
Genomic and Pathogenicity Mechanisms of the Main Theobroma cacao L. Eukaryotic Pathogens: A Systematic Review

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Planning
2.2. Execution
- Records were clustered based on their relation to the topic structure using affinity propagation [31] with the R package APCluster, version 1.14.10 [32]. The number of clusters was estimated with the R package NbClust, version 3.0.1 [33], which provides the most frequent best solution among a set of estimates proposed by different criteria.
- Word clouds were generated using the R package wordcloud, version 2.6 [34] based on the paper abstracts of each cluster. This allowed for identification and exclusion of clusters not related to the subject of this review. Before excluding a cluster, some papers were randomly selected, and their titles and abstracts were read first.
- Papers in the selected clusters were subjected to conventional screening using the StArt software.
- After the semiautomated screening, the titles and abstracts were read, and the remaining papers obtained from the databases were read fully to select those for the next step of analysis.
2.3. Summarization
2.4. Bioinformatic Analysis
3. Results
3.1. Bibliometric Analysis
3.2. Characterization of Studies
3.3. Genes and Proteins Associated with Pathogenicity
4. Discussion
4.1. Identification of Studies and Pathogens
4.2. Theobroma cacao Pathogens’ Molecular Characterization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Beg, M.S.; Ahmad, S.; Jan, K.; Bashir, K. Status, Supply Chain and Processing of Cocoa—A Review. Trends Food Sci. Technol. 2017, 66, 108–116. [Google Scholar] [CrossRef]
- Ploetz, R.C. Cacao Diseases: Important Threats to Chocolate Production Worldwide. Phytopathology 2007, 97, 1634–1639. [Google Scholar] [CrossRef] [Green Version]
- Fulton, R.H. The Cacao Disease Trilogy: Black Pod, Monilia Pod Rot, and Witches’-Broom. Plant Dis. 1989, 73, 601. [Google Scholar] [CrossRef] [Green Version]
- Evans, H.C. Cacao Diseases—The Trilogy Revisited. Phytopathology 2007, 97, 1640–1643. [Google Scholar] [CrossRef] [Green Version]
- International Cocoa Organization (ICCO). Pests & Diseases. Available online: https://www.icco.org/pests-diseases/ (accessed on 1 December 2022).
- Puig, A.S.; Ali, S.; Strem, M.; Sicher, R.; Gutierrez, O.A.; Bailey, B.A. The Differential Influence of Temperature on Phytophthora megakarya and Phytophthora palmivora Pod Lesion Expansion, Mycelia Growth, Gene Expression, and Metabolite Profiles. Physiol. Mol. Plant Pathol. 2018, 102, 95–112. [Google Scholar] [CrossRef]
- Ali, S.S.; Amoako-Attah, I.; Bailey, R.A.; Strem, M.D.; Schmidt, M.; Akrofi, A.Y.; Surujdeo-Maharaj, S.; Kolawole, O.O.; Begoude, B.A.D.; ten Hoopen, G.M.; et al. PCR-Based Identification of Cacao Black Pod Causal Agents and Identification of Biological Factors Possibly Contributing to Phytophthora megakarya’s Field Dominance in West Africa. Plant Pathol. 2016, 65, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Konam, J.K.; Guest, D.I. Leaf Litter Mulch Reduces the Survival of Phytophthora palmivora under Cocoa Trees in Papua New Guinea. Australas. Plant Pathol. 2002, 31, 381–383. [Google Scholar] [CrossRef]
- Torres-de-la-Cruz, M.; Quevedo-Damián, I.; Ortiz-García, C.F.; del Carmen Lagúnez-Espinoza, L.; Nieto-Angel, D.; Pérez-de la Cruz, M. Control Químico de Moniliophthora Roreri En México. Biotecnia 2019, 21, 55–61. [Google Scholar] [CrossRef]
- Gutiérrez, O.A.; Puig, A.S.; Phillips-Mora, W.; Bailey, B.A.; Ali, S.S.; Mockaitis, K.; Schnell, R.J.; Livingstone, D.; Mustiga, G.; Royaert, S.; et al. SNP Markers Associated with Resistance to Frosty Pod and Black Pod Rot Diseases in an F1 Population of Theobroma cacao L. Tree Genet. Genomes 2021, 17, 28. [Google Scholar] [CrossRef]
- Montezano Fernandes, F.; Vieira de Queiroz, M.; Lopes da Silva, L.; Maria Queiroz Azevedo, D.; Luis Badel, J.; Couto Alfenas, A. Chromosomal Polymorphism of the Ceratocystis fimbriata Species Complex in Brazil. Fungal Genet. Biol. 2022, 162, 103728. [Google Scholar] [CrossRef]
- Brown, J.S.; Phillips-Mora, W.; Power, E.J.; Krol, C.; Cervantes-Martinez, C.; Motamayor, J.C.; Schnell, R.J. Mapping QTLs for Resistance to Frosty Pod and Black Pod Diseases and Horticultural Traits in Theobroma cacao L. Crop Sci. 2007, 47, 1851–1858. [Google Scholar] [CrossRef] [Green Version]
