
Exploring Synergistic Combinations in Extended and Pan-Drug Resistant (XDR and PDR) Whole Genome Sequenced Acinetobacter baumannii
 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Whole Genome Sequencing (WGS)
2.2. MIC Determination
2.3. Antimicrobial Synergy Testing
2.4. Checkerboard Assay
2.5. Time-Kill Assay
2.6. Statistical Analysis
3. Results
3.1. Minimal Inhibitory Concentrations (MIC) of Fosfomycin, Meropenem, Tigecycline, Colistin and Amikacin in Acinetobacter baumannii
3.2. Synergy Outcomes
3.3. Outcome of Fosfomycin-Meropenem Combination by Checkerboard Assay
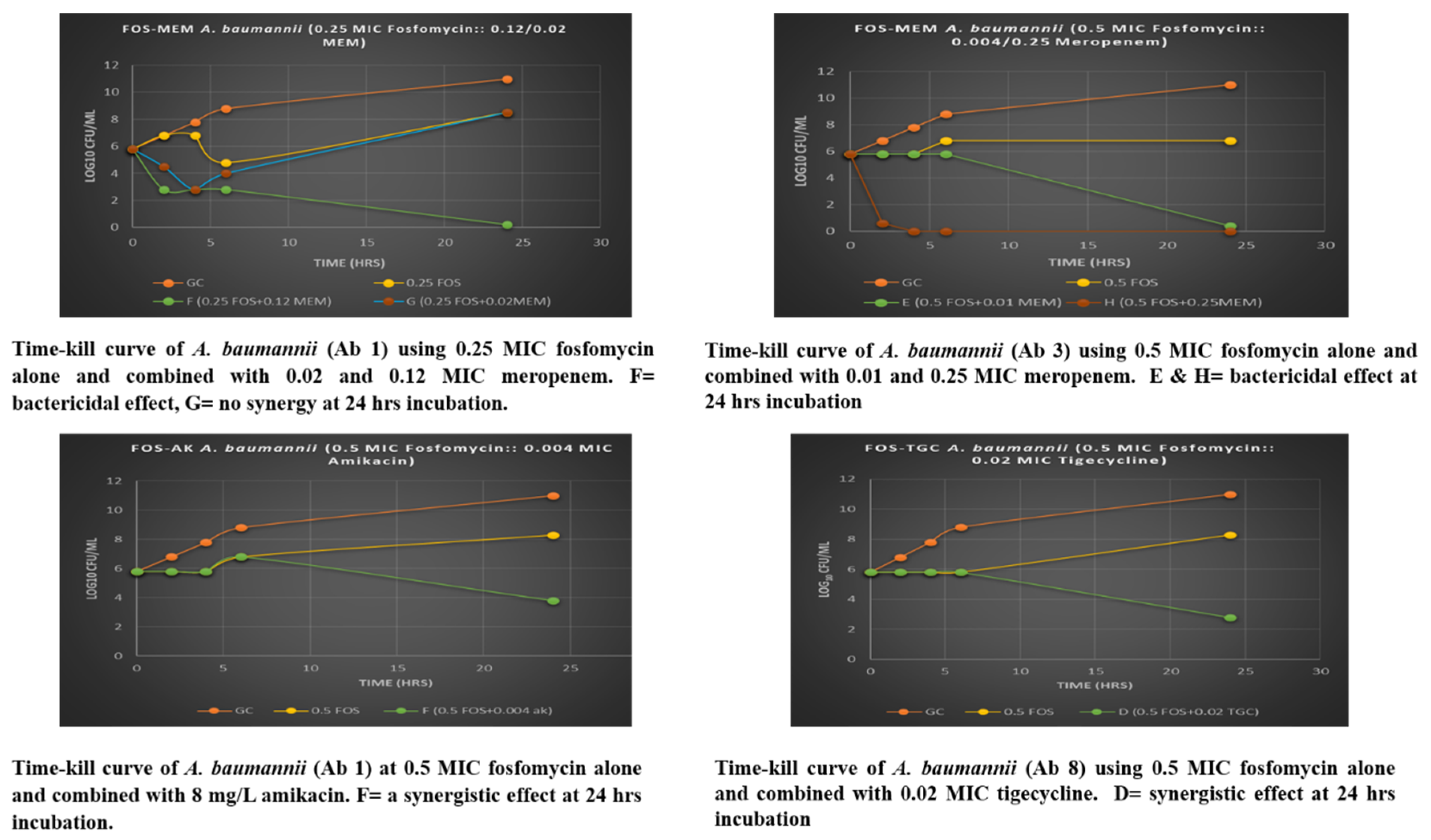
3.4. Assessment of Fosfomycin-Meropenem Interactions of A. baumannii by Time-Kill Assay
3.5. Synergy in PDR Strains
3.6. Identification of STs
3.7. Antibiotic Resistance Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Antunes, L.C.S.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Al Samawi, M.S.; Khan, F.Y.; Eldeeb, Y.; Almaslamani, M.; Alkhal, A.; Alsoub, H.; Ghadban, W.; Howady, F.; Hashim, S. Acinetobacter Infections among Adult Patients in Qatar: A 2-Year Hospital-Based Study. Can. J. Infect. Dis. Med. Microbiol. 2016, 2016, e6873689. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Ju, Y.G.; Lee, H.J.; Yim, H.S.; Lee, M.G.; Sohn, J.W.; Yoon, Y.K. In vitro synergistic antimicrobial activity of a combination of meropenem, colistin, tigecycline, rifampin, and ceftolozane/tazobactam against carbapenem-resistant Acinetobacter baumannii. Sci. Rep. 2022, 12, 7541. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Vouloumanou, E.K.; Samonis, G.; Vardakas, K.Z. Fosfomycin. Clin. Microbiol. Rev. 2016, 29, 321–347. [Google Scholar] [CrossRef]
- Sastry, S.; Clarke, L.G.; Alrowais, H.; Querry, A.M.; Shutt, K.A.; Doi, Y. Clinical Appraisal of Fosfomycin in the Era of Antimicrobial Resistance. Antimicrob. Agents Chemother. 2015, 59, 7355–7361. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Test for Bacteria That Grow Aerobically, 10th ed.; CLSI document M07-A10; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Rizvi, M.; Ahmed, J.; Khan, F.; Shukla, I.; Malik, A. Assessment of combination therapy by time kill curve analysis and chequerboard assay for treatment of multi-drug resistant Pseudomonas aeruginosa isolates. J. Glob. Antimicrob. Resist. 2013, 1, 103–108. [Google Scholar] [CrossRef]
