
Desulfovibrio in the Gut: The Enemy within?

Abstract
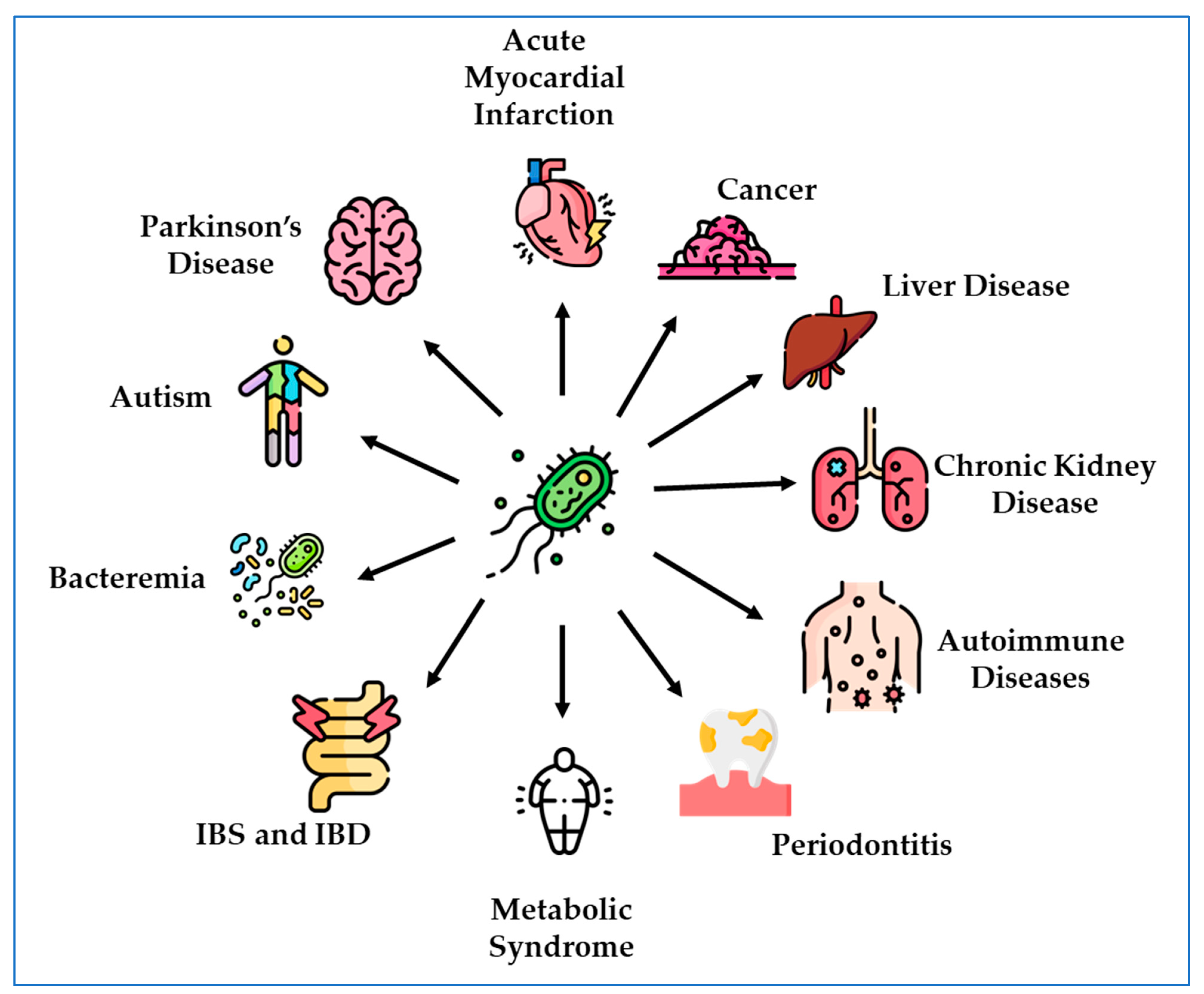
:1. Introduction
2. DSV and Diseases
2.1. Bacteremia
2.2. Intestinal Bacterial Overgrowth
2.2.1. Inflammatory Bowel Disease (IBD)
2.2.2. Neurodegenerative Diseases
2.2.3. Autism
2.2.4. Cognition
2.2.5. Cancer
2.2.6. Metabolic Syndrome

{kind=link}
{kind=link}
{kind=link}
| Disease Context | System | DSV Source | Methods | Findings | Reference |
|---|---|---|---|---|---|
| Parkinson’s Disease | C. elegans | DSV isolates from feces of PD patients | Worms expressing α–syn-YFP fed on DSV containing medium | Alpha synuclein aggregation in brain | [57] |
| Inflammatory Bowel Disease | C57/BL6 mice | D. vulgaris Hildenborough (ATCC | Intestinal transit of a fluorescent probe through the small intestine in animals gavaged with DSV | Intestinal transit was slowed down | [51] |
| Experimental Colitis | C57BL/6 mice (germ free and wild type) | D. indonesiensis isolated from biofilm on corroded ship or human SRB consortium from patients with colitis | H&E staining Cytokine analysis by CBA Th1/Th2/Th17 kit | H&E staining Cytokine analysis by CBA Th1/Th2/Th17 kit | [52] |
| Atherosclerosis | C57/Bl6 Apoe−/− Caco2 | D. desulfuricans | Intestinal permeability using FITC probe Inflammatory markers and tight junction proteins | Increased formation of atherosclerotic lesion Increased inflammation Increased intestinal permeability | [53] |
| Colorectal Cancer | BALB/c Mice | Desulfovibrio (species unspecified), from China General Microbiological Culture Collection Center | Real-Time qPCR ELISA kits for LPS, H2S. | Decreased mRNA for tight junction proteins Increased mRNA for inflammatory markers Increased mRNA levels of extacellular matrix proteins important for formation of pre-metastatic niche (PMN) in the liver Increased serum ALT and AST Increased H2S and LPS in serum | [109] |
| Obesity | C57Bl/6 Myd88LoxP/LoxP mice crossed to C57Bl/6 CD4-Cre animals to produceCD4-Cre+ (T-MyD88−/−) animals | D. desulfuricans subsp. desulfuricans | qPCR 16s rDNA sequencing | Expression of CD63 was increased by DSV Reduction in Clostridia | [145] |
| Cognition | C57/BL6 mice | D. vulgaris Hildenborough (ATCC) | 8-Arm radial learning maze performance Morris water maze performance H2S measurement | Impaired working memory Increased H2S in small intestine and cecum | [95] |
2.2.7. Other Diseases
3. Effect of DSV on Host Cells In Vitro
| Cell Lines | DSV Source | Methods | Findings | Ref. |
|---|---|---|---|---|
| HCT116 | D. indonesiensis mono-culture or co-culture with E. coli isolate 2R/BP SRB consortia from human biopsy samples | Flow cytometry and tunnel labeling for apoptosis Immunostaining | Induction of apoptosis antibody against exopolysaccharides of D. indonesiensis cross reacted with the SRB from UC patients but not with the SRB from non-UC controls. | [159] |
| KB cell line ATCC CCL-17 | D. desulfuricans ATCC 29577, D. desulfuricans ATCC 27774, D. fairfieldensis ATCC 700045 | Invasion assay Electron microscopy ELISA kit for cytokines | DSV invaded KB cells in microtubule dependent manner DSV are present in the free space in cytoplasm Induction of pro-inflammatory cytokines by DSV | [166] |
| RAW 264.7 | Desulfovibrio vulgaris Hildenborough (ATCC 29579) | Western blot siRNA transfection qPCR | Increased mRNA and protein expression of Notch1 and IL-1b. Activation of Notch intracellular domain Paracrine activation of Notch signaling in recipient cells by soluble factors in culture supernatant of DSV-treated cell. | [167] |
| RAW 264.7 | Desulfovibrio vulgaris Hildenborough (ATCC 29579) | Colorimetric assay for nitrite production | Increased nitrite production in D. vulgaris-infected macrophages | [168] |
| Polarized and differentiated Caco2 | Desulfovibrio vulgaris Hildenborough (ATCC 29579) | FITC flux to measure barrier permeability siRNA transfection Western blot | Increased paracellular permeability Increased snail protein expression | [169] |
| RAW 264.7 | Desulfovibrio vulgaris Hildenborough (ATCC 29579) | Western blot Lysozyme activity assay | Decreased lysozyme mRNA and protein expression | [170] |
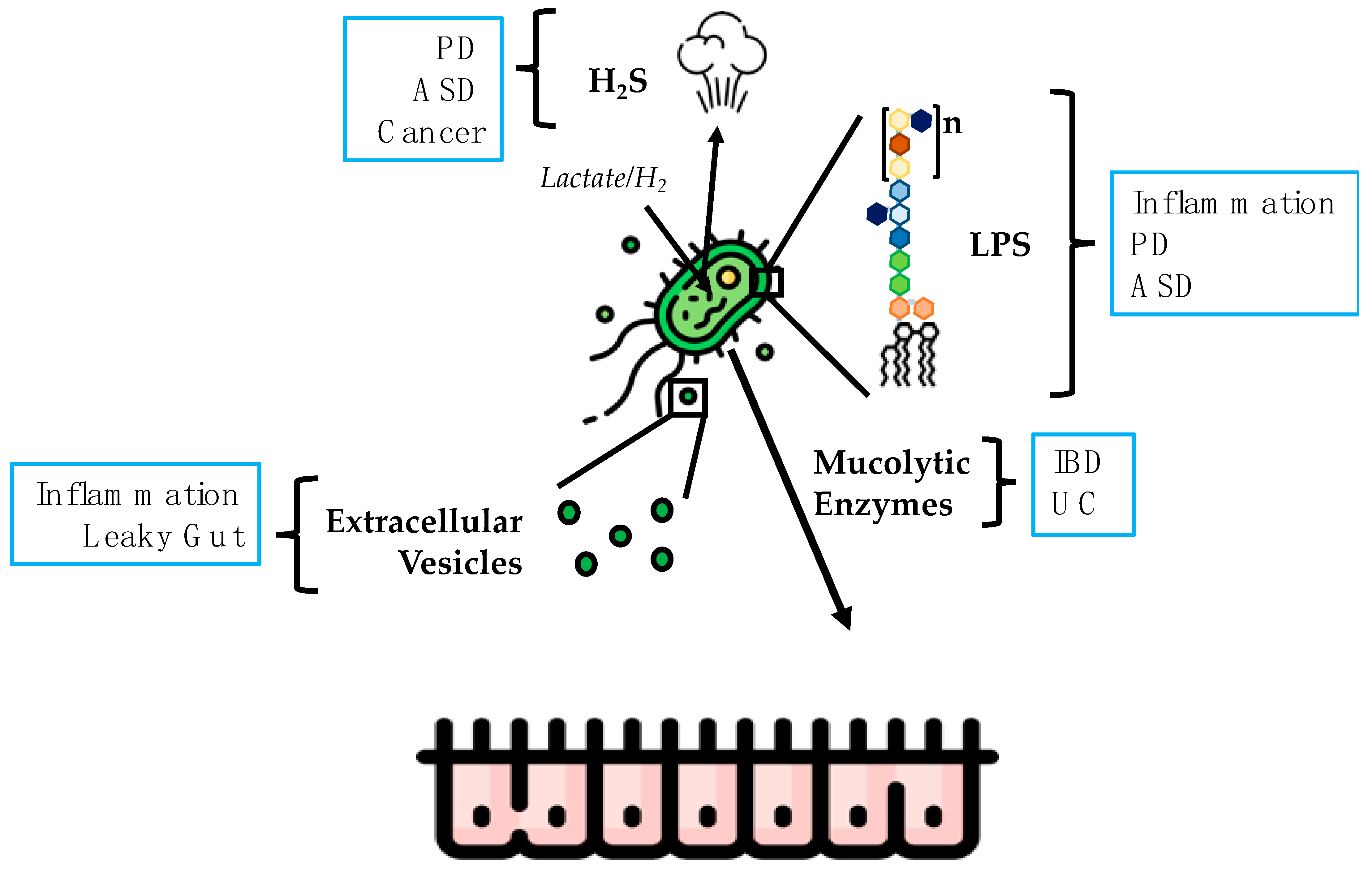
4. Products of DSV Responsible for Causing Potentially Harmful Effects
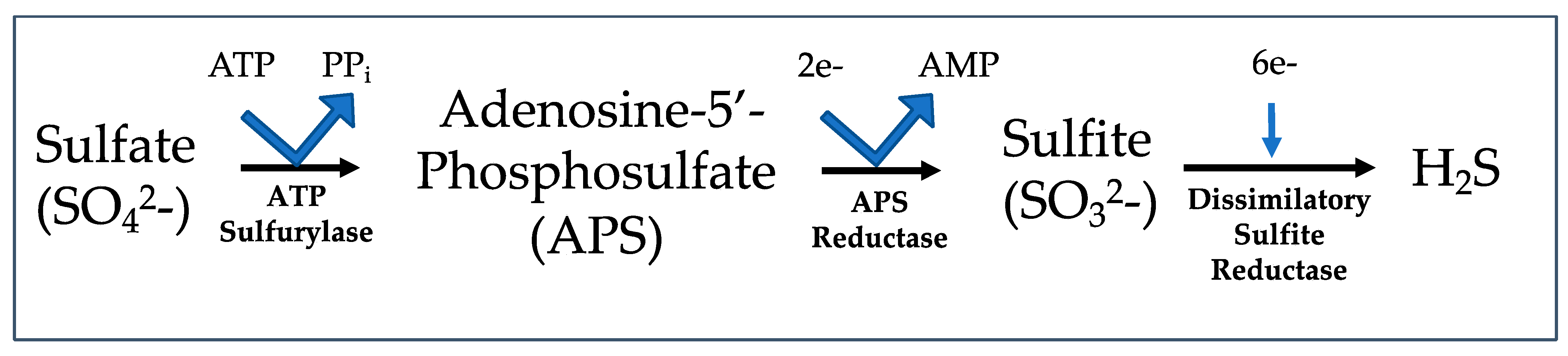
4.1. Hydrogen Sulfide
4.2. LPS
4.3. Extracellular Vesicles
4.4. Mucolytic Activity of DSV
5. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Muyzer, G.; Stams, A.J. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef]
- Jørgensen, B.B. Mineralization of organic matter in the sea bed—The role of sulphate reduction. Nature 1982, 296, 643–645. [Google Scholar] [CrossRef]
- Mckenzie, J.A.; Vasconcelos, C. Dolomite Mountains and the origin of the dolomite rock of which they mainly consist: Historical developments and new perspectives. Sedimentology 2009, 56, 205–219. [Google Scholar] [CrossRef]
- Schwartz, W.; Postgate, J.R. The Sulfate-Reducing Bacteria (2nd Edition) X + 208 S., 20 Abb., 4 Tab. University Press, Cambridge 1983. US $ 39.50. J. Basic Microbiol. 1985, 25, 202. [Google Scholar] [CrossRef]
- Loubinoux, J.; Valente, F.M.; Pereira, I.A.; Costa, A.; Grimont, P.A.; Le Faou, A.E. Reclassification of the only species of the genus Desulfomonas, Desulfomonas pigra, as Desulfovibrio piger comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1305–1308. [Google Scholar] [PubMed] [Green Version]
- Rey, F.E.; Gonzalez, M.D.; Cheng, J.; Wu, M.; Ahern, P.P.; Gordon, J.I. Metabolic niche of a prominent sulfate-reducing human gut bacterium. Proc. Natl. Acad. Sci. USA 2013, 110, 13582–13587. [Google Scholar] [CrossRef]
- Barton, L.L.; Fauque, G.D. Biochemistry, physiology and biotechnology of sulfate-reducing bacteria. Adv. Appl. Microbiol. 2009, 68, 41–98. [Google Scholar]
- Dordevic, D.; Jancikova, S.; Vitezova, M.; Kushkevych, I. Hydrogen sulfide toxicity in the gut environment: Meta-analysis of sulfate-reducing and lactic acid bacteria in inflammatory processes. J. Adv. Res. 2021, 27, 55–69. [Google Scholar] [CrossRef]
- Kushkevych, I.; Cejnar, J.; Treml, J.; Dordevic, D.; Kollar, P.; Vitezova, M. Recent Advances in Metabolic Pathways of Sulfate Reduction in Intestinal Bacteria. Cells 2020, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.W.; Zhu, J.; Zuo, S.; Zhang, J.L.; Chen, Z.Y.; Chen, G.W.; Wang, X.; Pan, Y.S.; Liu, Y.C.; Wang, P.Y. Protective effect of hydrogen sulfide on TNF-alpha and IFN-gamma-induced injury of intestinal epithelial barrier function in Caco-2 monolayers. Inflamm. Res. 2015, 64, 789–797. [Google Scholar] [CrossRef]