- Mondego, J.M.; Carazzolle, M.F.; Costa, G.G.; Formighieri, E.F.; Parizzi, L.P.; Rincones, J.; Cotomacci, C.; Carraro, D.M.; Cunha, A.F.; Carrer, H.; et al. A Genome Survey of Moniliophthora perniciosa Gives New Insights into Witches’ Broom Disease of Cacao. BMC Genom. 2008, 9, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, C.S.; da Fonseca, R.R.; Batista, T.M.; Barreto, M.A.; Argolo, C.S.; de Carvalho, M.R.; do Amaral, D.O.J.; de Andrade Silva, E.M.; Arévalo-Gardini, E.; Hidalgo, K.S.; et al. Genome Sequence and Effectorome of Moniliophthora perniciosa and Moniliophthora roreri Subpopulations. BMC Genom. 2018, 19, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinhardt, L.W.; Costa, G.G.; Thomazella, D.P.; Teixeira, P.J.P.; Carazzolle, M.; Schuster, S.C.; Carlson, J.E.; Guiltinan, M.J.; Mieczkowski, P.; Farmer, A.; et al. Genome and Secretome Analysis of the Hemibiotrophic Fungal Pathogen, Moniliophthora roreri, Which Causes Frosty Pod Rot Disease of Cacao: Mechanisms of the Biotrophic and Necrotrophic Phases. BMC Genom. 2014, 15, 164. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.S.; Shao, J.; Lary, D.J.; Strem, M.D.; Meinhardt, L.W.; Bailey, B.A. Phytophthora megakarya and P. palmivora, Causal Agents of Black Pod Rot, Induce Similar Plant Defense Responses Late during Infection of Susceptible Cacao Pods. Front. Plant Sci. 2017, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Morales-Cruz, A.; Ali, S.S.; Minio, A.; Figueroa-Balderas, R.; García, J.F.; Kasuga, T.; Puig, A.S.; Marelli, J.-P.; Bailey, B.A.; Cantu, D. Independent Whole-Genome Duplications Define the Architecture of the Genomes of the Devastating West African Cacao Black Pod Pathogen Phytophthora megakarya and Its Close Relative Phytophthora palmivora. G3 Genes Genomes Genet. 2020, 10, 2241–2255. [Google Scholar] [CrossRef]
- Molano, E.P.L.; Cabrera, O.G.; Jose, J.; do Nascimento, L.C.; Carazzolle, M.F.; Teixeira, P.J.P.L.; Alvarez, J.C.; Tiburcio, R.A.; Tokimatu Filho, P.M.; de Lima, G.M.A.; et al. Ceratocystis cacaofunesta Genome Analysis Reveals a Large Expansion of Extracellular Phosphatidylinositol-Specific Phospholipase-C Genes (PI-PLC). BMC Genom. 2018, 19, 58. [Google Scholar] [CrossRef] [Green Version]
- Garcia, O.; Macedo, J.A.N.; Tibúrcio, R.; Zaparoli, G.; Rincones, J.; Bittencourt, L.M.C.; Ceita, G.O.; Micheli, F.; Gesteira, A.; Mariano, A.C.; et al. Characterization of Necrosis and Ethylene-Inducing Proteins (NEP) in the Basidiomycete Moniliophthora perniciosa, the Causal Agent of Witches’ Broom in Theobroma cacao. Mycol. Res. 2007, 111, 443–455. [Google Scholar] [CrossRef]
- Bae, H.; Bowers, J.H.; Tooley, P.W.; Bailey, B.A. NEP1 Orthologs Encoding Necrosis and Ethylene Inducing Proteins Exist as a Multigene Family in Phytophthora megakarya, Causal Agent of Black Pod Disease on Cacao. Mycol. Res. 2005, 109, 1373–1385. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Barsottini, M.R.; de Oliveira, J.F.; Adamoski, D.; Teixeira, P.J.P.L.; do Prado, P.F.V.; Tiezzi, H.O.; Sforça, M.L.; Cassago, A.; Portugal, R.V.; de Oliveira, P.S.L.; et al. Functional Diversification of Cerato-Platanins in Moniliophthora perniciosa as Seen by Differential Expression and Protein Function Specialization. Mol. Plant Microbe Interact. 2013, 26, 1281–1293. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.S.; Mora-Ocampo, I.Y.; de Novais, D.P.S.; Aguiar, E.R.G.R.; Pirovani, C.P. State of the Art of the Molecular Biology of the Interaction between Cocoa and Witches’ Broom Disease: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 5684. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, D.L.; Alvarez, J.C.; Mosquera, S. Frosty Pod Rot: A Major Threat to Cacao Plantations on the Move. Trop. Plant Pathol. 2022, 47, 187–200. [Google Scholar] [CrossRef]
- Marelli, J.-P.; Guest, D.I.; Bailey, B.A.; Evans, H.C.; Brown, J.K.; Junaid, M.; Barreto, R.W.; Lisboa, D.O.; Puig, A.S. Chocolate Under Threat from Old and New Cacao Diseases. Phytopathology 2019, 109, 1331–1343. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, S.; Silva, C.; Hernandes, E.; Octaviano, F.; Di Thommazo, A.; Belgamo, A. Improvements in the StArt Tool to Better Support the Systematic Review Process. In Proceedings of the 20th International Conference on Evaluation and Assessment in Software Engineering, Limerick Ireland, 1–3 June 2016; pp. 1–5. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023.
- Blei, D.M. Latent Dirichlet Allocation. J. Mach. Learn. Res. 2003, 3, 993–1022. [Google Scholar]
- Jones, T. TextmineR: Functions for Text Mining and Topic Modeling. Available online: https://CRAN.R-project.org/package=textmineR (accessed on 9 December 2022).
- Nikita, M. Ldatuning: Tuning of the Latent Dirichlet Allocation Models Parameters. Available online: https://CRAN.R-project.org/package=ldatuning (accessed on 9 December 2022).
