
Tryptophanase Expressed by Salmonella Halts Breast Cancer Cell Growth In Vitro and Inhibits Production of Immunosuppressive Kynurenine

Abstract
:1. Introduction
2. Materials and Methods
3. Results
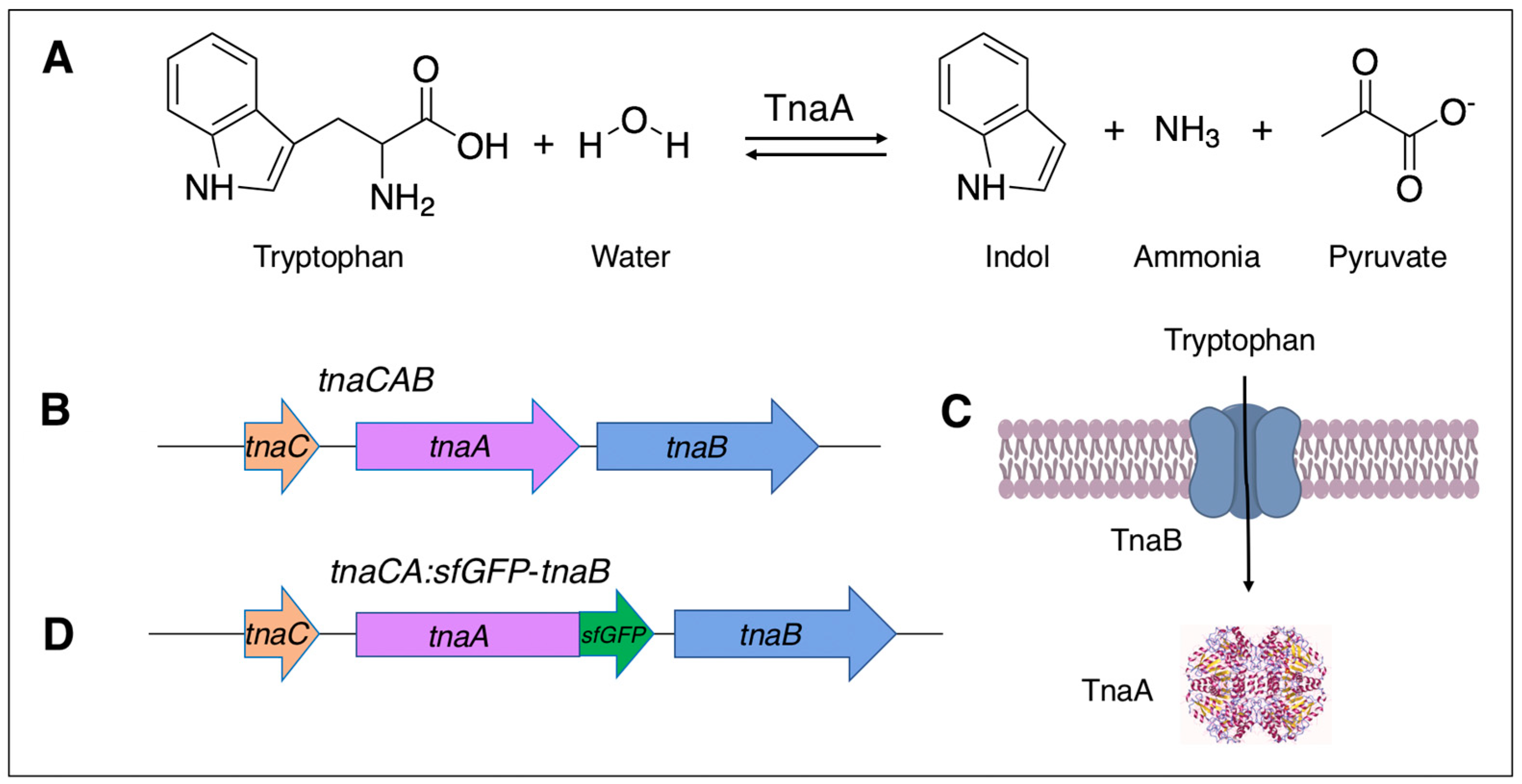
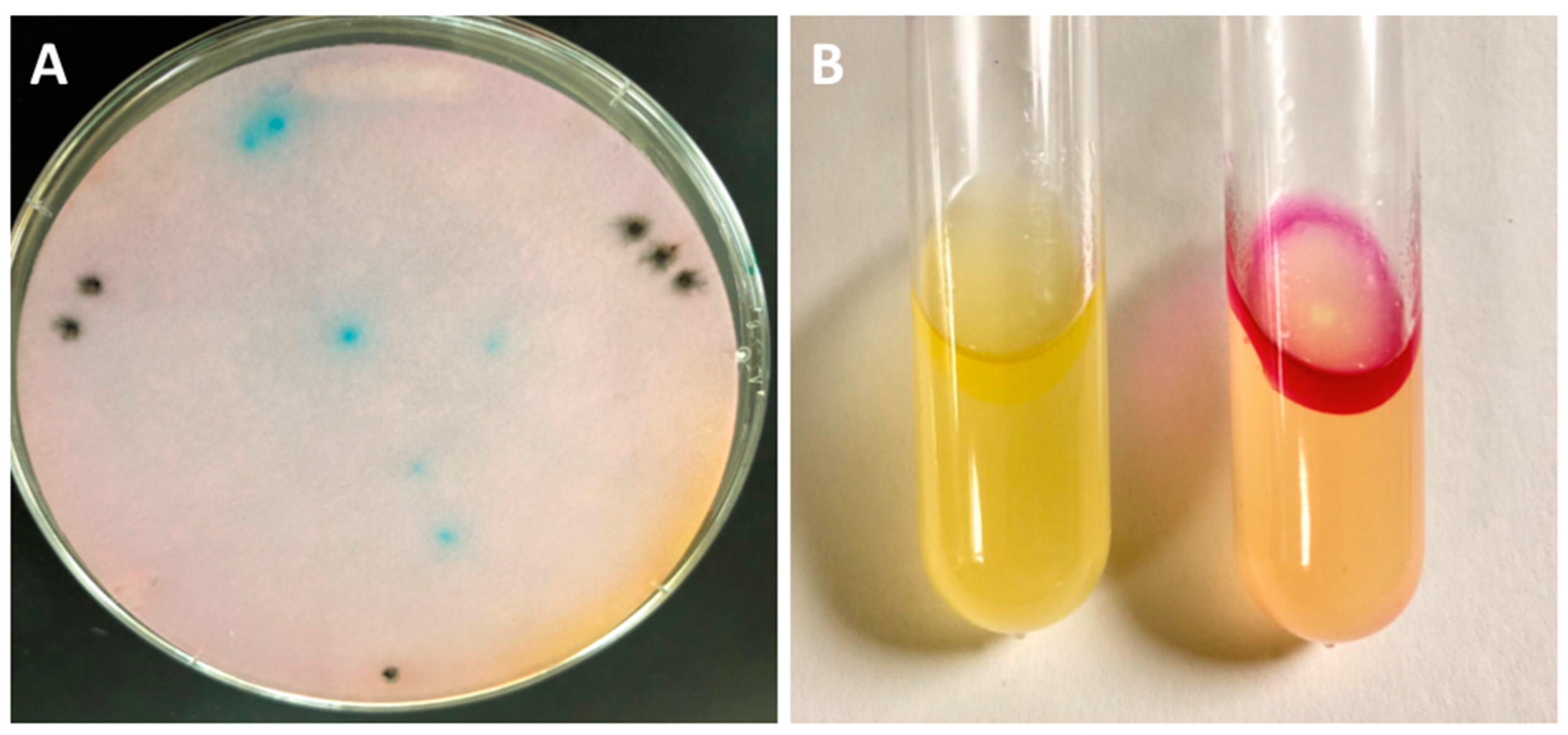
3.1. Cloning of Tryptophanase and Detecting Tryptophaase Activity in Salmonella VNP20009
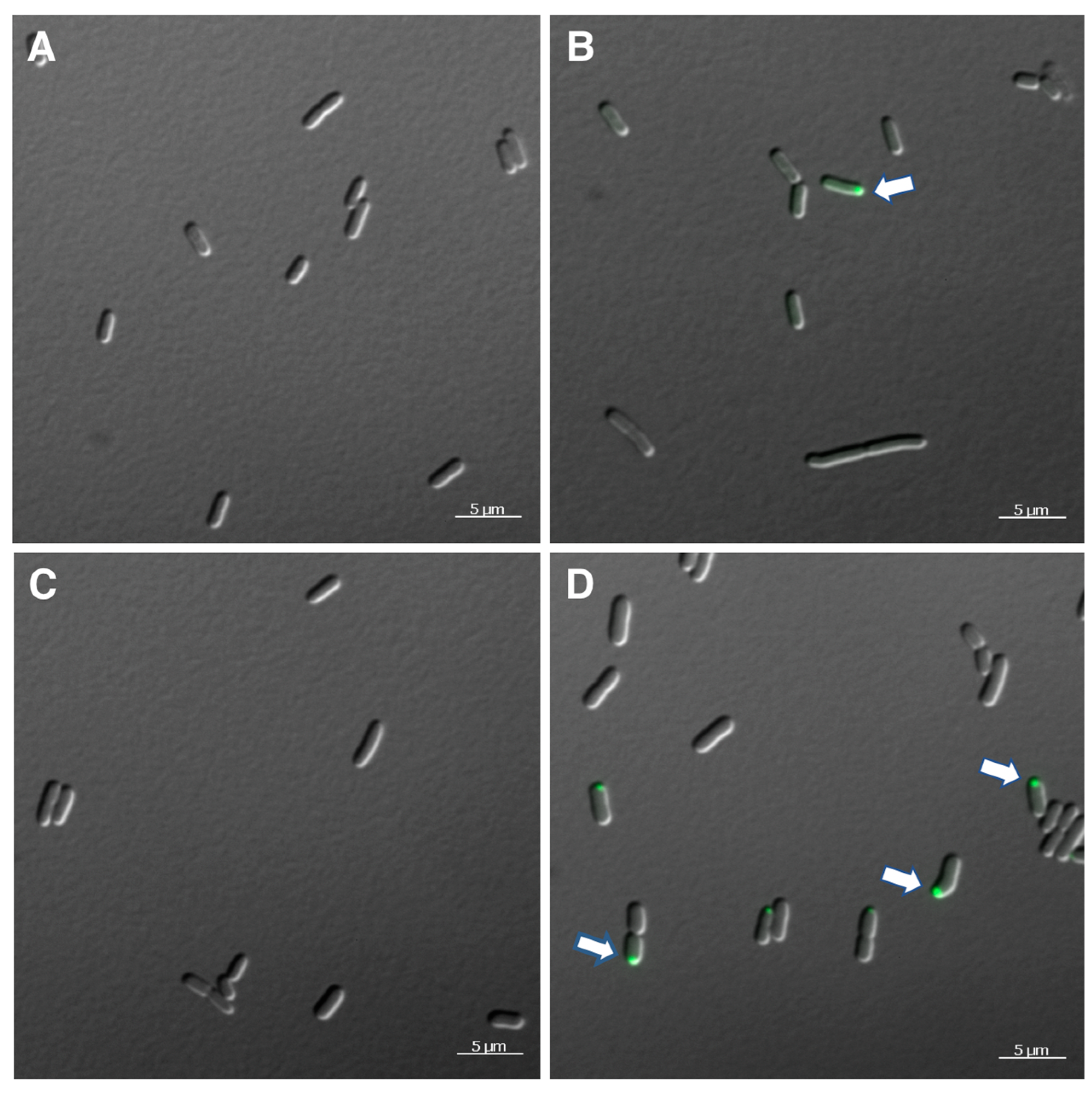
3.2. Intracellular Localization of Tryptophanase in Salmonella VNP20009
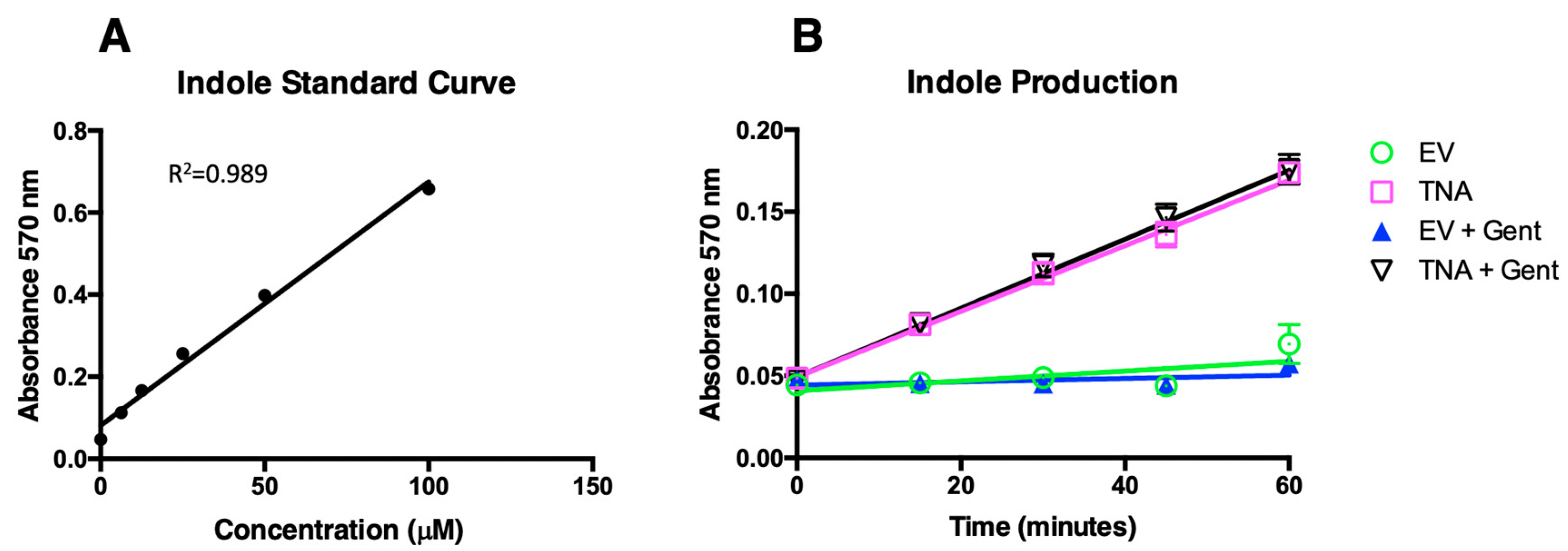
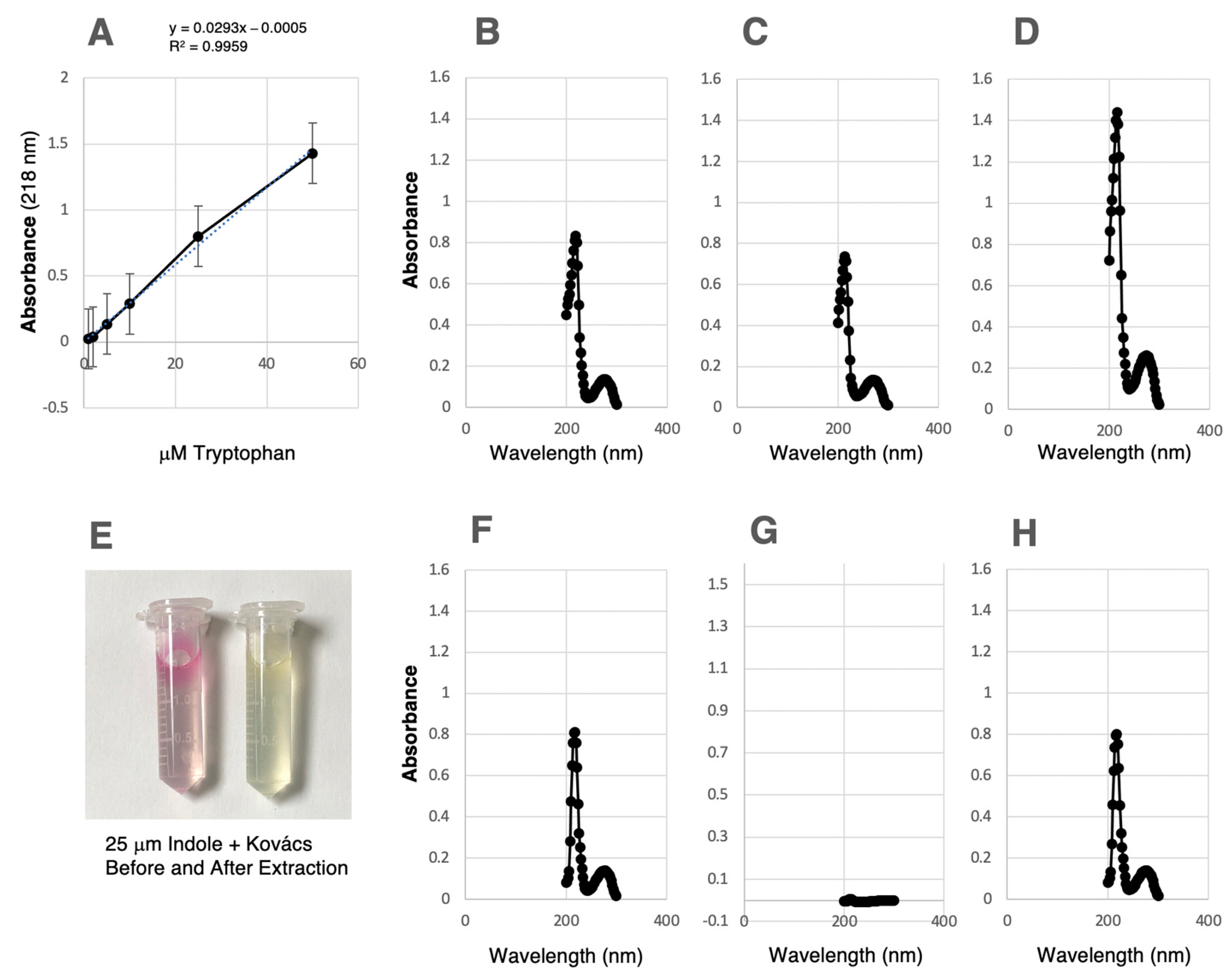
3.3. Quantitative Indole Determinations and Tryptophan Conversion by VNP20009-EV and VNP20009-tnaCAB
3.4. Separation of Indole from Tryptophan and Measurement of Tryptophan
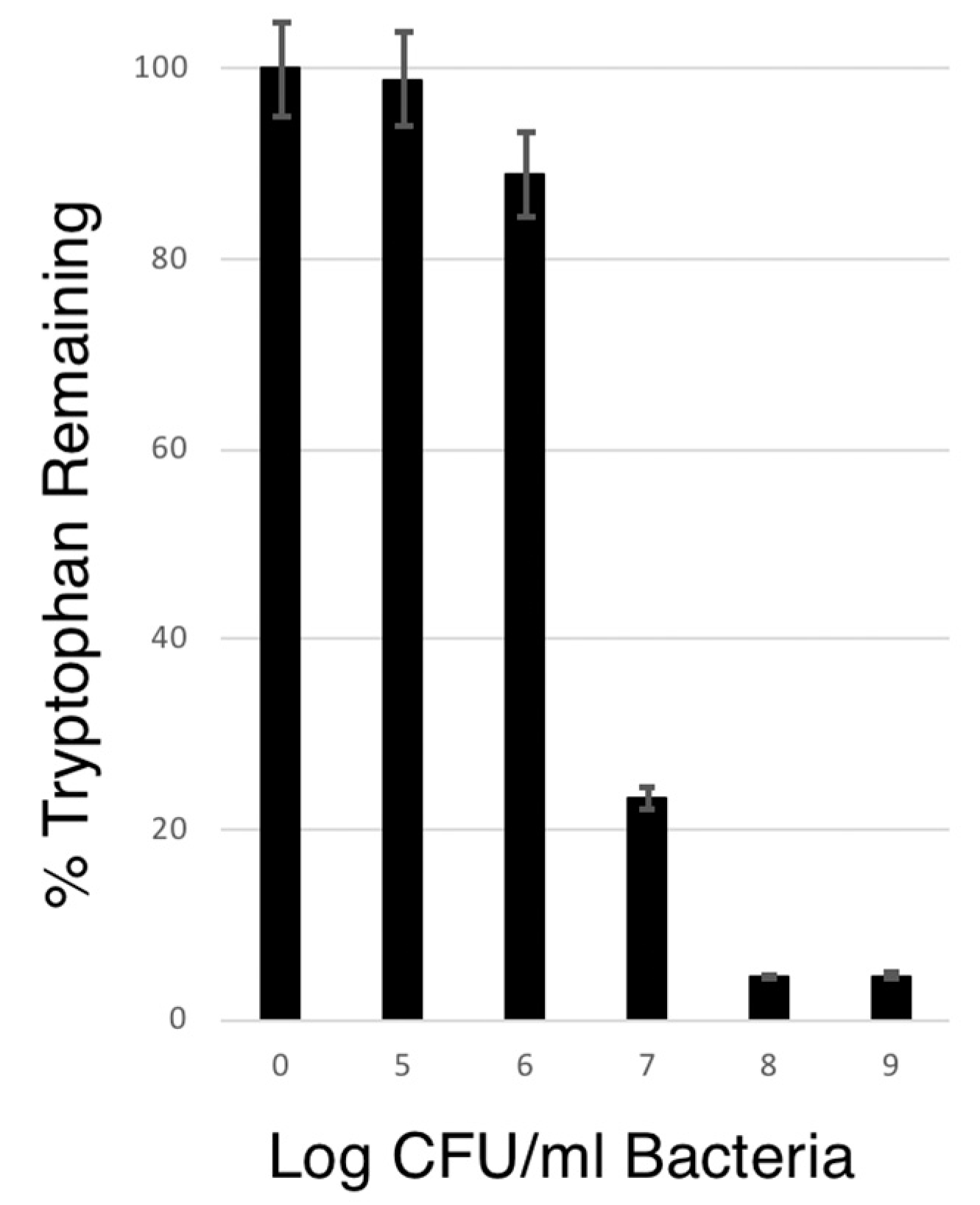
3.5. The Ability of VNP20009-tnaCAB to Deplete Tryptophan
3.6. Effect of Media Depletion of Tryptophan
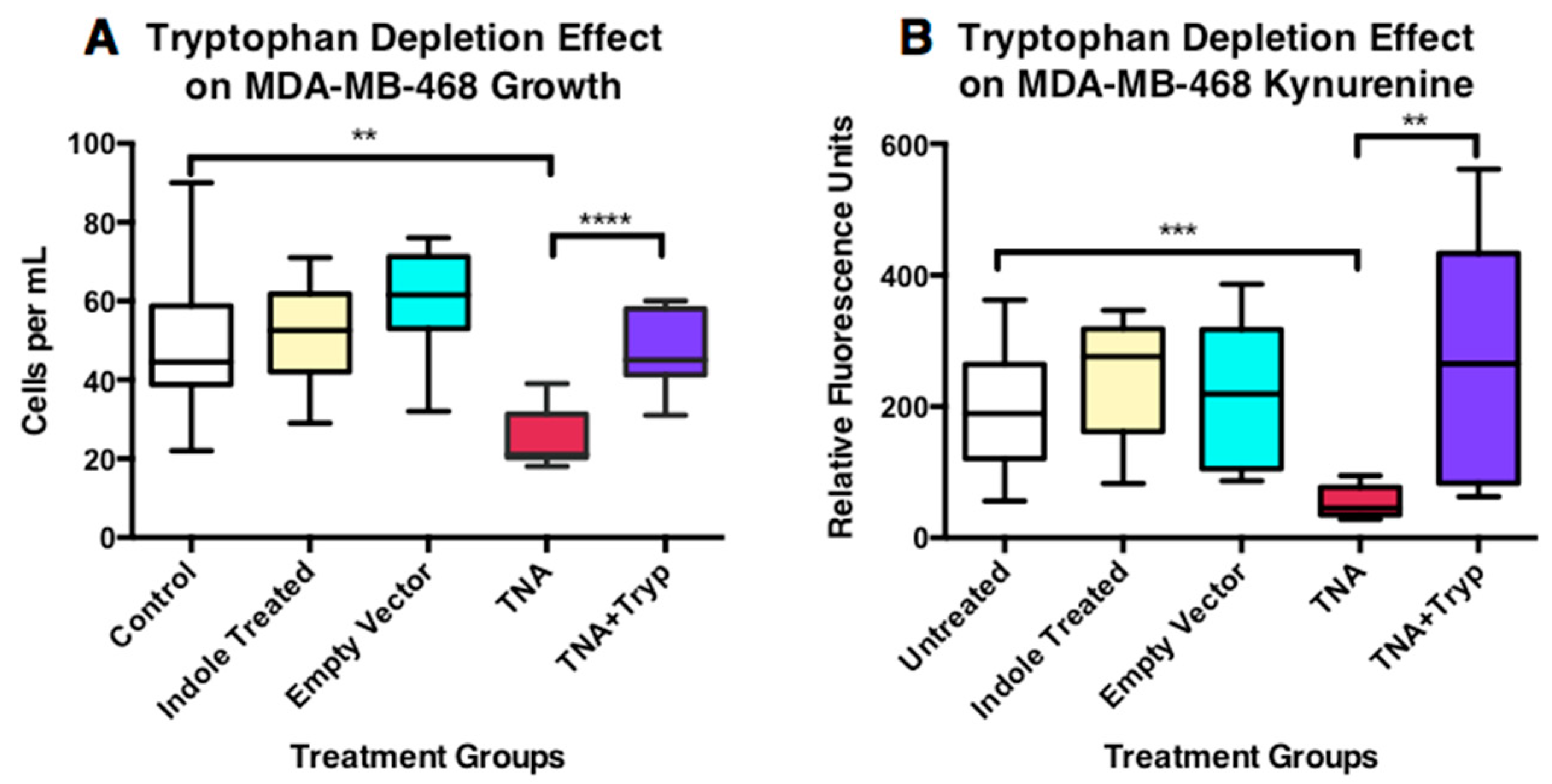
3.6.1. Tumor Cell Growth Alteration
3.6.2. Alteration in the Production of Kynurenine
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bröer, S. Amino Acid Transporters as Targets for Cancer Therapy: Why, Where, When, and How. Int. J. Mol. Sci. 2020, 21, 6156. [Google Scholar] [CrossRef] [PubMed]
- Fung, M.K.L.; Chan, G.C.-F. Drug-induced amino acid deprivation as strategy for cancer therapy. J. Hematol. Oncol. 2017, 10, 144. [Google Scholar] [CrossRef] [PubMed]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, H. Reprogramming of glucose, fatty acid and amino acid metabolism for cancer progression. Cell Mol. Life Sci. 2016, 73, 377–392. [Google Scholar] [CrossRef]