- Liu, Y.; Kalogeris, T.; Wang, M.; Zuidema, M.Y.; Wang, Q.; Dai, H.; Davis, M.J.; Hill, M.A.; Korthuis, R.J. Hydrogen sulfide preconditioning or neutrophil depletion attenuates ischemia-reperfusion-induced mitochondrial dysfunction in rat small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G44–G54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yan, R.; Zhou, X.; Ji, F.; Zhang, B. Hydrogen sulfide improves colonic barrier integrity in DSS-induced inflammation in Caco-2 cells and mice. Int. Immunopharmacol. 2016, 39, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, E.J.; Citron, D.M.; Peraino, V.A.; Cross, S.A. Desulfovibrio desulfuricans bacteremia and review of human Desulfovibrio infections. J. Clin. Microbiol. 2003, 41, 2752–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machaca, M.; Bodean, M.L.; Montana, S.; Garcia, S.D.; Stecher, D.; Vay, C.A.; Almuzara, M.N. Description of a case of abdominal sepsis due to Desulfovibrio desulfuricans. Rev. Argent. Microbiol. 2022, 54, 314–317. [Google Scholar] [PubMed]
- Marquis, T.J.; Williams, V.J.; Banach, D.B. Septic arthritis caused by Desulfovibrio desulfuricans: A case report and review of the literature. Anaerobe 2021, 70, 102407. [Google Scholar] [CrossRef]
- Liderot, K.; Larsson, M.; Borang, S.; Ozenci, V. Polymicrobial bloodstream infection with Eggerthella lenta and Desulfovibrio desulfuricans. J. Clin. Microbiol. 2010, 48, 3810–3812. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, Y.; Miyabe, Y.; Seki, M.; Ushio, Y.; Sato, K.; Kasama, E.; Akiyama, K.; Karasawa, K.; Uchida, K.; Kikuchi, K.; et al. First case of a renal cyst infection caused by Desulfovibrio: A case report and literature review. BMC Nephrol. 2022, 23, 194. [Google Scholar] [CrossRef]
- Chesdachai, S.; Eberly, A.R.; Razonable, R.R. A tale of two unusual anaerobic bacterial infections in an immunocompetent man: A case report and literature review. Anaerobe 2021, 71, 102416. [Google Scholar] [CrossRef]
- Predari, S.C.; de Paulis, A.N.; Bertona, E.; Guevara Nunez, D.; Suarez, J.P.; Castello, L. Anaerobiospirillum succiniciproducens and Desulfovibrio desulfuricans in 2 cases of insidious bacteremia. Rev. Argent. Microbiol. 2017, 49, 146–152. [Google Scholar]
- Urata, T.; Kikuchi, M.; Hino, T.; Yoda, Y.; Tamai, K.; Kodaira, Y.; Hitomi, S. Bacteremia caused by Desulfovibrio fairfieldensis. J. Infect. Chemother. 2008, 14, 368–370. [Google Scholar] [CrossRef]
- Hagiya, H.; Kimura, K.; Nishi, I.; Yamamoto, N.; Yoshida, H.; Akeda, Y.; Tomono, K. Desulfovibrio desulfuricans bacteremia: A case report and literature review. Anaerobe 2018, 49, 112–115. [Google Scholar] [CrossRef]
- Gaillard, T.; Pons, S.; Darles, C.; Beausset, O.; Monchal, T.; Brisou, P. Desulfovibrio fairfieldensis bacteremia associated with acute sigmoiditis. Med. Mal. Infect. 2011, 41, 267–268. [Google Scholar] [CrossRef]
- Koyano, S.; Tatsuno, K.; Okazaki, M.; Ohkusu, K.; Sasaki, T.; Saito, R.; Okugawa, S.; Moriya, K. A Case of Liver Abscess with Desulfovibrio desulfuricans Bacteremia. Case Rep. Infect. Dis. 2015, 2015, 354168. [Google Scholar] [PubMed] [Green Version]
- Hagiwara, S.; Yoshida, A.; Omata, Y.; Tsukada, Y.; Takahashi, H.; Kamewada, H.; Koike, S.; Okuzumi, K.; Hishinuma, A.; Kobayashi, K.; et al. Desulfovibrio desulfuricans bacteremia in a patient hospitalized with acute cerebral infarction: Case report and review. J. Infect. Chemother. 2014, 20, 274–277. [Google Scholar] [CrossRef]
- Pimentel, J.D.; Chan, R.C. Desulfovibrio fairfieldensis bacteremia associated with choledocholithiasis and endoscopic retrograde cholangiopancreatography. J. Clin. Microbiol. 2007, 45, 2747–2750. [Google Scholar] [CrossRef] [Green Version]
- Verstreken, I.; Laleman, W.; Wauters, G.; Verhaegen, J. Desulfovibrio desulfuricans bacteremia in an immunocompromised host with a liver graft and ulcerative colitis. J. Clin. Microbiol. 2012, 50, 199–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weglarz, L.; Dzierzewicz, Z.; Skop, B.; Orchel, A.; Parfiniewicz, B.; Wisniowska, B.; Swiatkowska, L.; Wilczok, T. Desulfovibrio desulfuricans lipopolysaccharides induce endothelial cell IL-6 and IL-8 secretion and E-selectin and VCAM-1 expression. Cell Mol. Biol. Lett. 2003, 8, 991–1003. [Google Scholar]
- Kapral, M.; Weglarz, L.; Parfiniewicz, B.; Lodowska, J.; Jaworska-Kik, M. Quantitative evaluation of transcriptional activation of NF-kappaB p65 and p50 subunits and IkappaBalpha encoding genes in colon cancer cells by Desulfovibrio desulfuricans endotoxin. Folia Microbiol. 2010, 55, 657–661. [Google Scholar] [CrossRef]
- Dzierzewicz, Z.; Szczerba, J.; Lodowska, J.; Wolny, D.; Gruchlik, A.; Orchel, A.; Weglarz, L. The role of Desulfovibrio desulfuricans lipopolysaccharides in modulation of periodontal inflammation through stimulation of human gingival fibroblasts. Arch. Oral. Biol. 2010, 55, 515–522. [Google Scholar] [CrossRef]
- Nie, Y.; Xie, X.Q.; Zhou, L.; Guan, Q.; Ren, Y.; Mao, Y.; Shi, J.S.; Xu, Z.H.; Geng, Y. Desulfovibrio fairfieldensis-Derived Outer Membrane Vesicles Damage Epithelial Barrier and Induce Inflammation and Pyroptosis in Macrophages. Cells 2022, 12, 89. [Google Scholar] [CrossRef]
- Collaborators, G.B.D.I.B.D. The global, regional, and national burden of inflammatory bowel disease in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [Google Scholar]
- Haneishi, Y.; Furuya, Y.; Hasegawa, M.; Picarelli, A.; Rossi, M.; Miyamoto, J. Inflammatory Bowel Diseases and Gut Microbiota. Int. J. Mol. Sci. 2023, 24, 3817. [Google Scholar] [CrossRef] [PubMed]
- Rowan, F.; Docherty, N.G.; Murphy, M.; Murphy, B.; Calvin Coffey, J.; O’Connell, P.R. Desulfovibrio bacterial species are increased in ulcerative colitis. Dis. Colon. Rectum 2010, 53, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Loubinoux, J.; Bronowicki, J.P.; Pereira, I.A.; Mougenel, J.L.; Faou, A.E. Sulfate-reducing bacteria in human feces and their association with inflammatory bowel diseases. FEMS Microbiol. Ecol. 2002, 40, 107–112. [Google Scholar] [CrossRef]
- Fite, A.; Macfarlane, S.; Furrie, E.; Bahrami, B.; Cummings, J.H.; Steinke, D.T.; Macfarlane, G.T. Longitudinal analyses of gut mucosal microbiotas in ulcerative colitis in relation to patient age and disease severity and duration. J. Clin. Microbiol. 2013, 51, 849–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humbel, F.; Rieder, J.H.; Franc, Y.; Juillerat, P.; Scharl, M.; Misselwitz, B.; Schreiner, P.; Begre, S.; Rogler, G.; von Kanel, R.; et al. Association of Alterations in Intestinal Microbiota With Impaired Psychological Function in Patients With Inflammatory Bowel Diseases in Remission. Clin. Gastroenterol. Hepatol. 2020, 18, 2019–2029.e11. [Google Scholar] [CrossRef]