- Garcia, L. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Clinical Microbiology Procedures Handbook: Update 2007; American Society for Microbiology: Washington, DC, USA, 2012. [Google Scholar]
- Zaidan, N.; Hornak, J.P.; Reynoso, D. Extensively Drug-Resistant Acinetobacter baumannii Nosocomial Pneumonia Successfully Treated with a Novel Antibiotic Combination. Antimicrob. Agents Chemother. 2021, 65, e00924-21. [Google Scholar] [CrossRef]
- Oo, C.; Sy, S.K.B. Fixed-dose combinations: A potential means to boost drug development for selected drugs. Drug Discov. Today 2018, 23, 457–459. [Google Scholar] [CrossRef]
- AL-Quraini, M.; Rizvi, M.; AL-Jabri, Z.; Sami, H.; AL-Muzahmi, M.; AL-Muharrmi, Z.; Taneja, N.; Al-Busaidi, I.; Soman, R. Assessment of In-Vitro Synergy of Fosfomycin with Meropenem, Amikacin and Tigecycline in Whole Genome Sequenced Extended and Pan Drug Resistant Klebsiella Pneumoniae: Exploring A Colistin Sparing Protocol. Antibiotics 2022, 11, 153. [Google Scholar] [CrossRef]
- Karakonstantis, S.; Ioannou, P.; Kofteridis, D.D. In search for a synergistic combination against pandrug-resistant A. baumannii; methodological considerations. Infection 2022, 50, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sazlly Lim, S.; Heffernan, A.J.; Roberts, J.A.; Sime, F.B. Pharmacodynamic Analysis of Meropenem and Fosfomycin Combination Against Carbapenem-Resistant Acinetobacter baumannii in Patients with Normal Renal Clearance: Can It Be a Treatment Option? Microb. Drug Resist. 2021, 27, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Singkham-In, U.; Chatsuwan, T. In vitro activities of carbapenems in combination with amikacin, colistin, or fosfomycin against carbapenem-resistant Acinetobacter baumannii clinical isolates. Diagn. Microbiol. Infect. Dis. 2018, 91, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Lv, Y.; Yuan, M.; Hu, X.; Nie, T.; Yang, X.; Li, G.; Pang, J.; Zhang, J.; Li, C.; et al. Genetic basis of high level aminoglycoside resistance in Acinetobacter baumannii from Beijing, China. Acta Pharm. Sin. B 2014, 4, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Kishk, R.; Soliman, N.; Nemr, N.; Eldesouki, R.; Mahrous, N.; Gobouri, A.; Azab, E.; Anani, M. Prevalence of Aminoglycoside Resistance and Aminoglycoside Modifying Enzymes in Acinetobacter baumannii Among Intensive Care Unit Patients, Ismailia, Egypt. Infect. Drug Resist. 2021, 14, 143–150. [Google Scholar] [CrossRef]