- Frey, B.J.; Dueck, D. Clustering by Passing Messages Between Data Points. Science 2007, 315, 972–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodenhofer, U.; Kothmeier, A.; Hochreiter, S. APCluster: An R Package for Affinity Propagation Clustering. Bioinformatics 2011, 27, 2463–2464. [Google Scholar] [CrossRef] [Green Version]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R Package for Determining the Relevant Number of Clusters in a Data Set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Fellows, I. Wordcloud Makes Words Less Cloudy. Available online: https://blog.fellstat.com/?p=248 (accessed on 7 June 2023).
- Müller, K.; Ooms, J.; James, D.; DebRoy, S.; Wickham, H.; Horner, J. RMariaDB: Database Interface and MariaDB Driver. Available online: https://CRAN.R-project.org/package=RMariaDB (accessed on 9 December 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-Tool for Comprehensive Science Mapping Analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Van Eck, N.J.; Waltman, L. Visualizing Bibliometric Networks. In Measuring Scholarly Impact; Ding, Y., Rousseau, R., Wolfram, D., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 285–320. ISBN 978-3-319-10376-1. [Google Scholar]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A Web Server for Whole-Genome Comparison and Annotation of Orthologous Clusters across Multiple Species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Elkan, C. Fitting a Mixture Model by Expectation Maximization to Discover Motifs in Bipolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Ali, S.S.; Shao, J.; Strem, M.D.; Phillips-Mora, W.; Zhang, D.; Meinhardt, L.W.; Bailey, B.A. Combination of RNAseq and SNP Nanofluidic Array Reveals the Center of Genetic Diversity of Cacao Pathogen Moniliophthora roreri in the Upper Magdalena Valley of Colombia and Its Clonality. Front. Microbiol. 2015, 6, 850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaimes, Y.Y.; Gonzalez, C.; Rojas, J.; Cornejo, O.E.; Mideros, M.F.; Restrepo, S.; Cilas, C.; Furtado, E.L. Geographic Differentiation and Population Genetic Structure of Moniliophthora roreri in the Principal Cocoa Production Areas in Colombia. Plant Dis. 2016, 100, 1548–1558. [Google Scholar] [CrossRef] [Green Version]
- Maridueña-Zavala, M.G.; Freire-Peñaherrera, A.; Espinoza-Lozano, R.F.; Villavicencio-Vasquez, M.; Jimenez-Feijoo, M.; Cevallos-Cevallos, J.M. Genetic Characterization of Moniliophthora perniciosa from Ecuador and in Vitro Sensitivity to Compost Tea. Eur. J. Plant Pathol. 2019, 154, 943–959. [Google Scholar] [CrossRef]
- Brugman, E.; Wibowo, A.; Widiastuti, A. Phytophthora palmivora from Sulawesi and Java Islands, Indonesia, Reveals High Genotypic Diversity and Lack of Population Structure. Fungal Biol. 2022, 126, 267–276. [Google Scholar] [CrossRef]
- Veerappa Hanumanthappa, P.; Hegde, V.; Kuriyathadka Mahalingeshwara, S.; Muliyar Krishna, R.; Kaitheri Edathil, R.; Pallem, C. Differentiation of Phytophthora Species Associated with Plantation Crops Using PCR and High-Resolution Melting Curve Analysis. J. Plant Pathol. 2018, 100, 233–240. [Google Scholar] [CrossRef]
- De Oliveira, B.V.; Teixeira, G.S.; Reis, O.; Barau, J.G.; Teixeira, P.J.P.L.; do Rio, M.C.S.; Domingues, R.R.; Meinhardt, L.W.; Paes Leme, A.F.; Rincones, J.; et al. A Potential Role for an Extracellular Methanol Oxidase Secreted by Moniliophthora perniciosa in Witches’ Broom Disease in Cacao. Fungal Genet. Biol. 2012, 49, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Pungartnik, C.; Melo, S.C.O.; Basso, T.S.; Macena, W.G.; Cascardo, J.C.M.; Brendel, M. Reactive Oxygen Species and Autophagy Play a Role in Survival and Differentiation of the Phytopathogen Moniliophthora perniciosa. Fungal Genet. Biol. 2009, 46, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Santana, L.S.; Costa, M.G.C.; Pirovani, N.M.; Almeida, A.F.; Alvim, F.C.; Pirovani, C.P. TcCYS4, a Cystatin from Cocoa, Reduces Necrosis Triggered by MpNEP2 in Tobacco Plants. Genet. Mol. Res. 2014, 13, 7636–7648. [Google Scholar] [CrossRef] [PubMed]
- Artero, A.S.; Silva, J.Q.; Albuquerque, P.S.B.; Bressan, E.A.; Leal, G.A.; Sebbenn, A.M.; Griffith, G.W.; Figueira, A. Spatial Genetic Structure and Dispersal of the Cacao Pathogen Moniliophthora perniciosa in the Brazilian Amazon. Plant Pathol. 2017, 66, 912–923. [Google Scholar] [CrossRef] [Green Version]
- Gramacho, K.P.; Risterucci, A.M.; Lanaud, C.; Lima, L.S.; Lopes, U.V. Characterization of Microsatellites in the Fungal Plant Pathogen Crinipellis Perniciosa: PRIMER NOTE. Mol. Ecol. Notes 2006, 7, 153–155. [Google Scholar] [CrossRef]