- Sun, Y.; Xie, R.; Li, L.; Jin, G.; Zhou, B.; Huang, H.; Li, M.; Yang, Y.; Liu, X.; Cao, X.; et al. Prenatal Maternal Stress Exacerbates Experimental Colitis of Offspring in Adulthood. Front. Immunol. 2021, 12, 700995. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Dong, N.; Fang, Q.; Zhang, Y.; Chen, C.; Cui, S.W.; Nie, S. Polysaccharides from natural Cordyceps sinensis attenuated dextran sodium sulfate-induced colitis in C57BL/6J mice. Food Funct. 2023, 14, 720–733. [Google Scholar] [CrossRef]
- Ye, X.; Cen, Y.; Wu, K.; Xu, L.; Ni, J.; Zheng, W.; Liu, W. Gas-Mediated Intestinal Microbiome Regulation Prompts the Methanol Extract of Schizonepetae Spica to Relieve Colitis. Nutrients 2023, 15, 519. [Google Scholar] [CrossRef]
- Wan, Y.; Yang, L.; Li, H.; Ren, H.; Zhu, K.; Dong, Z.; Jiang, S.; Shang, E.; Qian, D.; Duan, J. Zingiber officinale and Panax ginseng ameliorate ulcerative colitis in mice via modulating gut microbiota and its metabolites. J. Chromatogr. B Analyt Technol. Biomed. Life Sci. 2022, 1203, 123313. [Google Scholar] [CrossRef]
- Bi, Z.; Cui, E.; Yao, Y.; Chang, X.; Wang, X.; Zhang, Y.; Xu, G.X.; Zhuang, H.; Hua, Z.C. Recombinant Bifidobacterium longum Carrying Endostatin Protein Alleviates Dextran Sodium Sulfate-Induced Colitis and Colon Cancer in Rats. Front. Microbiol. 2022, 13, 927277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.X.; Yuan, X.; Cui, Y.Y.; Liu, J.; Shen, J.; Jin, B.Y.; Feng, B.C.; Zhai, Y.J.; Zheng, M.Q.; Kou, G.J.; et al. Melatonin Mitigates Oxazolone-Induced Colitis in Microbiota-Dependent Manner. Front. Immunol. 2021, 12, 783806. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Yue, Y.; Guan, Q.; Ren, Y.; Guo, L.; Fan, Y.; Lu, Z.M.; Shi, J.S.; Xu, Z.H. Cereal Vinegar Sediment Alleviates Spontaneous Ulcerative Colitis in Il-10 Deficient Mice. Mol. Nutr. Food Res. 2021, 65, e2001227. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, G.; Yang, Q.; Ye, J.; Cai, X.; Tsering, P.; Cheng, X.; Hu, C.; Zhang, S.; Cao, P. Gut microbiota drives the attenuation of dextran sulphate sodium-induced colitis by Huangqin decoction. Oncotarget 2017, 8, 48863–48874. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Peng, C.; Li, J.; Cheng, R.; He, F.; Yang, L.; Lu, J.; Shen, X. Protective effect and mechanism of Bifidobacterium bifidum TMC3115 on long-term colitis in mice which exposed to antibiotic in early life. Wei Sheng Yan Jiu 2022, 51, 624–644. [Google Scholar]
- Song, H.; Wang, W.; Shen, B.; Jia, H.; Hou, Z.; Chen, P.; Sun, Y. Pretreatment with probiotic Bifico ameliorates colitis-associated cancer in mice: Transcriptome and gut flora profiling. Cancer Sci. 2018, 109, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Pitcher, M.C.; Beatty, E.R.; Cummings, J.H. The contribution of sulphate reducing bacteria and 5-aminosalicylic acid to faecal sulphide in patients with ulcerative colitis. Gut 2000, 46, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Zinkevich, V.V.; Beech, I.B. Screening of sulfate-reducing bacteria in colonoscopy samples from healthy and colitic human gut mucosa. FEMS Microbiol. Ecol. 2000, 34, 147–155. [Google Scholar] [CrossRef]
- Fischer, M.; Siva, S.; Wo, J.M.; Fadda, H.M. Assessment of Small Intestinal Transit Times in Ulcerative Colitis and Crohn’s Disease Patients with Different Disease Activity Using Video Capsule Endoscopy. AAPS PharmSciTech 2017, 18, 404–409. [Google Scholar] [CrossRef]
- Buret, A.G.; Allain, T.; Motta, J.P.; Wallace, J.L. Effects of Hydrogen Sulfide on the Microbiome: From Toxicity to Therapy. Antioxid. Redox Signal. 2022, 36, 211–219. [Google Scholar] [CrossRef]
- Ritz, N.L.; Lin, D.M.; Wilson, M.R.; Barton, L.L.; Lin, H.C. Sulfate-reducing bacteria slow intestinal transit in a bismuth-reversible fashion in mice. Neurogastroenterol. Motil. 2017, 29, e12907. [Google Scholar] [CrossRef] [PubMed]
- Figliuolo, V.R.; Dos Santos, L.M.; Abalo, A.; Nanini, H.; Santos, A.; Brittes, N.M.; Bernardazzi, C.; de Souza, H.S.P.; Vieira, L.Q.; Coutinho-Silva, R.; et al. Sulfate-reducing bacteria stimulate gut immune responses and contribute to inflammation in experimental colitis. Life Sci. 2017, 189, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Qin, X.; Qiu, J.; Sun, T.; Qu, K.; Din, A.U.; Yan, W.; Li, T.; Chen, Y.; Gu, W.; et al. Desulfovibrio desulfuricans aggravates atherosclerosis by enhancing intestinal permeability and endothelial TLR4/NF-kappaB pathway in Apoe (−/−) mice. Genes. Dis. 2023, 10, 239–253. [Google Scholar] [CrossRef]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Ryman, S.; Vakhtin, A.A.; Richardson, S.P.; Lin, H.C. Microbiome-gut-brain dysfunction in prodromal and symptomatic Lewy body diseases. J. Neurol. 2023, 270, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liang, H.; Hu, Y.; Lu, L.; Zheng, C.; Fan, Y.; Wu, B.; Zou, T.; Luo, X.; Zhang, X.; et al. Gut bacterial profiles in Parkinson’s disease: A systematic review. CNS Neurosci. Ther. 2023, 29, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Huynh, V.A.; Takala, T.M.; Murros, K.E.; Diwedi, B.; Saris, P.E.J. Desulfovibrio bacteria enhance alpha-synuclein aggregation in a Caenorhabditis elegans model of Parkinson’s disease. Front. Cell Infect. Microbiol. 2023, 13, 1181315. [Google Scholar] [CrossRef]
- Murros, K.E.; Huynh, V.A.; Takala, T.M.; Saris, P.E.J. Desulfovibrio Bacteria Are Associated with Parkinson’s Disease. Front. Cell Infect. Microbiol. 2021, 11, 652617. [Google Scholar] [CrossRef]
- Murros, K.E. Hydrogen Sulfide Produced by Gut Bacteria May Induce Parkinson’s Disease. Cells 2022, 11, 978. [Google Scholar] [CrossRef]
- Bayir, H.; Kapralov, A.A.; Jiang, J.; Huang, Z.; Tyurina, Y.Y.; Tyurin, V.A.; Zhao, Q.; Belikova, N.A.; Vlasova, I.I.; Maeda, A.; et al. Peroxidase mechanism of lipid-dependent cross-linking of synuclein with cytochrome C: Protection against apoptosis versus delayed oxidative stress in Parkinson disease. J. Biol. Chem. 2009, 284, 15951–15969. [Google Scholar] [CrossRef] [Green Version]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Carboni, E.; Lingor, P. Insights on the interaction of alpha-synuclein and metals in the pathophysiology of Parkinson’s disease. Metallomics 2015, 7, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Joppe, K.; Roser, A.E.; Maass, F.; Lingor, P. The Contribution of Iron to Protein Aggregation Disorders in the Central Nervous System. Front. Neurosci. 2019, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Devos, D.; Cabantchik, Z.I.; Moreau, C.; Danel, V.; Mahoney-Sanchez, L.; Bouchaoui, H.; Gouel, F.; Rolland, A.S.; Duce, J.A.; Devedjian, J.C.; et al. Conservative iron chelation for neurodegenerative diseases such as Parkinson’s disease and amyotrophic lateral sclerosis. J. Neural Transm. 2020, 127, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.; Basak, S.; Kundu, S.; De, G.; Mukhopadhyay, A.; Chattopadhyay, K. Attenuation of the early events of alpha-synuclein aggregation: A fluorescence correlation spectroscopy and laser scanning microscopy study in the presence of surface-coated Fe3O4 nanoparticles. Langmuir 2015, 31, 1469–1478. [Google Scholar] [CrossRef]
