70 | 1, 1, 1 | 63 | 5 | 58 |
| B-9404 | JASKIO000000000 | 39.7 | 3821 | 3752 | 3644 | 69 | 0, 1, 1 | 63 | 5 | 108 |
| B-9405 | JASKIN000000000 | 39.2 | 3957 | 3887 | 3830 | 70 | 1, 1, 1 | 70 | 5 | 57 |
| B-9406 | JASKIM000000000 | 39.3 | 3803 | 3733 | 3675 | 70 | 1, 1, 1 | 63 | 4 | 58 |
| B-9407 | JASKIL000000000 | 39.3 | 3956 | 3886 | 3830 | 70 | 1, 1, 1 | 63 | 4 | 56 |
| B-9408 | JASKIK000000000 | 39.9 | 3804 | 3733 | 3682 | 71 | 1, 1, 1 | 64 | 4 | 51 |
| B-9409 | JASKIJ000000000 | 39.5 | 3845 | 3775 | 3716 | 70 | 1, 1, 1 | 62 | 5 | 59 |
| B-9410 | JASKII000000000 | 39.4 | 3853 | 3783 | 3728 | 70 | 1, 1, 1 | 63 | 4 | 55 |
| B-9411 | JASKIH000000000 | 38.6 | 3933 | 3862 | 3800 | 71 | 1, 1, 1 | 64 | 4 | 62 |
| B-9412 | JASKIG000000000 | 39.1 | 3654 | 3584 | 3532 | 70 | 1, 1, 1 | 63 | 4 | 52 |
| B-9413 | JASKIF000000000 | 39.4 | 3745 | 3674 | 3626 | 71 | 1, 1, 1 | 64 | 4 | 48 |
| B-9414 | JASKIE000000000 | 39.4 | 3665 | 3593 | 3538 | 72 | 1, 1, 1 | 64 | 5 | 55 |
| B-9415 | JASKID000000000 | 39.4 | 3939 | 3868 | 3813 | 71 | 1, 1, 1 | 64 | 4 | 55 |
| B-9416 | JASKIC000000000 | 39.4 | 3928 | 3859 | 3797 | 69 | 0, 1, 1 | 63 | 4 | 62 |
| B-9417 | JASKIB000000000 | 39.1 | 3743 | 3673 | 3622 | 70 | 1, 1, 1 | 63 | 4 | 51 |
| Strain | ST | katA (AKA32231.1) | katE (AKA32163.1) | katG (AKA33165.1) | katX (AKA31788.1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ident., % | SNP | SAV | Ident., % | SNP | SAV | Ident., % | SNP | SAV | Ident., % | SNP | SAV | ||
| B-9387 | 1 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 97.85 | 25 | 6 |
| B-9396 | 1 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 |
| B-9400 | 1 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 |
| B-9371 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-8161 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9379 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9391 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9398 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9399 | 2 | 97,65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9403 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9410 | 2 | 97.56 | 26 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9411 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9415 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9416 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-10836 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-8162 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9374 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9380 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9381 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9389 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9395 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9397 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9402 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9405 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9407 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9412 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9417 | 2 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | - | - | - |
| B-9401 | 19 | - | - | - | - | - | - | 100 | 0 | 0 | 100 | 0 | 0 |
| B-9409 | 19 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 |
| B-9414 | 19 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 |
| B-8158 | 45 | 97.65 | 25 | 1 | 98.38 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9378 | 45 | 97.65 | 25 | 1 | 98.36 | 35 | 3 | 98.89 | 24 | 4 | 97.85 | 25 | 6 |
| B-9390 | 78 | 98.03 | 21 | 2 | 98.6 | 30 | 1 | 98.75 | 27 | 4 | pseudo | - | - |
| B-9408 | 78 | 98.03 | 21 | 2 | 98.6 | 30 | 1 | 98.75 | 27 | 4 | pseudo | - | - |
| B-9413 | 78 | 98.03 | 21 | 2 | 98.6 | 30 | 1 | 98.75 | 27 | 4 | pseudo | - | - |
| B-9404 | 106 | pseudo | - | - | 98.83 | 25 | 1 | 98.56 | 31 | 4 | 97.85 | 25 | 6 |
| B-9406 | 400 | 97.84 | 23 | 2 | 98.88 | 24 | 2 | 98.79 | 26 | 2 | 97.85 | 25 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fursova, N.K.; Fursov, M.V.; Astashkin, E.I.; Fursova, A.D.; Novikova, T.S.; Kislichkina, A.A.; Sizova, A.A.; Fedyukina, G.N.; Savin, I.A.; Ershova, O.N. Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Causing Nosocomial Meningitis in the Neurological Intensive Care Unit. Microorganisms 2023, 11, 2020. https://doi.org/10.3390/microorganisms11082020
Fursova NK, Fursov MV, Astashkin EI, Fursova AD, Novikova TS, Kislichkina AA, Sizova AA, Fedyukina GN, Savin IA, Ershova ON. Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Causing Nosocomial Meningitis in the Neurological Intensive Care Unit. Microorganisms. 2023; 11(8):2020. https://doi.org/10.3390/microorganisms11082020
Chicago/Turabian StyleFursova, Nadezhda K., Mikhail V. Fursov, Evgeny I. Astashkin, Anastasiia D. Fursova, Tatiana S. Novikova, Angelina A. Kislichkina, Angelika A. Sizova, Galina N. Fedyukina, Ivan A. Savin, and Olga N. Ershova. 2023. "Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Causing Nosocomial Meningitis in the Neurological Intensive Care Unit" Microorganisms 11, no. 8: 2020. https://doi.org/10.3390/microorganisms11082020