- Geck, R.; Toker, A. Nonessential amino acid metabolism in breast cancer. Adv. Biol. Regul. 2016, 62, 11–17. [Google Scholar] [CrossRef]
- Pokrovsky, V.S.; Chepikova, O.E.; Davydov, D.Z.; Zamyatnin, A.A., Jr.; Lukashev, A.N.; Lukasheva, E.V. Amino Acid Degrading Enzymes and their Application in Cancer Therapy. Curr. Med. Chem. 2019, 26, 446–464. [Google Scholar] [CrossRef]
- Dhankhar, R.; Gupta, V.; Kumar, S.; Kapoor, R.K.; Gulati, P. Microbial enzymes for deprivation of amino acid metabolism in malignant cells: Biological strategy for cancer treatment. Appl. Microbiol. Biotechnol. 2020, 104, 2857–2869. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, Q.; Zhou, H.; Zhang, M.; Shen, J.; Ju, D. Amino Acid Degrading Enzymes and Autophagy in Cancer Therapy. Front. Pharm. 2020, 11, 582587. [Google Scholar] [CrossRef]
- Timosenko, E.; Hadjinicolaou, A.V.; Cerundolo, V. Modulation of cancer-specific immune responses by amino acid degrading enzymes. Immunotherapy 2017, 9, 83–97. [Google Scholar] [CrossRef]
- Hou, D.-Y.; Muller, A.J.; Sharma, M.D.; DuHadaway, J.; Banerjee, T.; Johnson, M.; Mellor, A.L.; Prendergast, G.C.; Munn, D.H. Inhibition of indoleamine 2,3-dioxygenase in dendritic cells by stereoisomers of 1-methyl-tryptophan correlates with antitumor responses. Cancer Res. 2007, 67, 792–801. [Google Scholar] [CrossRef]
- Koblish, H.K.; Hansbury, M.J.; Bowman, K.J.; Yang, G.; Neilan, C.L.; Haley, P.J.; Burn, T.C.; Waeltz, P.; Sparks, R.B.; Yue, E.W.; et al. Hydroxyamidine inhibitors of indoleamine-2,3-dioxygenase potently suppress systemic tryptophan catabolism and the growth of IDO-expressing tumors. Mol. Cancer 2010, 9, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Opitz, C.A.; Patterson, L.F.S.; Mohapatra, S.R.; Dewi, D.L.; Sadik, A.; Platten, M.; Trump, S. The