- Xu, C.; Bilya, S.R.; Xu, W. adeABC efflux gene in Acinetobacter baumannii. New Microbes New Infect. 2019, 30, 100549. [Google Scholar] [CrossRef] [PubMed]
- Leite, G.C.; Oliveira, M.S.; Perdigão-Neto, L.V.; Rocha, C.K.D.; Guimarães, T.; Rizek, C.; Levin, A.S.; Costa, S.F. Antimicrobial Combinations against Pan-Resistant Acinetobacter baumannii Isolates with Different Resistance Mechanisms. PLoS ONE 2016, 11, e0151270. [Google Scholar] [CrossRef]
- Montgomery, A.B.; Rhomberg, P.R.; Abuan, T.; Walters, K.A.; Flamm, R.K. Potentiation Effects of Amikacin and Fosfomycin against Selected Amikacin-Nonsusceptible Gram-Negative Respiratory Tract Pathogens. Antimicrob. Agents Chemother. 2014, 58, 3714–3719. [Google Scholar] [CrossRef]
- Martinez-Martinez, L.; Rodriguez, G.; Pascual, A.; Suárez, A.I.; Perea, E.J. In-vitro activity of antimicrobial agent combinations against multiresistant Acinetobacter baumannii. J. Antimicrob. Chemother. 1996, 38, 1107–1108. [Google Scholar] [CrossRef]
- Fan, B.; Guan, J.; Wang, X.; Cong, Y. Activity of Colistin in Combination with Meropenem, Tigecycline, Fosfomycin, Fusidic Acid, Rifampin or Sulbactam against Extensively Drug-Resistant Acinetobacter baumannii in a Murine Thigh-Infection Model. PLoS ONE 2016, 11, e0157757. [Google Scholar] [CrossRef]
- Bian, X.; Liu, X.; Chen, Y.; Chen, D.; Li, J.; Zhang, J. Dose Optimization of Colistin Combinations against Carbapenem-Resistant Acinetobacter baumannii from Patients with Hospital-Acquired Pneumonia in China by Using an In Vitro Pharmacokinetic/Pharmacodynamic Model. Antimicrob. Agents Chemother. 2019, 63, e01989-18. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G.R.; Lehár, J.; Keith, C.T. Multi-target therapeutics: When the whole is greater than the sum of the parts. Drug Discov. Today 2007, 12, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Maifiah, M.H.M.; Creek, D.J.; Nation, R.L.; Forrest, A.; Tsuji, B.T.; Velkov, T.; Li, J. Untargeted metabolomics analysis reveals key pathways responsible for the synergistic killing of colistin and doripenem combination against Acinetobacter baumannii. Sci. Rep. 2017, 7, 45527. [Google Scholar] [CrossRef] [PubMed]
- Gaiarsa, S.; Batisti Biffignandi, G.; Esposito, E.P.; Castelli, M.; Jolley, K.A.; Brisse, S.; Sassera, D.; Zarrilli, R. Comparative Analysis of the Two Acinetobacter baumanniiMultilocus Sequence Typing (MLST) Schemes. Front. Microbiol. 2019, 10, 930. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Isolate | MIC Value (mg/L) | |||||