- Pereira, I.A.; Ramos, A.R.; Grein, F.; Marques, M.C.; da Silva, S.M.; Venceslau, S.S. A comparative genomic analysis of energy metabolism in sulfate reducing bacteria and archaea. Front. Microbiol. 2011, 2, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murros, K.; Wasiljeff, J.; Macias-Sanchez, E.; Faivre, D.; Soinne, L.; Valtonen, J.; Pohja, M.; Saari, P.; Pesonen, L.J.; Salminen, J.M. Magnetic Nanoparticles in Human Cervical Skin. Front. Med. 2019, 6, 123. [Google Scholar] [CrossRef]
- Hertel, J.; Harms, A.C.; Heinken, A.; Baldini, F.; Thinnes, C.C.; Glaab, E.; Vasco, D.A.; Pietzner, M.; Stewart, I.D.; Wareham, N.J.; et al. Integrated Analyses of Microbiome and Longitudinal Metabolome Data Reveal Microbial-Host Interactions on Sulfur Metabolism in Parkinson’s Disease. Cell Rep. 2019, 29, 1767–1777.e1768. [Google Scholar] [CrossRef] [Green Version]
- Licking, N.; Murchison, C.; Cholerton, B.; Zabetian, C.P.; Hu, S.C.; Montine, T.J.; Peterson-Hiller, A.L.; Chung, K.A.; Edwards, K.; Leverenz, J.B.; et al. Homocysteine and cognitive function in Parkinson’s disease. Park. Relat. Disord. 2017, 44, 1–5. [Google Scholar] [CrossRef]
- Miller, J.W. Homocysteine and Alzheimer’s disease. Nutr. Rev. 1999, 57, 126–129. [Google Scholar]
- Kumar, M.; Sandhir, R. Hydrogen sulfide attenuates hyperhomocysteinemia-induced mitochondrial dysfunctions in brain. Mitochondrion 2020, 50, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ning, J.; Bao, X.Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. A New Concept of Associations between Gut Microbiota, Immunity and Central Nervous System for the Innovative Treatment of Neurodegenerative Disorders. Metabolites 2022, 12, 1052. [Google Scholar] [CrossRef] [PubMed]
- Susmitha, G.; Kumar, R. Role of microbial dysbiosis in the pathogenesis of Alzheimer’s disease. Neuropharmacology 2023, 229, 109478. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Dawson, T.M.; Kulkarni, S. Neurodegenerative disorders and gut-brain interactions. J. Clin. Investig. 2021, 131, e143775. [Google Scholar] [CrossRef]
- Finegold, S.M. Desulfovibrio species are potentially important in regressive autism. Med. Hypotheses 2011, 77, 270–274. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Wu, F.; Zheng, H.; Peng, Q.; Zhou, H. Altered composition and function of intestinal microbiota in autism spectrum disorders: A systematic review. Transl. Psychiatry 2019, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef] [Green Version]
- Finegold, S.M.; Dowd, S.E.; Gontcharova, V.; Liu, C.; Henley, K.E.; Wolcott, R.D.; Youn, E.; Summanen, P.H.; Granpeesheh, D.; Dixon, D.; et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 2010, 16, 444–453. [Google Scholar] [CrossRef]
- Tomova, A.; Husarova, V.; Lakatosova, S.; Bakos, J.; Vlkova, B.; Babinska, K.; Ostatnikova, D. Gastrointestinal microbiota in children with autism in Slovakia. Physiol. Behav. 2015, 138, 179–187. [Google Scholar] [CrossRef]
- Karnachuk, O.V.; Ikkert, O.P.; Avakyan, M.R.; Knyazev, Y.V.; NVolochaev, M.; Zyusman, V.S.; Panov, V.L.; Kadnikov, V.V.; Mardanov, A.V.; Ravin, N.V. Desulfovibrio desulfuricans AY5 Isolated from a Patient with Autism Spectrum Disorder Binds Iron in Low-Soluble Greigite and Pyrite. Microorganisms 2021, 9, 2558. [Google Scholar] [CrossRef]
- Sidrak, S.; Yoong, T.; Woolfenden, S. Iron deficiency in children with global developmental delay and autism spectrum disorder. J. Paediatr. Child. Health 2014, 50, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Yanagimoto, Y.; Ishizaki, Y.; Kaneko, K. Iron deficiency anemia, stunted growth, and developmental delay due to avoidant/restrictive food intake disorder by restricted eating in autism spectrum disorder. Biopsychosoc. Med. 2020, 14, 8. [Google Scholar] [CrossRef]
- Emanuele, E.; Orsi, P.; Boso, M.; Broglia, D.; Brondino, N.; Barale, F.; di Nemi, S.U.; Politi, P. Low-grade endotoxemia in patients with severe autism. Neurosci. Lett. 2010, 471, 162–165. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Usui, N.; Kobayashi, H.; Shimada, S. Neuroinflammation and Oxidative Stress in the Pathogenesis of Autism Spectrum Disorder. Int. J. Mol. Sci. 2023, 24, 5487. [Google Scholar] [CrossRef] [PubMed]
- Nogay, N.H.; Nahikian-Nelms, M. Can we reduce autism-related gastrointestinal and behavior problems by gut microbiota based dietary modulation? A review. Nutr. Neurosci. 2021, 24, 327–338. [Google Scholar] [CrossRef]
- Thomas, R.H.; Meeking, M.M.; Mepham, J.R.; Tichenoff, L.; Possmayer, F.; Liu, S.; MacFabe, D.F. The enteric bacterial metabolite propionic acid alters brain and plasma phospholipid molecular species: Further development of a rodent model of autism spectrum disorders. J. Neuroinflam. 2012, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, K.A.; MacFabe, D.F.; Vaz, A.; Ossenkopp, K.P.; Kavaliers, M. Sexually dimorphic effects of prenatal exposure to propionic acid and lipopolysaccharide on social behavior in neonatal, adolescent, and adult rats: Implications for autism spectrum disorders. Int. J. Dev. Neurosci. 2014, 39, 68–78. [Google Scholar] [CrossRef]
- MacFabe, D.F. Enteric short-chain fatty acids: Microbial messengers of metabolism, mitochondria, and mind: Implications in autism spectrum disorders. Microb. Ecol. Health Dis. 2015, 26, 28177. [Google Scholar] [CrossRef] [Green Version]
- MacFabe, D.F.; Cain, N.E.; Boon, F.; Ossenkopp, K.P.; Cain, D.P. Effects of the enteric bacterial metabolic product propionic acid on object-directed behavior, social behavior, cognition, and neuroinflammation in adolescent rats: Relevance to autism spectrum disorder. Behav. Brain Res. 2011, 217, 47–54. [Google Scholar] [CrossRef]