therapeutic potential of targeting tryptophan catabolism in cancer. Br. J. Cancer 2020, 122, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Brochez, L.; Chevolet, I.; Kruse, V. The rationale of indoleamine 2,3-dioxygenase inhibition for cancer therapy. Eur. J. Cancer 2017, 76, 167–182. [Google Scholar] [CrossRef]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug. Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef] [PubMed]
- Low, K.B.; Ittensohn, M.; Luo, X.; Zheng, L.-M.; King, I.; Pawelek, J.M.; Bermudes, D. Construction of VNP20009: A novel, genetically stable antibiotic-sensitive strain of tumor-targeting Salmonella for parenteral administration in humans. Methods Mol. Med. 2004, 90, 47–60. [Google Scholar] [PubMed]
- Forbes, N.S. Engineering the perfect (bacterial) cancer therapy. Nat. Rev. Cancer 2010, 10, 785–794. [Google Scholar] [CrossRef]
- Hoffman, R.M. Back to the Future: Are Tumor-Targeting Bacteria the Next-Generation Cancer Therapy? Methods Mol. Biol. 2015, 1317, 239–260. [Google Scholar]
- Zhou, S.; Gravekamp, C.; Bermudes, D.; Liu, K. Tumour-targeting bacteria engineered to fight cancer. Nat. Rev. Cancer 2018, 18, 727–743. [Google Scholar] [CrossRef]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F.; et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef]
- Cunningham, C.; Nemunaitis, J. A phase I trial of genetically modified Salmonella typhimurium expressing cytosine deaminase (TAPET-CD, VNP20029) administered by intratumoral injection in combination with 5-fluorocytosine for patients with advanced or metastatic cancer. Protocol no: CL-017. Version: 9 April 2001. Hum. Gene 2001, 12, 1594–1596. [Google Scholar]