|---|---|---|---|---|---|---|
| Fosfomycin | Meropenem | Tigecycline | Amikacin | Colistin | ||
| AD * | BMD # | BMD # | BMD # | BMD # | BMD # | |
| Ab1 (PDR) | 128 | 128 | 16 | 2 | >512 | 16 |
| Ab 2 | 128 | 64 | 32 | 2 | >512 | 0.5 |
| Ab 3 | 128 | ≤128 | 32 | 4 | >512 | 1 |
| Ab 4 | 32 | 64 | 32 | 4 | >512 | 1 |
| Ab 5 | 32 | 64 | 32 | 2 | >512 | 1 |
| Ab 6 | ≤128 | 128 | 32 | 4 | >512 | 1 |
| Ab 7 | 64 | 32 | 32 | 4 | >512 | 0.25 |
| Ab 8 | 64 | 64 | 64 | ≤0.5 | ≤8 | ≤1 |
| Isolate | Fos MIC (mg/L) | Fold Decline | MEM MIC (mg/L) | Fold Decline | FICI (x̄) | ||
|---|---|---|---|---|---|---|---|
| Alone | Combined with MEM | Alone | Combined with FOS | ||||
| Ab1 | 128 | 32 | 4 | 16 | 4 | 4 | 0.50 (S) |
| Ab2 | 64 | 1 | ≥64 | 32 | 16 | 2 | 0.52 (PS) |
| Ab3 | 64 | 16 | 4 | 32 | 8 | 4 | 0.50 (S) |
| Ab4 | 64 | 16 | 4 | 32 | 8 | 4 | 0.50 (S) |
| Ab5 | 64 | 16 | 4 | 32 | 8 | 4 | 0.50 (S) |
| Ab6 | 128 | 32 | 4 | 32 | 8 | 4 | 0.50 (S) |
| Ab7 | 32 | 8 | 4 | 32 | 8 | 4 | 0.50 (S) |
| Ab8 | 64 | 16 | 4 | 64 | 16 | 4 | 0.50 (S) |
| FOS MIC (mg/L) | AK MIC (mg/L) | ||||||
| alone | Combined with AK | alone | Combined with FOS | ||||
| Ab1 | 128 | 128 | 0 | >1024 | ≤8 | ≥256 | 1.00 (IN) |
| Ab2 | 128 | 128 | 0 | >1024 | ≤8 | ≥256 | 1.00 (IN) |
| Ab3 | 128 | 128 | 0 | >1024 | ≤8 | ≥256 | 1.00 (IN) |
| Ab4 | 64 | 64 | 0 | 1024 | ≤4 | ≥256 | 1.00 (IN) |
| Ab5 | 64 | 64 | 0 | 1024 | ≤4 | ≥256 | 1.00 (IN) |
| Ab6 | 128 | 128 | 0 | 1024 | ≤4 | ≥256 | 1.00 (IN) |
| Ab7 | 32 | 32 | 0 | >1024 | ≤4 | ≥512 | 1.00 (IN) |
| MEM MIC (mg/L) | CL MIC (mg/L) | ||||||
| alone | Combined with COL | alone | Combined with MEM | ||||
| Ab 1 | 16 | 0.5 | 32 | 8 | ≤1 | ≥8 | 0.16 (S) |
| Ab 2 | 32 | 4 | 8 | ≤1 | 0.25 | 4 | 0.37 (S) |
| Ab 3 | 32 | 8 | 4 | 1 | 0.25 | 4 | 0.50 (S) |
| FOS MIC (mg/L) | TGC MIC (mg/L) | ||||||
| alone | Combined with TGC | alone | Combined with FOS | ||||
| Ab 1 | 128 | ≤2 | ≥64 | 2 | 1 | 2 | 0.52 |
| Ab 2 | 128 | ≤4 | ≥32 | 2 | 1 | 2 | 0.53 |
| Ab 3 | 128 | ≤4 | ≥32 | 4 | 2 | 2 | 0.53 |
| Ab 4 | 64 64 | ≤1 32 | ≥64 2 | 4 4 | 2 0.25 | 2 16 | 0.52 0.56 |
| Ab 5 | 64 64 | ≤1 32 | ≥64 2 | 2 2 | 1 ≤0.06 | 2 ≥32 | 0.52 0.53 |
| Ab 6 | 128 128 | ≤1 64 | ≥128 2 | 4 4 | 2 ≤0.06 | 2 ≥64 | 0.51 0.52 |
| Ab 7 | 32 | ≤1 | ≥32 | 4 | 2 | 2 | 0.53 |
| Combination | [(FOS) + (MEM)] mg/L | 6-h Effect | 24-h Effect | 24-h log10 Killing ∆ 24 h | Checkerboard |