- Rincones, J.; Mazotti, G.D.; Griffith, G.W.; Pomela, A.; Figueira, A.; Leal, G.A.; Queiroz, M.V.; Pereira, J.F.; Azevedo, R.A.; Pereira, G.A.G.; et al. Genetic Variability and Chromosome-Length Polymorphisms of the Witches’ Broom Pathogen Crinipellis perniciosa from Various Plant Hosts in South America. Mycol. Res. 2006, 110, 821–832. [Google Scholar] [CrossRef]
- Mares, J.H.; Gramacho, K.P.; Santana, J.O.; Oliveira de Souza, A.; Alvim, F.C.; Pirovani, C.P. Hydrosoluble Phylloplane Components of Theobroma cacao Modulate the Metabolism of Moniliophthora Perniciosa Spores during Germination. Fungal Biol. 2020, 124, 73–81. [Google Scholar] [CrossRef]
- Santos Junior, M.C.; Gonçalves, P.A.; Taranto, A.G.; Koblitz, M.G.B.; Góes-Neto, A.; Pirovani, C.P.; Cascardo, J.C.M.; da Cruz, S.H.; Zingali, R.B.; Pereira, G.A.G.; et al. Purification, Characterization and Structural Determination of UDP-N-Acetylglucosamine Pyrophosphorylase Produced by Moniliophthora perniciosa. J. Braz. Chem. Soc. 2011, 22, 1015–1023. [Google Scholar] [CrossRef]
- Teixeira, P.J.P.L.; Thomazella, D.P.T.; Vidal, R.O.; do Prado, P.F.V.; Reis, O.; Baroni, R.M.; Franco, S.F.; Mieczkowski, P.; Pereira, G.A.G.; Mondego, J.M.C. The Fungal Pathogen Moniliophthora perniciosa Has Genes Similar to Plant PR-1 That Are Highly Expressed during Its Interaction with Cacao. PLoS ONE 2012, 7, e45929. [Google Scholar] [CrossRef]
- Bailey, B.A.; Crozier, J.; Sicher, R.C.; Strem, M.D.; Melnick, R.; Carazzolle, M.F.; Costa, G.G.L.; Pereira, G.A.G.; Zhang, D.; Maximova, S.; et al. Dynamic Changes in Pod and Fungal Physiology Associated with the Shift from Biotrophy to Necrotrophy during the Infection of Theobroma cacao by Moniliophthora roreri. Physiol. Mol. Plant Pathol. 2013, 81, 84–96. [Google Scholar] [CrossRef]
- Melnick, R.L.; Strem, M.D.; Crozier, J.; Sicher, R.C.; Bailey, B.A. Molecular and Metabolic Changes of Cherelle Wilt of Cacao and Its Effect on Moniliophthora roreri. Physiol. Mol. Plant Pathol. 2013, 84, 153–162. [Google Scholar] [CrossRef]
- Aime, M.C.; Phillips-Mora, W. The Causal Agents of Witches’ Broom and Frosty Pod Rot of Cacao (Chocolate, Theobroma cacao) Form a New Lineage of Marasmiaceae. Mycologia 2005, 97, 1012–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelbrecht, C.J.B.; Harrington, T.C. Intersterility, Morphology and Taxonomy of Ceratocystis fimbriata on Sweet Potato, Cacao and Sycamore. Mycologia 2005, 97, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Decloquement, J.; Ramos-Sobrinho, R.; Elias, S.G.; Britto, D.S.; Puig, A.S.; Reis, A.; da Silva, R.A.F.; Honorato-Júnior, J.; Martins Newman Luz, E.D.; Pinho, D.B.; et al. Phytophthora theobromicola sp. Nov.: A New Species Causing Black Pod Disease on Cacao in Brazil. Front. Microbiol. 2021, 12, 537399. [Google Scholar] [CrossRef] [PubMed]
- Santana, M.F.; de Araújo, E.F.; de Souza, J.T.; Mizubuti, E.S.G.; de Queiroz, M.V. Development of Molecular Markers Based on Retrotransposons for the Analysis of Genetic Variability in Moniliophthora perniciosa. Eur. J. Plant Pathol. 2012, 134, 497–507. [Google Scholar] [CrossRef]
- Silva, J.R.Q.; Figueira, A.; Pereira, G.A.G.; Albuquerque, P. Development of Novel Microsatellites from Moniliophthora perniciosa, Causal Agent of the Witches’ Broom Disease of Theobroma cacao: Permanent genetic resources. Mol. Ecol. Resour. 2008, 8, 783–785. [Google Scholar] [CrossRef]
- Melo, B.L.B.; de Souza, J.T.; Santos, R.M.F.; Rehner, S.A.; Solis, K.H.; Suarez, C.; Hebbar, P.K.; Lemos, L.S.L.; Gramacho, K.P. Development of Microsatellites for the Cacao Frosty Pod Rot Pathogen, Moniliophthora roreri. For. Pathol. 2014, 44, 320–324. [Google Scholar] [CrossRef]
- Chowdappa, P.; Brayford, D.; Smith, J.; Flood, J. Molecular Discrimination of Phytophthora Isolates on Cocoa and Their Relationship with Coconut, Black Pepper and Bell Pepper Isolates Based on RDNA Repeat and AFLP Fingerprints. Curr. Sci. 2003, 84, 1235–1238. [Google Scholar]
- Guha Roy, S.; Bhattacharyya, S.; Mukherjee, S.K.; Khatua, D.C. Molecular Identification of Phytophthora spp. Affecting Some Economically Important Crops in Eastern India through ITS-RFLP and Sequencing of the ITS Region: Phytophthora spp. Affecting Crops in Eastern India. J. Phytopathol. 2009, 157, 666–674. [Google Scholar] [CrossRef]
- Mfegue, C.V.; Herail, C.; Adreit, H.; Mbenoun, M.; Techou, Z.; Ten Hoopen, M.; Tharreau, D.; Ducamp, M. Microsatellite Markers for Population Studies of Phytophthora megakarya (Pythiaceae), a Cacao Pathogen in Africa. Am. J. Bot. 2012, 99, e353–e356. [Google Scholar] [CrossRef] [Green Version]