- Heimann, D.M.; Rosenberg, S.A. Continuous intravenous administration of live genetically modified Salmonella typhimurium in patients with metastatic melanoma. J. Immunother. 2003, 26, 179–180. [Google Scholar] [CrossRef]
- Nemunaitis, J.; Cunningham, C.; Senzer, N.; Kuhn, J.; Cramm, J.; Litz, C.; Cavagnolo, R.; Cahill, A.; Clairmont, C.; Sznol, M. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene 2003, 10, 737–744. [Google Scholar] [CrossRef]
- Thamm, D.H.; Kurzman, I.D.; King, I.; Li, Z.; Sznol, M.; Dubielzig, R.R.; Vail, D.M.; Gregory MacEwen, E. Systemic administration of an attenuated, tumor-targeting Salmonella typhimurium to dogs with spontaneous neoplasia: Phase I evaluation. Clin. Cancer Res. 2005, 11, 4827–4834. [Google Scholar] [CrossRef]
- Kim, K.; Jeong, J.H.; Lim, D.; Hong, Y.; Lim, H.-J.; Kim, G.-J.; Shin, S.-R.; Lee, J.-J.; Yun, M.; A Harris, R.; et al. L-Asparaginase delivered by Salmonella typhimurium suppresses solid tumors. Mol. Oncolytics 2015, 2, 15007. [Google Scholar] [CrossRef]
- Tsai, S.P.; Hartin, R.J.; Ryu, J.-I. Transformation in restriction-deficient Salmonella typhimurium LT2. J. Gen. Microbiol. 1989, 135, 2561–2567. [Google Scholar] [CrossRef]
- Broadway, K.M.; Modise, T.; Jensen, R.V.; Scharf, B.E. Complete genome sequence of Salmonella enterica serovar Typhimurium VNP20009, a strain engineered for tumor targeting. J. Biotechnol. 2014, 192, 177–178. [Google Scholar] [CrossRef]
- Murray, S.R.; Bermudes, D.; de Felipe, S.W.; Low, K.B. Extragenic suppressors of msbB- growth defects in Salmonella. J. Bacteriol. 2001, 183, 5554–5561. [Google Scholar] [CrossRef]