|---|---|---|---|---|---|
| Fosfomycin + Meropenem | Isolate Ab 3 | ||||
| 0.25 FOS + 0.02 MEM | (32, 2) | Synergy | None | >+2 | Growth |
| 0.25 FOS + 0.12 MEM | (32, 8) | Bactericidal | Bactericidal | >−5 | Synergy |
| 0.5 FOS + 0.01 MEM | (64, 0.5) | None | Bactericidal | >−5 | Growth |
| 0.5 FOS + 0.25 MEM | (64, 16) | Bactericidal | Bactericidal | −6 | Partial synergy |
| Fosfomycin + Amikacin | Isolate Ab 1 | ||||
| 0.5 FOS + 0.004 AK | (64, 8) | None | Synergy | −2 | Growth |
| Fosfomycin + Tigecycline | Isolate Ab 4 | ||||
| 0.5 FOS + 0.02 TGC | (64, 0.06) | None | Bactericidal | −3 | Partial synergy |
| Sample | ST | gltA | gyrB | gdhB | recA | cpn60 | gpi | rpoD | Mismatches | Uncertainty | Depth | maxMAF |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ab1 | 2062 | 1 | 17 | 189 | 2 | 2 | 108 | 3 | 0 | - | 197.8256 | 0.016667 |
| Ab2 | 2063 | 1 | 17 | 3 | 2 | 2 | 108 | 3 | 0 | - | 187.7057 | 0.013793 |
| Ab3 | 1816 | 1 | 3 | 189 | 2 | 2 | 96 | 3 | 0 | - | 161.3086 | 0.015625 |
| Ab4 | 1962 | 1 | 3 | 189 | 2 | 2 | 140 | 3 | 0 | - | 234.2391 | 0.00885 |
| Ab5 | 1962 | 1 | 3 | 189 | 2 | 2 | 140 | 3 | 0 | - | 210.5916 | 0.011364 |
| Ab6 | 1806 | 1 | 3 | 189 | 2 | 2 | 97 | 3 | 0 | - | 179.503 | 0.012422 |
| Ab7 | 234 | 21 | 48 | 58 | 42 | 36 | 109 | 4 | 0 | - | 258.423 | 0.013263 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quraini, M.A.; Jabri, Z.A.; Sami, H.; Mahindroo, J.; Taneja, N.; Muharrmi, Z.A.; Busaidi, I.A.; Rizvi, M. Exploring Synergistic Combinations in Extended and Pan-Drug Resistant (XDR and PDR) Whole Genome Sequenced Acinetobacter baumannii. Microorganisms 2023, 11, 1409. https://doi.org/10.3390/microorganisms11061409
Quraini MA, Jabri ZA, Sami H, Mahindroo J, Taneja N, Muharrmi ZA, Busaidi IA, Rizvi M. Exploring Synergistic Combinations in Extended and Pan-Drug Resistant (XDR and PDR) Whole Genome Sequenced Acinetobacter baumannii. Microorganisms. 2023; 11(6):1409. https://doi.org/10.3390/microorganisms11061409
Chicago/Turabian StyleQuraini, Munawr AL, Zaaema AL Jabri, Hiba Sami, Jaspreet Mahindroo, Neelam Taneja, Zakariya AL Muharrmi, Ibrahim AL Busaidi, and Meher Rizvi. 2023. "Exploring Synergistic Combinations in Extended and Pan-Drug Resistant (XDR and PDR) Whole Genome Sequenced Acinetobacter baumannii" Microorganisms 11, no. 6: 1409. https://doi.org/10.3390/microorganisms11061409