- Rincones, J.; Scarpari, L.M.; Carazzolle, M.F.; Mondego, J.M.C.; Formighieri, E.F.; Barau, J.G.; Costa, G.G.L.; Carraro, D.M.; Brentani, H.P.; Vilas-Boas, L.A.; et al. Differential Gene Expression Between the Biotrophic-Like and Saprotrophic Mycelia of the Witches’ Broom Pathogen Moniliophthora perniciosa. Mol. Plant Microbe Interact 2008, 21, 891–908. [Google Scholar] [CrossRef] [Green Version]
- De Freitas Franco, S.; Baroni, R.M.; Carazzolle, M.F.; Teixeira, P.J.P.L.; Reis, O.; Pereira, G.A.G.; Mondego, J.M.C. Genomic Analyses and Expression Evaluation of Thaumatin-like Gene Family in the Cacao Fungal Pathogen Moniliophthora perniciosa. Biochem. Biophys. Res. Commun. 2015, 466, 629–636. [Google Scholar] [CrossRef]
- Bailey, B.A.; Melnick, R.L.; Strem, M.D.; Crozier, J.; Shao, J.; Sicher, R.; Phillips-Mora, W.; Ali, S.S.; Zhang, D.; Meinhardt, L. Differential Gene Expression by Moniliophthora roreri While Overcoming Cacao Tolerance in the Field: The M. roreri Interaction with Tolerant Cacao. Mol. Plant Pathol. 2014, 15, 711–729. [Google Scholar] [CrossRef]
- Bailey, B.A.; Ali, S.S.; Strem, M.D.; Meinhardt, L.W. Morphological Variants of Moniliophthora roreri on Artificial Media and the Biotroph/Necrotroph Shift. Fungal Biol. 2018, 122, 701–716. [Google Scholar] [CrossRef] [PubMed]
- Masanto, M.; Wibowo, A.; Ridwan, N.F.; Sawitri, W.D.; Kageyama, K.; Subandiyah, S. The Expression of Pathogenicity-Related Genes in Phytophthora palmivora Causing Black Pod Rot Disease on Cacao (Theobroma cacao L.) in Indonesia. J. Plant Interact. 2021, 16, 284–295. [Google Scholar] [CrossRef]
- Ali, S.S.; Asman, A.; Shao, J.; Firmansyah, A.P.; Susilo, A.W.; Rosmana, A.; McMahon, P.; Junaid, M.; Guest, D.; Kheng, T.Y.; et al. Draft Genome Sequence of Fastidious Pathogen Ceratobasidium theobromae, Which Causes Vascular-Streak Dieback in Theobroma Cacao. Fungal Biol. Biotechnol. 2019, 6, 14. [Google Scholar] [CrossRef]
- Santos Gomes, D.; de Andrade Silva, E.M.; de Andrade Rosa, E.C.; Silva Gualberto, N.G.; de Jesus Souza, M.Á.; Santos, G.; Pirovani, C.P.; Micheli, F. Identification of a Key Protein Set Involved in Moniliophthora perniciosa Necrotrophic Mycelium and Basidiocarp Development. Fungal Genet. Biol. 2021, 157, 103635. [Google Scholar] [CrossRef]
- Silva, F.A.C.; Pirovani, C.P.; Menezes, S.; Pungartnik, C.; Santiago, A.S.; Costa, M.G.C.; Micheli, F.; Gesteira, A.S. Proteomic Response of Moniliophthora perniciosa Exposed to Pathogenesis-Related Protein-10 from Theobroma cacao. Genet. Mol. Res. 2013, 12, 4855–4868. [Google Scholar] [CrossRef] [PubMed]
- Zaparoli, G.; de Oliveira Barsottini, M.R.; de Oliveira, J.F.; Dyszy, F.; Teixeira, P.J.P.L.; Barau, J.G.; Garcia, O.; Costa-Filho, A.J.; Ambrosio, A.L.B.; Pereira, G.A.G.; et al. The Crystal Structure of Necrosis- and Ethylene-Inducing Protein 2 from the Causal Agent of Cacao’s Witches’ Broom Disease Reveals Key Elements for Its Activity. Biochemistry 2011, 50, 9901–9910. [Google Scholar] [CrossRef]
- Baroni, R.M.; Luo, Z.; Darwiche, R.; Hudspeth, E.M.; Schneiter, R.; Pereira, G.A.G.; Mondego, J.M.C.; Asojo, O.A. Crystal Structure of MpPR-1i, a SCP/TAPS Protein from Moniliophthora perniciosa, the Fungus That Causes Witches’ Broom Disease of Cacao. Sci. Rep. 2017, 7, 7818. [Google Scholar] [CrossRef]
- Zaparoli, G.; Cabrera, O.G.; Medrano, F.J.; Tiburcio, R.; Lacerda, G.; Pereira, G.G. Identification of a Second Family of Genes in Moniliophthora perniciosa, the Causal Agent of Witches’ Broom Disease in Cacao, Encoding Necrosis-Inducing Proteins Similar to Cerato-Platanins. Mycol. Res. 2009, 113, 61–72. [Google Scholar] [CrossRef]
- Teh, C.-Y.; Pang, C.-L.; Tor, X.-Y.; Ho, P.-Y.; Lim, Y.-Y.; Namasivayam, P.; Ho, C.-L. Molecular Cloning and Functional Analysis of a Necrosis and Ethylene Inducing Protein (NEP) from Ganoderma Boninense. Physiol. Mol. Plant Pathol. 2019, 106, 42–48. [Google Scholar] [CrossRef]
- Golinelli, D.; Nuzzolese, A.G.; Sanmarchi, F.; Bulla, L.; Mongiovì, M.; Gangemi, A.; Rucci, P. Semi-Automatic Systematic Literature Reviews and Information Extraction of COVID-19 Scientific Evidence: Description and Preliminary Results of the COKE Project. Information 2022, 13, 117. [Google Scholar] [CrossRef]
- Yu, Z.; Carver, J.C.; Rothermel, G.; Menzies, T. Assessing Expert System-Assisted Literature Reviews with a Case Study. Expert Syst. Appl. 2022, 200, 116958. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 1 December 2022).