- Amann, E.; Ochs, B.; Abel, K.-J. Tightly regulated tac promoter vectors useful for the expression of unfused and fused proteins in Escherichia coli. Gene 1988, 69, 301–315. [Google Scholar] [CrossRef]
- O’Callaghan, D.; Charbit, A. High efficiency transformation of Salmonella typhimurium and Salmonella typhi by electroporation. Mol. Gen. Genet. 1990, 223, 156–158. [Google Scholar] [CrossRef]
- Quintero, D.; Carrafa, J.; Vincent, L.; Bermudes, D. EGFR-targeted Chimeras of Pseudomonas ToxA released into the extracellular milieu by attenuated Salmonella selectively kill tumor cells. Biotechnol. Bioeng. 2016, 113, 2698–2711. [Google Scholar] [CrossRef]
- Elsinghorst, E.A. Measurement of invasion by gentamicin resistance. Methods Enzym. 1994, 236, 405–420. [Google Scholar]
- Dewi, D.L.; Mohapatra, S.R.; Cabañes, S.B.; Adam, I.; Patterson, L.F.S.; Berdel, B.; Kahloon, M.; Thürmann, L.; Loth, S.; Heilmann, K.; et al. Suppression of indoleamine-2,3-dioxygenase 1 expression by promoter hypermethylation in ER-positive breast cancer. Oncoimmunology 2017, 6, e12744772017. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Young, K.D. A new suite of tnaA mutants suggests that Escherichia coli tryptophanase is regulated by intracellular sequestration and by occlusion of its active site. BMC Microbiol. 2015, 15, 14. [Google Scholar] [CrossRef]
- Stewart, K.L.; Stewart, A.M.; Bobik, T.A. Prokaryotic Organelles: Bacterial Microcompartments in E. coli and Salmonella. EcoSal Plus 2020, 9. [Google Scholar] [CrossRef]
- Routy, J.-P.; Routy, B.; Graziani, G.M.; Mehraj, V. The Kynurenine Pathway Is a Double-Edged Sword in Immune-Privileged Sites and in Cancer: Implications for Immunotherapy. Int. J. Tryptophan Res. 2016, 9, 67–77. [Google Scholar] [CrossRef]