- MacFabe, D.F.; Cain, D.P.; Rodriguez-Capote, K.; Franklin, A.E.; Hoffman, J.E.; Boon, F.; Taylor, A.R.; Kavaliers, M.; Ossenkopp, K.P. Neurobiological effects of intraventricular propionic acid in rats: Possible role of short chain fatty acids on the pathogenesis and characteristics of autism spectrum disorders. Behav. Brain Res. 2007, 176, 149–169. [Google Scholar] [CrossRef]
- Bell, J.G.; MacKinlay, E.E.; Dick, J.R.; MacDonald, D.J.; Boyle, R.M.; Glen, A.C. Essential fatty acids and phospholipase A2 in autistic spectrum disorders. Prostaglandins Leukot. Essent. Fat. Acids 2004, 71, 201–204. [Google Scholar] [CrossRef]
- Kang, D.W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome 2017, 5, 10. [Google Scholar] [CrossRef]
- Ritz, N.L.; Burnett, B.J.; Setty, P.; Reinhart, K.M.; Wilson, M.R.; Alcock, J.; Singh, S.B.; Barton, L.L.; Lin, H.C. Sulfate-reducing bacteria impairs working memory in mice. Physiol. Behav. 2016, 157, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Lin, H.C. Hydrogen Sulfide in Physiology and Diseases of the Digestive Tract. Microorganisms 2015, 3, 866–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilburn, K.H.; Warshaw, R.H. Hydrogen sulfide and reduced-sulfur gases adversely affect neurophysiological functions. Toxicol. Ind. Health 1995, 11, 185–197. [Google Scholar] [CrossRef]
- Tvedt, B.; Edland, A.; Skyberg, K.; Forberg, O. Delayed neuropsychiatric sequelae after acute hydrogen sulfide poisoning: Affection of motor function, memory, vision and hearing. Acta Neurol. Scand. 1991, 84, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Partlo, L.A.; Sainsbury, R.S.; Roth, S.H. Effects of repeated hydrogen sulphide (H2S) exposure on learning and memory in the adult rat. Neurotoxicology 2001, 22, 177–189. [Google Scholar] [CrossRef]
- Kilburn, K.H. Case report: Profound neurobehavioral deficits in an oil field worker overcome by hydrogen sulfide. Am. J. Med. Sci. 1993, 306, 301–305. [Google Scholar] [CrossRef]
- Thomas, A.M.; Jesus, E.C.; Lopes, A.; Aguiar, S., Jr.; Begnami, M.D.; Rocha, R.M.; Carpinetti, P.A.; Camargo, A.A.; Hoffmann, C.; Freitas, H.C.; et al. Tissue-Associated Bacterial Alterations in Rectal Carcinoma Patients Revealed by 16S rRNA Community Profiling. Front. Cell Infect. Microbiol. 2016, 6, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, V.L.; Chen, J.; Johnson, S.; Harrington, S.C.; Yab, T.C.; Smyrk, T.C.; Nelson, H.; Boardman, L.A.; Druliner, B.R.; Levin, T.R.; et al. Shifts in the Fecal Microbiota Associated with Adenomatous Polyps. Cancer Epidemiol. Biomark. Prev. 2017, 26, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, Y.; Wang, Z.; Tang, J.; Cao, D.X.; Qian, Y.; Xie, Y.H.; Chen, H.Y.; Chen, Y.X.; Chen, Z.F.; et al. A clinical nomogram incorporating salivary Desulfovibrio desulfuricans level and oral hygiene index for predicting colorectal cancer. Ann. Transl. Med. 2021, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Cai, Y.; Yang, B.; Xie, S.; Shen, W.; Wu, Y.; Sui, Z.; Cai, J.; Ni, C.; Ye, J. High-Fat Diet Enhances the Liver Metastasis Potential of Colorectal Cancer through Microbiota Dysbiosis. Cancers 2022, 14, 2573. [Google Scholar] [CrossRef]
- Kaplan, R.N.; Riba, R.D.; Zacharoulis, S.; Bramley, A.H.; Vincent, L.; Costa, C.; MacDonald, D.D.; Jin, D.K.; Shido, K.; Kerns, S.A.; et al. VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature 2005, 438, 820–827. [Google Scholar] [CrossRef] [Green Version]
- Szabo, C.; Coletta, C.; Chao, C.; Modis, K.; Szczesny, B.; Papapetropoulos, A.; Hellmich, M.R. Tumor-derived hydrogen sulfide, produced by cystathionine-beta-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 12474–12479. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E. Impact of Hydrogen Sulfide on Mitochondrial and Bacterial Bioenergetics. Int. J. Mol. Sci. 2021, 22, 12688. [Google Scholar] [CrossRef]
- Szabo, C. Hydrogen Sulfide, an Endogenous Stimulator of Mitochondrial Function in Cancer Cells. Cells 2021, 10, 220. [Google Scholar] [CrossRef]
- Zhu, Q.; Jin, Z.; Wu, W.; Gao, R.; Guo, B.; Gao, Z.; Yang, Y.; Qin, H. Analysis of the Intestinal Lumen Microbiota in an Animal Model of Colorectal Cancer. PLoS ONE 2014, 9, e90849. [Google Scholar] [CrossRef]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.-X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2020, 70, 761–774. [Google Scholar] [CrossRef]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-time polymerase chain reaction quantification of specific butyrate-producing bacteria, Desulfovibrio and Enterococcus faecalis in the feces of patients with colorectal cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef]
- Scanlan, P.D.; Shanahan, F.; Marchesi, J.R. Culture-independent analysis of desulfovibrios in the human distal colon of healthy, colorectal cancer and polypectomized individuals. FEMS Microbiol. Ecol. 2009, 69, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Z.-W.; Zhao, B.-C.; Yang, X.; Lei, S.-H.; Jiang, Y.-M.; Liu, K.-X. Relationships of sleep disturbance, intestinal microbiota, and postoperative pain in breast cancer patients: A prospective observational study. Sleep Breath. 2020, 25, 1655–1664. [Google Scholar] [CrossRef]
- Cao, W.; Zheng, C.; Xu, X.; Jin, R.; Huang, F.; Shi, M.; He, Z.; Luo, Y.; Liu, L.; Liu, Z.; et al. Clostridium butyricum potentially improves inflammation and immunity through alteration of the microbiota and metabolism of gastric cancer patients after gastrectomy. Front. Immunol. 2022, 13, 1076245. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, L.; Wang, P.; Liu, Y.; Wang, G.; Shan, Y.; Yi, Y.; Zhou, Y.; Liu, B.; Wang, X.; et al. Lactobacillus coryniformis MXJ32 administration ameliorates azoxymethane/dextran sulfate sodium-induced colitis-associated colorectal cancer via reshaping intestinal microenvironment and alleviating inflammatory response. Eur. J. Nutr. 2021, 61, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Olsson, L.M.; Poitou, C.; Tremaroli, V.; Coupaye, M.; Aron-Wisnewsky, J.; Bäckhed, F.; Clément, K.; Caesar, R. Gut microbiota of obese subjects with Prader-Willi syndrome is linked to metabolic health. Gut 2019, 69, 1229–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Doumatey, A.P.; Adeyemo, A.; Zhou, J.; Lei, L.; Adebamowo, S.N.; Adebamowo, C.; Rotimi, C.N. Gut Microbiome Profiles Are Associated With Type 2 Diabetes in Urban Africans. Front. Cell Infect. Microbiol. 2020, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Alcoholado, L.; Castellano-Castillo, D.; Jordan-Martinez, L.; Moreno-Indias, I.; Cardila-Cruz, P.; Elena, D.; Munoz-Garcia, A.J.; Queipo-Ortuno, M.I.; Jimenez-Navarro, M. Role of Gut Microbiota on Cardio-Metabolic Parameters and Immunity in Coronary Artery Disease Patients with and without Type-2 Diabetes Mellitus. Front. Microbiol. 2017, 8, 1936. [Google Scholar] [CrossRef] [Green Version]