- Pereira, J.L.; de Almeida, L.C.C.; Santos, S.M. Witches’ Broom Disease of Cocoa in Bahia: Attempts at Eradication and Containment. Crop Prot. 1996, 15, 743–752. [Google Scholar] [CrossRef]
- Ekdale, B.; Rinaldi, A.; Ashfaquzzaman, M.; Khanjani, M.; Matanji, F.; Stoldt, R.; Tully, M. Geographic Disparities in Knowledge Production: A Big Data Analysis of Peer-Reviewed Communication Publications from 1990 to 2019. Int. J. Commun. 2022, 16, 2498–2525. [Google Scholar]
- SCIMAGO. Scimago Journal & Country Rank. Available online: https://www.scimagojr.com/countryrank.php (accessed on 7 June 2023).
- Paladines-Rezabala, A.; Moreira-Morrillo, A.A.; Mieles, A.E.; Garcés-Fiallos, F.R. Advances in Understanding of the Interaction between Ceratocystis cacaofunesta and Xyleborus Ferrugineus (Coleoptera: Curculionidae: Scolytinae) on Cocoa Trees. Sci. Agropecu. 2022, 13, 43–52. [Google Scholar] [CrossRef]
- Ramos-Sobrinho, R.; Chingandu, N.; Gutierrez, O.A.; Marelli, J.P.; Brown, J.K. A Complex of Badnavirus Species Infecting Cacao Reveals Mixed Infections, Extensive Genomic Variability, and Interspecific Recombination. Viruses 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Ocampo, I.Y.; Pirovani, C.P.; Luz, E.D.M.N.; Rêgo, A.P.B.; Silva, E.M.A.; Rhodes-Valbuena, M.; Corrêa, R.X. Ceratocystis cacaofunesta Differentially Modulates the Proteome in Xylem-Enriched Tissue of Cocoa Genotypes with Contrasting Resistance to Ceratocystis Wilt. Planta 2021, 254, 94. [Google Scholar] [CrossRef] [PubMed]
- Mucherino Muñoz, J.J.; De Melo, C.A.F.; Santana Silva, R.J.; Luz, E.D.M.N.; Corrêa, R.X. Structural and Functional Genomics of the Resistance of Cacao to Phytophthora palmivora. Pathogens 2021, 10, 961. [Google Scholar] [CrossRef]
- Barreto, M.A.; Rosa, J.R.B.F.; Holanda, I.S.A.; Cardoso-Silva, C.B.; Vildoso, C.I.A.; Ahnert, D.; Souza, M.M.; Corrêa, R.X.; Royaert, S.; Marelli, J.; et al. QTL Mapping and Identification of Corresponding Genomic Regions for Black Pod Disease Resistance to Three Phytophthora Species in Theobroma cacao L. Euphytica 2018, 214, 188. [Google Scholar] [CrossRef]
- Albuquerque, P.S.B. Mapas de Ligação e Identificação de Locos Controladores de Características Quantitativas (QTLs) Associados à Resistência a Crinipellis perniciosa Em Acessos de Cacaueiro (Theobroma cacao) Originários da Amazônia Brasileira; Universidade de São Paulo: Piracicaba, Brazil, 2006. [Google Scholar]
- Jaimez, R.E.; Barragan, L.; Fernández-Niño, M.; Wessjohann, L.A.; Cedeño-Garcia, G.; Sotomayor Cantos, I.; Arteaga, F. Theobroma cacao L. Cultivar CCN 51: A Comprehensive Review on Origin, Genetics, Sensory Properties, Production Dynamics, and Physiological Aspects. PeerJ 2022, 10, e12676. [Google Scholar] [CrossRef] [PubMed]