- Estrella, V.; Chen, T.; Lloyd, M.; Wojtkowiak, J.; Cornnell, H.H.; Ibrahim-Hashim, A.; Bailey, K.; Balagurunathan, Y.; Rothberg, J.M.; Sloane, B.F.; et al. Acidity generated by the tumor microenvironment drives local invasion. Cancer Res. 2013, 73, 1524–1535. [Google Scholar] [CrossRef]
- Nuidate, T.; Tansila, N.; Chomchuen, P.; Phattaranit, P.; Eangchuan, S.; Vuddhakul, V. Characterization of tryptophanase from Vibrio cholerae. Appl. Biochem. Biotechnol. 2015, 175, 243–252. [Google Scholar] [CrossRef]
- Mehraj, V.; Routy, J.-P. Tryptophan Catabolism in Chronic Viral Infections: Handling Uninvited Guests. Int. J. Tryptophan Res. 2015, 8, 41–48. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Kimura, A.; Nakahama, T.; Chinen, I.; Masuda, K.; Nohara, K.; Fujii-Kuriyama, Y.; Kishimoto, T. Aryl hydrocarbon receptor negatively regulates dendritic cell immunogenicity via a kynurenine-dependent mechanism. Proc. Natl. Acad. Sci. USA 2010, 107, 19961–19966. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by tryptophan catabolism. Cell. Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by kynurenines. Adv. Exp. Med. Biol. 2003, 527, 183–190. [Google Scholar] [PubMed]
- Sharma, B.; Singh, S.; Kanwar, S.S. L-methionase: A therapeutic enzyme to treat malignancies. Biomed Res. Int. 2014, 2014, 506287. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-I.; Liao, J.C.; Kuo, L. Arginase modulates nitric oxide production in activated macrophages. Am. J. Physiol. Heart Circ. Physiol. 1998, 274, H342–H348. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.I.; Liao, J.C.; Kuo, L. Macrophage arginase promotes tumor cell growth and suppresses nitric oxide-mediated tumor cytotoxicity. Cancer Res. 2001, 61, 1100–1106. [Google Scholar]