- Crusell, M.K.W.; Hansen, T.H.; Nielsen, T.; Allin, K.H.; Ruhlemann, M.C.; Damm, P.; Vestergaard, H.; Rorbye, C.; Jorgensen, N.R.; Christiansen, O.B.; et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome 2018, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, B.; Ren, L.; Du, H.; Fei, C.; Qian, C.; Li, B.; Zhang, R.; Liu, H.; Li, Z.; et al. High-fiber diet ameliorates gut microbiota, serum metabolism and emotional mood in type 2 diabetes patients. Front. Cell Infect. Microbiol. 2023, 13, 1069954. [Google Scholar] [CrossRef]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 10, 5804–5815. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Hong, Z.; Zhou, T.; Jian, Y.; Xu, M.; Zhang, X.; Zhu, X.; Wang, J. Health improvements of type 2 diabetic patients through diet and diet plus fecal microbiota transplantation. Sci. Rep. 2022, 12, 1152. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Xu, K.; Ni, X.; Li, X.; Zhu, X.; Xu, W. Habitual Dietary Fiber Intake, Fecal Microbiota, and Hemoglobin A1c Level in Chinese Patients with Type 2 Diabetes. Nutrients 2022, 14, 1003. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, L.; Zhang, Q.; Zhang, J.; Liu, S.; Li, C.; Wang, L. Glycolipid Metabolism and Metagenomic Analysis of the Therapeutic Effect of a Phenolics-Rich Extract from Noni Fruit on Type 2 Diabetic Mice. J. Agric. Food Chem. 2022, 70, 2876–2888. [Google Scholar] [CrossRef]
- Hong, Y.; Sheng, L.; Zhong, J.; Tao, X.; Zhu, W.; Ma, J.; Yan, J.; Zhao, A.; Zheng, X.; Wu, G.; et al. Desulfovibrio vulgaris, a potent acetic acid-producing bacterium, attenuates nonalcoholic fatty liver disease in mice. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Chen, Y.Z.; Gu, J.; Chuang, W.T.; Du, Y.F.; Zhang, L.; Lu, M.L.; Xu, J.Y.; Li, H.Q.; Liu, Y.; Feng, H.T.; et al. Slowly Digestible Carbohydrate Diet Ameliorates Hyperglycemia and Hyperlipidemia in High-Fat Diet/Streptozocin-Induced Diabetic Mice. Front. Nutr. 2022, 9, 854725. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, L.; Gao, Y.; Wu, H.; Zhang, J. The dynamic effects of maternal high-calorie diet on glycolipid metabolism and gut microbiota from weaning to adulthood in offspring mice. Front. Nutr. 2022, 9, 941969. [Google Scholar] [CrossRef]
- Pang, Y.; Zheng, Y.; Yang, N.; Zan, M.; Zhang, L.; Ding, W. Potential novel biomarkers in small intestine for obesity/obesity resistance revealed by multi-omics analysis. Lipids Health Dis. 2022, 21, 98. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, H.F.; Wu, C.C.; Chen, C.L.; Ni, Y.H. Pathogenic effects of Desulfovibrio in the gut on fatty liver in diet-induced obese mice and children with obesity. J. Gastroenterol. 2022, 57, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xu, W.; Yu, J.; Liu, Y.; Ma, H.; Ji, C.; Zhang, C.; Xue, J.; Li, R.; Cui, H. Astaxanthin From Haematococcus pluvialis Prevents High-Fat Diet-Induced Hepatic Steatosis and Oxidative Stress in Mice by Gut-Liver Axis Modulating Properties. Front. Nutr. 2022, 9, 840648. [Google Scholar] [CrossRef]
- Zhu, Y.; Wei, Y.L.; Karras, I.; Cai, P.J.; Xiao, Y.H.; Jia, C.L.; Qian, X.L.; Zhu, S.Y.; Zheng, L.J.; Hu, X.; et al. Modulation of the gut microbiota and lipidomic profiles by black chokeberry (Aronia melanocarpa L.) polyphenols via the glycerophospholipid metabolism signaling pathway. Front. Nutr. 2022, 9, 913729. [Google Scholar] [CrossRef]
- Ye, Y.; Shi, L.; Wang, P.; Yang, M.; Zhan, P.; Tian, H.; Liu, J. Water extract of Ferula lehmanni Boiss. prevents high-fat diet-induced overweight and liver injury by modulating the intestinal microbiota in mice. Food Funct. 2022, 13, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Liu, M.; Liu, H.; Jin, Z.; Guan, F.; Ge, S.; Yan, J.; Zheng, M.; Cai, D.; Liu, J. Zeaxanthin ameliorates obesity by activating the beta3-adrenergic receptor to stimulate inguinal fat thermogenesis and modulating the gut microbiota. Food Funct. 2021, 12, 12734–12750. [Google Scholar] [CrossRef]
- Li, S.; You, J.; Wang, Z.; Liu, Y.; Wang, B.; Du, M.; Zou, T. Curcumin alleviates high-fat diet-induced hepatic steatosis and obesity in association with modulation of gut microbiota in mice. Food Res. Int. 2021, 143, 110270. [Google Scholar] [CrossRef]
- Li, A.; Wang, N.; Li, N.; Li, B.; Yan, F.; Song, Y.; Hou, J.; Huo, G. Modulation effect of chenpi extract on gut microbiota in high-fat diet-induced obese C57BL/6 mice. J. Food Biochem. 2021, 45, e13541. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, M.; Sundralingam, U.; Palanisamy, U.D. Polyphenols as Prebiotics in the Management of High-Fat Diet-Induced Obesity: A Systematic Review of Animal Studies. Foods 2021, 10, 299. [Google Scholar] [CrossRef]
- Wang, P.; Gao, J.; Ke, W.; Wang, J.; Li, D.; Liu, R.; Jia, Y.; Wang, X.; Chen, X.; Chen, F.; et al. Resveratrol reduces obesity in high-fat diet-fed mice via modulating the composition and metabolic function of the gut microbiota. Free Radic. Biol. Med. 2020, 156, 83–98. [Google Scholar] [CrossRef]
- Lee, H.B.; Oh, M.J.; Do, M.H.; Kim, Y.S.; Park, H.Y. Molokhia leaf extract prevents gut inflammation and obesity. J. Ethnopharmacol. 2020, 257, 112866. [Google Scholar] [CrossRef]
- Zhang, X.L.; Chen, L.; Yang, J.; Zhao, S.S.; Jin, S.; Ao, N.; Yang, J.; Liu, H.X.; Du, J. Vitamin D alleviates non-alcoholic fatty liver disease via restoring gut microbiota and metabolism. Front. Microbiol. 2023, 14, 1117644. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yan, H.; Lu, Y.; Li, X.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lu, X. Anti-obesity effect of Lactobacillus rhamnosus LS-8 and Lactobacillus crustorum MN047 on high-fat and high-fructose diet mice base on inflammatory response alleviation and gut microbiota regulation. Eur. J. Nutr. 2020, 59, 2709–2728. [Google Scholar] [CrossRef]
- Wang, N.; Sheng, Z.; Zhou, S.; Jiang, F.; Zhang, Z. Chronic lead exposure exacerbates hepatic glucolipid metabolism disorder and gut microbiota dysbiosis in high-fat-diet mice. Food Chem. Toxicol. 2022, 170, 113451. [Google Scholar] [CrossRef] [PubMed]
- Panasevich, M.R.; Meers, G.M.; Linden, M.A.; Booth, F.W.; Perfield, J.W., 2nd; Fritsche, K.L.; Wankhade, U.D.; Chintapalli, S.V.; Shankar, K.; Ibdah, J.A.; et al. High-fat, high-fructose, high-cholesterol feeding causes severe NASH and cecal microbiota dysbiosis in juvenile Ossabaw swine. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E78–E92. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T cell-mediated regulation of the microbiota protects against obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef]