- De Mello Paim, V.R.L.; Luz, E.D.M.N.; Pires, J.L.; Silva, S.D.V.M.; de Souza, J.T.; Albuquerque, P.S.B.; Santos Filho, L.P. dos Sources of Resistance to Crinipellis perniciosa in Progenies of Cacao Accessions Collected in the Brazilian Amazon. Sci. Agric. 2006, 63, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Tirado-Gallego, P.A.; Lopera-Álvarez, A.; Ríos-Osorio, L.A. Estrategias de Control de Moniliophthora roreri y Moniliophthora perniciosa en Theobroma cacao L.: Revisión Sistemática. Cienc. Tecnol. Agropecu. 2016, 17, 417–430. [Google Scholar] [CrossRef] [Green Version]
- Krauss, U.; Hidalgo, E.; Bateman, R.; Adonijah, V.; Arroyo, C.; García, J.; Crozier, J.; Brown, N.A.; ten Hoopen, G.M.; Holmes, K.A. Improving the Formulation and Timing of Application of Endophytic Biocontrol and Chemical Agents against Frosty Pod Rot (Moniliophthora roreri) in Cocoa (Theobroma cacao). Biol. Control 2010, 54, 230–240. [Google Scholar] [CrossRef]
- Loguercio, L.L.; de Carvalho, A.C.; Niella, G.R.; De Souza, J.T.; Pomella, A.W.V. Selection of Trichoderma stromaticum Isolates for Efficient Biological Control of Witches’ Broom Disease in Cacao. Biol. Control 2009, 51, 130–139. [Google Scholar] [CrossRef]
- Bertier, L.; Leus, L.; D’hondt, L.; de Cock, A.W.A.M.; Höfte, M. Host Adaptation and Speciation through Hybridization and Polyploidy in Phytophthora. PLoS ONE 2013, 8, e85385. [Google Scholar] [CrossRef] [Green Version]
- Tiburcio, R.A.; Costa, G.G.L.; Carazzolle, M.F.; Mondego, J.M.C.; Schuster, S.C.; Carlson, J.E.; Guiltinan, M.J.; Bailey, B.A.; Mieczkowski, P.; Meinhardt, L.W.; et al. Genes Acquired by Horizontal Transfer Are Potentially Involved in the Evolution of Phytopathogenicity in Moniliophthora perniciosa and Moniliophthora roreri, Two of the Major Pathogens of Cacao. J. Mol. Evol. 2010, 70, 85–97. [Google Scholar] [CrossRef]
- Vries, S.; Stukenbrock, E.H.; Rose, L.E. Rapid Evolution in Plant–Microbe Interactions—An Evolutionary Genomics Perspective. New Phytol. 2020, 226, 1256–1262. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xiao, S.; Zheng, L.; Pan, Y.; Zhao, D.; Zhang, D.; Li, Q.; Zhu, J.; Yang, Z. Multiomic Approaches Reveal Novel Lineage-Specific Effectors in the Potato and Tomato Early Blight Pathogen Alternaria Solani. Phytopathol. Res. 2022, 4, 29. [Google Scholar] [CrossRef]
- Kronmiller, B.A.; Feau, N.; Shen, D.; Tabima, J.F.; Ali, S.S.; Armitage, A.D.; Arredondo, F.; Bailey, B.A.; Bollmann, S.R.; Dale, A.; et al. Comparative Genomic Analysis of 31 Phytophthora Genomes Reveals Genome Plasticity and Horizontal Gene Transfer. Mol. Plant-Microbe Interact. 2023, 36, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Soanes, D.; Richards, T.A. Horizontal Gene Transfer in Eukaryotic Plant Pathogens. Annu. Rev. Phytopathol. 2014, 52, 583–614. [Google Scholar] [CrossRef] [PubMed]
- Mehrabi, R.; Bahkali, A.H.; Abd-Elsalam, K.A.; Moslem, M.; Ben M’Barek, S.; Gohari, A.M.; Jashni, M.K.; Stergiopoulos, I.; Kema, G.H.J.; de Wit, P.J.G.M. Horizontal Gene and Chromosome Transfer in Plant Pathogenic Fungi Affecting Host Range. FEMS Microbiol. Rev. 2011, 35, 542–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayers, C.G.; Harrington, T.C.; Wai, A.; Hausner, G. Recent and Ongoing Horizontal Transfer of Mitochondrial Introns Between Two Fungal Tree Pathogens. Front. Microbiol. 2021, 12, 656609. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Questions |