- Liu, S.; Monks, N.R.; Hanes, J.W.; Begley, T.P.; Yu, H.; Moscow, J.A. Sensitivity of breast cancer cell lines to recombinant thiaminase I. Cancer Chemother. Pharm. 2010, 66, 171–179. [Google Scholar] [CrossRef]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Tumor-targeted Salmonella as a novel anticancer vector. Cancer Res. 1997, 57, 4537–4544. [Google Scholar]
- Low, K.B.; Ittensohn, M.; Le, T.; Platt, J.; Sodi, S.; Amoss, M.; Ash, O.; Carmichael, E.; Chakraborty, A.; Fischer, J.; et al. Lipid A mutant Salmonella with suppressed virulence and TNFα induction retain tumor-targeting in vivo. Nat. Biotechnol. 1999, 17, 37–41. [Google Scholar] [CrossRef]
- Quintero, D.; Carrafa, J.; Vincent, L.; Kim, H.J.; Wohlschlegel, J.; Bermudes, D. Co-Expression of a chimeric protease inhibitor secreted by a tumor-targeted Salmonella protects therapeutic proteins from proteolytic degradation. J. Microbiol. Biotechnol. 2018, 28, 2079–2094. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Q.; Zhang, Z.; Cheng, W.; Cao, W.; Jiang, C.; Han, C.; Li, J.; Hua, Z. Chloroquine enhanced the anticancer capacity of VNP20009 by inhibiting autophagy. Sci. Rep. 2016, 6, 29774. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer # | Primer Name | Primer Sequence 5’-3’ | Primer Restriction Enzyme Site(s) (Underlined in Sequence) |
|---|---|---|---|
| 1 | ptnaCA_KpnI_F1 | GATCGGTACCAGGAGGAATTCACCATGAATATCTTACATATATGTGTGACCTCAA | KpnI |