- Buttet, M.; Traynard, V.; Tran, T.T.; Besnard, P.; Poirier, H.; Niot, I. From fatty-acid sensing to chylomicron synthesis: Role of intestinal lipid-binding proteins. Biochimie 2014, 96, 37–47. [Google Scholar] [CrossRef]
- Buttet, M.; Poirier, H.; Traynard, V.; Gaire, K.; Tran, T.T.; Sundaresan, S.; Besnard, P.; Abumrad, N.A.; Niot, I. Deregulated Lipid Sensing by Intestinal CD36 in Diet-Induced Hyperinsulinemic Obese Mouse Model. PLoS ONE 2016, 11, e0145626. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, U.; Murthy, R.T.; Felicita, A.S.; Alshehri, A.; Awadh, W.; Almalki, A.; Vinothkumar, T.S.; Baeshen, H.A.; Bhandi, S.; Kathir, A.; et al. Sulfate-Reducing Bacteria in Patients Undergoing Fixed Orthodontic Treatment. Int. Dent. J. 2023, 73, 274–279. [Google Scholar] [CrossRef]
- Liu, S.; Xie, G.; Chen, M.; He, Y.; Yu, W.; Chen, X.; Mao, W.; Liu, N.; Zhang, Y.; Chang, Q.; et al. Oral microbial dysbiosis in patients with periodontitis and chronic obstructive pulmonary disease. Front. Cell Infect. Microbiol. 2023, 13, 1121399. [Google Scholar] [CrossRef]
- Li, F.; Wang, M.; Wang, J.; Li, R.; Zhang, Y. Alterations to the Gut Microbiota and Their Correlation with Inflammatory Factors in Chronic Kidney Disease. Front. Cell Infect. Microbiol. 2019, 9, 206. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ning, X.; Liu, B.; Dong, R.; Bai, M.; Sun, S. Specific alterations in gut microbiota in patients with chronic kidney disease: An updated systematic review. Ren. Fail. 2021, 43, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Zhang, Y.; Ren, Y.; Shi, J.S.; Xu, Z.H.; Geng, Y. Diversity and Comparison of Intestinal Desulfovibrio in Patients with Liver Cirrhosis and Healthy People. Microorganisms 2023, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Gong, Z.; Sun, G.; Xu, J.; Qi, C.; Sun, W.; Jiang, H.; Cao, P.; Ju, H. Dysbiosis of Gut Microbiota in Patients with Acute Myocardial Infarction. Front. Microbiol. 2021, 12, 680101. [Google Scholar] [CrossRef]
- Yin, J.; Liao, S.X.; He, Y.; Wang, S.; Xia, G.H.; Liu, F.T.; Zhu, J.J.; You, C.; Chen, Q.; Zhou, L.; et al. Dysbiosis of Gut Microbiota With Reduced Trimethylamine-N-Oxide Level in Patients with Large-Artery Atherosclerotic Stroke or Transient Ischemic Attack. J. Am. Heart Assoc. 2015, 4, e002699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva-Millan, M.J.; Leite, G.; Wang, J.; Morales, W.; Parodi, G.; Pimentel, M.L.; Barlow, G.M.; Mathur, R.; Rezaie, A.; Sanchez, M.; et al. Methanogens and Hydrogen Sulfide Producing Bacteria Guide Distinct Gut Microbe Profiles and Irritable Bowel Syndrome Subtypes. Am. J. Gastroenterol. 2022, 117, 2055–2066. [Google Scholar] [CrossRef]
- Gobert, A.P.; Sagrestani, G.; Delmas, E.; Wilson, K.T.; Verriere, T.G.; Dapoigny, M.; Del’homme, C.; Bernalier-Donadille, A. The human intestinal microbiota of constipated-predominant irritable bowel syndrome patients exhibits anti-inflammatory properties. Sci. Rep. 2016, 6, 39399. [Google Scholar] [CrossRef] [Green Version]
- Bellocchi, C.; Fernandez-Ochoa, A.; Montanelli, G.; Vigone, B.; Santaniello, A.; Milani, C.; Quirantes-Pine, R.; Borras-Linares, I.; Ventura, M.; Segura-Carrettero, A.; et al. Microbial and metabolic multi-omic correlations in systemic sclerosis patients. Ann. N. Y. Acad. Sci. 2018, 1421, 97–109. [Google Scholar] [CrossRef]
- Tremlett, H.; Fadrosh, D.W.; Faruqi, A.A.; Zhu, F.; Hart, J.; Roalstad, S.; Graves, J.; Lynch, S.; Waubant, E.; US Network of Pediatric MS Centers. Gut microbiota in early pediatric multiple sclerosis: A case-control study. Eur. J. Neurol. 2016, 23, 1308–1321. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, C.; Coutinho-Silva, R.; Zinkevich, V.; Pearce, C.B.; Ojcius, D.M.; Beech, I. Sulphate-reducing bacteria from ulcerative colitis patients induce apoptosis of gastrointestinal epithelial cells. Microb. Pathog. 2017, 112, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Boopathy, R.; Robichaux, M.; LaFont, D.; Howell, M. Activity of sulfate-reducing bacteria in human periodontal pocket. Can. J. Microbiol. 2002, 48, 1099–1103. [Google Scholar] [CrossRef]
- Loubinoux, J.; Bisson-Boutelliez, C.; Miller, N.; Le Faou, A.E. Isolation of the provisionally named Desulfovibrio fairfieldensis from human periodontal pockets. Oral. Microbiol. Immunol. 2002, 17, 321–323. [Google Scholar] [CrossRef]
- Langendijk, P.S.; Hanssen, J.T.; Van der Hoeven, J.S. Sulfate-reducing bacteria in association with human periodontitis. J. Clin. Periodontol. 2000, 27, 943–950. [Google Scholar] [CrossRef] [PubMed]
- van der Hoeven, J.S.; van den Kieboom, C.W.; Schaeken, M.J. Sulfate-reducing bacteria in the periodontal pocket. Oral. Microbiol. Immunol. 1995, 10, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I.; Coufalova, M.; Vitezova, M.; Rittmann, S.K.R. Sulfate-Reducing Bacteria of the Oral Cavity and Their Relation with Periodontitis-Recent Advances. J. Clin. Med. 2020, 9, 2347. [Google Scholar] [CrossRef]
- Cross, K.L.; Chirania, P.; Xiong, W.; Beall, C.J.; Elkins, J.G.; Giannone, R.J.; Griffen, A.L.; Guss, A.M.; Hettich, R.L.; Joshi, S.S.; et al. Insights into the Evolution of Host Association through the Isolation and Characterization of a Novel Human Periodontal Pathobiont, Desulfobulbus oralis. mBio 2018, 9, e02061-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisson-Boutelliez, C.; Massin, F.; Dumas, D.; Miller, N.; Lozniewski, A. Desulfovibrio spp. survive within KB cells and modulate inflammatory responses. Mol. Oral. Microbiol. 2010, 25, 226–235. [Google Scholar] [CrossRef]
- Singh, S.B.; Coffman, C.N.; Carroll-Portillo, A.; Varga, M.G.; Lin, H.C. Notch Signaling Pathway Is Activated by Sulfate Reducing Bacteria. Front. Cell Infect. Microbiol. 2021, 11, 695299. [Google Scholar] [CrossRef]
- Figueiredo, M.C.; Lobo, S.A.; Sousa, S.H.; Pereira, F.P.; Wall, J.D.; Nobre, L.S.; Saraiva, L.M. Hybrid cluster proteins and flavodiiron proteins afford protection to Desulfovibrio vulgaris upon macrophage infection. J. Bacteriol. 2013, 195, 2684–2690. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.B.; Coffman, C.N.; Varga, M.G.; Carroll-Portillo, A.; Braun, C.A.; Lin, H.C. Intestinal Alkaline Phosphatase Prevents Sulfate Reducing Bacteria-Induced Increased Tight Junction Permeability by Inhibiting Snail Pathway. Front. Cell Infect. Microbiol. 2022, 12, 882498. [Google Scholar] [CrossRef]