|---|
| What genera and species of eukaryotic microorganism pathogens of T. cacao have been the focus of omics studies in recent years? |
| Which molecular techniques have been applied, and what kind of data are available for each of these species? |
| How is scientific production about this subject distributed globally? |
| Which scientific journals and subject areas contain the majority of available studies? |
| How are the most cited papers found in the systematic review related to each other? |
| Which proteins for each species are already associated with pathogenicity in the current literature? |
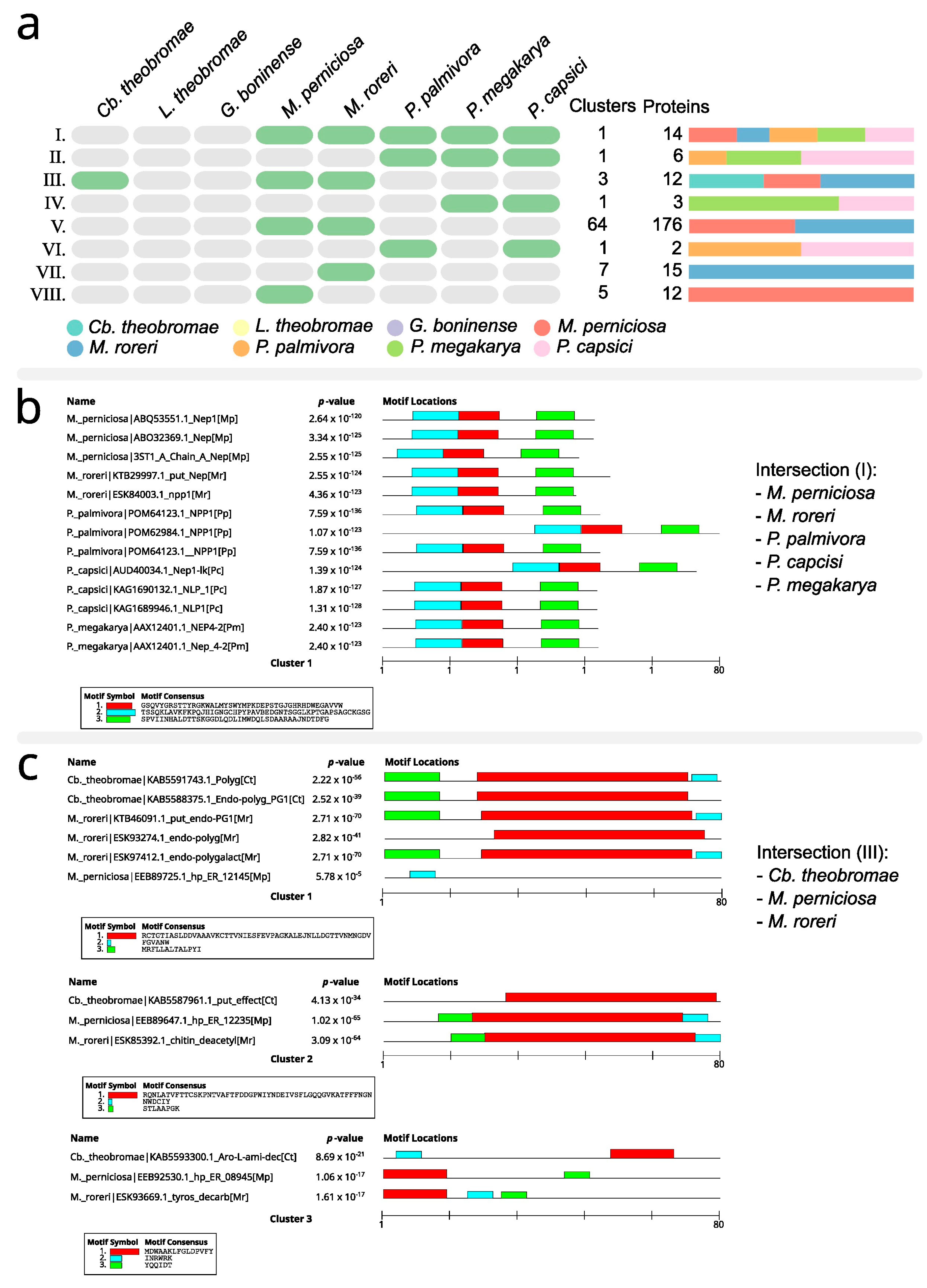
| Do these proteins have orthologs among the T. cacao pathogens discussed in the systematic review? |
| Database | String |
|---|---|
| Scopus | TITLE-ABS-KEY (cocoa OR cacao AND pathogen* OR disease*) AND PUBYEAR > 1999 AND (LIMIT-TO (DOCTYPE, “ar”)) AND (LIMIT-TO (LANGUAGE, “English”)) |
| Web of science | TS = (cocoa OR cacao) AND TS = (pathogen* OR disease*) AND Language: (English) AND DOCUMENT TYPE: (Article) |
| Paper | Species | Assembly Deposit | Genome Size (Mb) | Number of Gene Models |
|---|---|---|---|---|
| [13] | M. perniciosa | Genbank: GCA_000183025.1 | 26.7 | 16,329 |
| [15] | M. roreri | Genbank: GCA_000488995.1 | 52.2 | 17,910 |
| [16] | P. megakarya | Genbank: GCA_002215365.1 | 101.18 | 42,036 |
| P. palmivora | Genbank: GCA_002911725.1 | 107.42 | 44,327 | |
| [18] | C. cacaofunesta | Genbank: GCA_002776505.1 | 30.5 | 7382 |
| [14] | M. perniciosa (MP 4145) + | http://nbcgib.uesc.br/mperniciosa (accessed on 27 April 2023) | 47.01 | 14,210 |
| M. roreri | 45.17 | 14,154 | ||
| [74] | C. theobromae | Genbank: GCA_009078325.1 | 31.2 | 9264 |
| [17] | P. megakarya | http://www.cacaopathogenomics.com/ (accessed on 27 April 2023) | 222.04 ± 25.19 * | 57,577 ± 7904 * |
| P. palmivora | 135.32 ± 17.21 * | 36,778 ± 4481 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Novais, D.P.S.; Batista, T.M.; Costa, E.A.; Pirovani, C.P. Genomic and Pathogenicity Mechanisms of the Main Theobroma cacao L. Eukaryotic Pathogens: A Systematic Review. Microorganisms 2023, 11, 1567. https://doi.org/10.3390/microorganisms11061567
de Novais DPS, Batista TM, Costa EA, Pirovani CP. Genomic and Pathogenicity Mechanisms of the Main Theobroma cacao L. Eukaryotic Pathogens: A Systematic Review. Microorganisms. 2023; 11(6):1567. https://doi.org/10.3390/microorganisms11061567
Chicago/Turabian Stylede Novais, Diogo Pereira Silva, Thiago Mafra Batista, Eduardo Almeida Costa, and Carlos Priminho Pirovani. 2023. "Genomic and Pathogenicity Mechanisms of the Main Theobroma cacao L. Eukaryotic Pathogens: A Systematic Review" Microorganisms 11, no. 6: 1567. https://doi.org/10.3390/microorganisms11061567