| 2 | ptnaB _XbaI_R1 | GATCTCTAGAGAAGGATTTAGCCAAATTTAGGTAACAC | XbaI |
| 3 | pAraF1 | ACCTGACGCTTTTTATCGCA | |
| 4 | pTrcR2 | CCGCCAGGCAAATTCTGT | |
| 5 | For_1 | TTAGCCGAGTCAGTGAAAAA | |
| 6 | For_2 | AGACGACAGCTTCTTTGATGTG | |
| 7 | For_3 | GCCACTCTCTTACCCTACATCC | |
| 8 | For_4 | CACTGCGGGAACGTCTTACT | |
| 9 | For_5 | CAATCTTATTCCGGCGATTG | |
| 10 | Rev_1 | AATCAGTACCGGAATATAGATTTGC | |
| 11 | Rev_2 | CACATCAAAGAAGCTGTCGTCT | |
| 12 | Rev_3 | CACATCAAAGAAGCTGTCGTCT | |
| 13 | Rev_4 | ATATTGCCGTGGAAACCAAA- | |
| 14 | Rev_5 | GCCAAATTTAGGTAACACGTTAAA | |
| 15 | ptnaA_nostop_R1 | GCCAAATTTAGG TAACACGTTAAAGA | |
| 16 | ptnaA_nostop_ WsfGFP_F1 | TCTTTAACGTGTTACCTAAATTTGGCATGGTGAGCAAGGGCGA | |
| 17 | and psfGFP stopPacIXbaI_R1 | GACTTCTAGAGACTTTAATTAATTACTTGTACAGCTCGTCCATGC | XbaI, PacI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hababag, E.A.C.; Cauilan, A.; Quintero, D.; Bermudes, D. Tryptophanase Expressed by Salmonella Halts Breast Cancer Cell Growth In Vitro and Inhibits Production of Immunosuppressive Kynurenine. Microorganisms 2023, 11, 1355. https://doi.org/10.3390/microorganisms11051355
Hababag EAC, Cauilan A, Quintero D, Bermudes D. Tryptophanase Expressed by Salmonella Halts Breast Cancer Cell Growth In Vitro and Inhibits Production of Immunosuppressive Kynurenine. Microorganisms. 2023; 11(5):1355. https://doi.org/10.3390/microorganisms11051355
Chicago/Turabian StyleHababag, Eljoie Anice Cada, Allea Cauilan, David Quintero, and David Bermudes. 2023. "Tryptophanase Expressed by Salmonella Halts Breast Cancer Cell Growth In Vitro and Inhibits Production of Immunosuppressive Kynurenine" Microorganisms 11, no. 5: 1355. https://doi.org/10.3390/microorganisms11051355