- Singh, S.B.; Lin, H.C. Autophagy counters LPS-mediated suppression of lysozyme. Innate Immun. 2017, 23, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Ostaff, M.J.; Stange, E.F.; Wehkamp, J. Antimicrobial peptides and gut microbiota in homeostasis and pathology. EMBO Mol. Med. 2013, 5, 1465–1483. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I.; Vítězová, M.; Fedrová, P.; Vochyanová, Z.; Paráková, L.; Hošek, J. Kinetic properties of growth of intestinal sulphate-reducing bacteria isolated from healthy mice and mice with ulcerative colitis. Acta Veterinaria Brno 2018, 86, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Rowan, F.E.; Docherty, N.G.; Coffey, J.C.; O’Connell, P.R. Sulphate-reducing bacteria and hydrogen sulphide in the aetiology of ulcerative colitis. Br. J. Surg. 2009, 96, 151–158. [Google Scholar] [CrossRef]
- Zehnder, A.J. Biology of Anaerobic Microorganisms; John Wiley and Sons Inc.: Hoboken, NJ, USA, 1988. [Google Scholar]
- Caffrey, S.M.; Voordouw, G. Effect of sulfide on growth physiology and gene expression of Desulfovibrio vulgaris Hildenborough. Antonie Van. Leeuwenhoek 2010, 97, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Andriamihaja, M.; Lan, A.; Khodorova, N.; Audebert, M.; Blouin, J.M.; Grauso, M.; Lancha, L.; Benetti, P.H.; Benamouzig, R.; et al. Detrimental effects for colonocytes of an increased exposure to luminal hydrogen sulfide: The adaptive response. Free Radic. Biol. Med. 2016, 93, 155–164. [Google Scholar] [CrossRef]
- Lodowska, J.; Wolny, D.; Jaworska-Kik, M.; Kurkiewicz, S.; Dzierzewicz, Z.; Weglarz, L. The chemical composition of endotoxin isolated from intestinal strain of Desulfovibrio desulfuricans. Sci. World J. 2012, 2012, 647352. [Google Scholar] [CrossRef] [Green Version]
- Zhang-Sun, W.; Augusto, L.A.; Zhao, L.; Caroff, M. Desulfovibrio desulfuricans isolates from the gut of a single individual: Structural and biological lipid A characterization. FEBS Lett. 2015, 589, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Węglarz, L.; Dzierżewicz, Z.; Orchel, A.; Szczerba, J.; Jaworska-Kik, M.; Wilczok, T. Biological Activity of Desulfovibrio desulfuricans Lipopolysaccharides Evaluated via Interleukin-8 Secretion by Caco-2 Cells. Scand. J. Gastroenterol. 2003, 38, 73–79. [Google Scholar]
- Weglarz, L.; Wawszczyk, J.; Orchel, A.; Jaworska-Kik, M.; Dzierzewicz, Z. Phytic acid modulates in vitro IL-8 and IL-6 release from colonic epithelial cells stimulated with LPS and IL-1beta. Dig. Dis. Sci. 2007, 52, 93–102. [Google Scholar] [CrossRef]
- Sartorio, M.G.; Pardue, E.J.; Feldman, M.F.; Haurat, M.F. Bacterial Outer Membrane Vesicles: From Discovery to Applications. Annu. Rev. Microbiol. 2021, 75, 609–630. [Google Scholar] [CrossRef]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 2005, 437, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loubinoux, J.; Mory, F.; Pereira, I.A.; Le Faou, A.E. Bacteremia caused by a strain of Desulfovibrio related to the provisionally named Desulfovibrio fairfieldensis. J. Clin. Microbiol. 2000, 38, 931–934. [Google Scholar] [CrossRef] [Green Version]
- Fleetwood, A.J.; Lee, M.K.S.; Singleton, W.; Achuthan, A.; Lee, M.C.; O’Brien-Simpson, N.M.; Cook, A.D.; Murphy, A.J.; Dashper, S.G.; Reynolds, E.C.; et al. Metabolic Remodeling, Inflammasome Activation, and Pyroptosis in Macrophages Stimulated by Porphyromonas gingivalis and Its Outer Membrane Vesicles. Front. Cell Infect. Microbiol. 2017, 7, 351. [Google Scholar] [CrossRef] [Green Version]
- Tavano, R.; Franzoso, S.; Cecchini, P.; Cartocci, E.; Oriente, F.; Arico, B.; Papini, E. The membrane expression of Neisseria meningitidis adhesin A (NadA) increases the proimmune effects of MenB OMVs on human macrophages, compared with NadA- OMVs, without further stimulating their proinflammatory activity on circulating monocytes. J. Leukoc. Biol. 2009, 86, 143–153. [Google Scholar] [CrossRef]
- Elmi, A.; Nasher, F.; Jagatia, H.; Gundogdu, O.; Bajaj-Elliott, M.; Wren, B.; Dorrell, N. Campylobacter jejuni outer membrane vesicle-associated proteolytic activity promotes bacterial invasion by mediating cleavage of intestinal epithelial cell E-cadherin and occludin. Cell Microbiol. 2016, 18, 561–572. [Google Scholar] [CrossRef]
- Walian, P.J.; Allen, S.; Shatsky, M.; Zeng, L.; Szakal, E.D.; Liu, H.; Hall, S.C.; Fisher, S.J.; Lam, B.R.; Singer, M.E.; et al. High-throughput isolation and characterization of untagged membrane protein complexes: Outer membrane complexes of Desulfovibrio vulgaris. J. Proteome Res. 2012, 11, 5720–5735. [Google Scholar] [CrossRef]
- Earley, H.; Lennon, G.; Balfe, A.; Kilcoyne, M.; Clyne, M.; Joshi, L.; Carrington, S.; Martin, S.T.; Coffey, J.C.; Winter, D.C.; et al. A Preliminary Study Examining the Binding Capacity of Akkermansia muciniphila and Desulfovibrio spp., to Colonic Mucin in Health and Ulcerative Colitis. PLoS ONE 2015, 10, e0135280. [Google Scholar] [CrossRef] [Green Version]
- Lennon, G.; Balfe, A.; Earley, H.; Devane, L.A.; Lavelle, A.; Winter, D.C.; Coffey, J.C.; O’Connell, P.R. Influences of the colonic microbiome on the mucous gel layer in ulcerative colitis. Gut Microbes 2014, 5, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Longman, R.J.; Poulsom, R.; Corfield, A.P.; Warren, B.F.; Wright, N.A.; Thomas, M.G. Alterations in the composition of the supramucosal defense barrier in relation to disease severity of ulcerative colitis. J. Histochem. Cytochem. 2006, 54, 1335–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultsz, C.; Van Den Berg, F.M.; Ten Kate, F.W.; Tytgat, G.N.; Dankert, J. The intestinal mucus layer from patients with inflammatory bowel disease harbors high numbers of bacteria compared with controls. Gastroenterology 1999, 117, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Raouf, A.H.; Tsai, H.H.; Parker, N.; Hoffman, J.; Walker, R.J.; Rhodes, J.M. Sulphation of colonic and rectal mucin in inflammatory bowel disease: Reduced sulphation of rectal mucus in ulcerative colitis. Clin. Sci. 1992, 83, 623–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.B.; Carroll-Portillo, A.; Lin, H.C. Desulfovibrio in the Gut: The Enemy within? Microorganisms 2023, 11, 1772. https://doi.org/10.3390/microorganisms11071772
Singh SB, Carroll-Portillo A, Lin HC. Desulfovibrio in the Gut: The Enemy within? Microorganisms. 2023; 11(7):1772. https://doi.org/10.3390/microorganisms11071772
Chicago/Turabian StyleSingh, Sudha B., Amanda Carroll-Portillo, and Henry C. Lin. 2023. "Desulfovibrio in the Gut: The Enemy within?" Microorganisms 11, no. 7: 1772. https://doi.org/10.3390/microorganisms11071772