
Structure of Microbial Communities and Biological Activity in Tundra Soils of the Euro-Arctic Region (Rybachy Peninsula, Russia)
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of the Climate and Soils
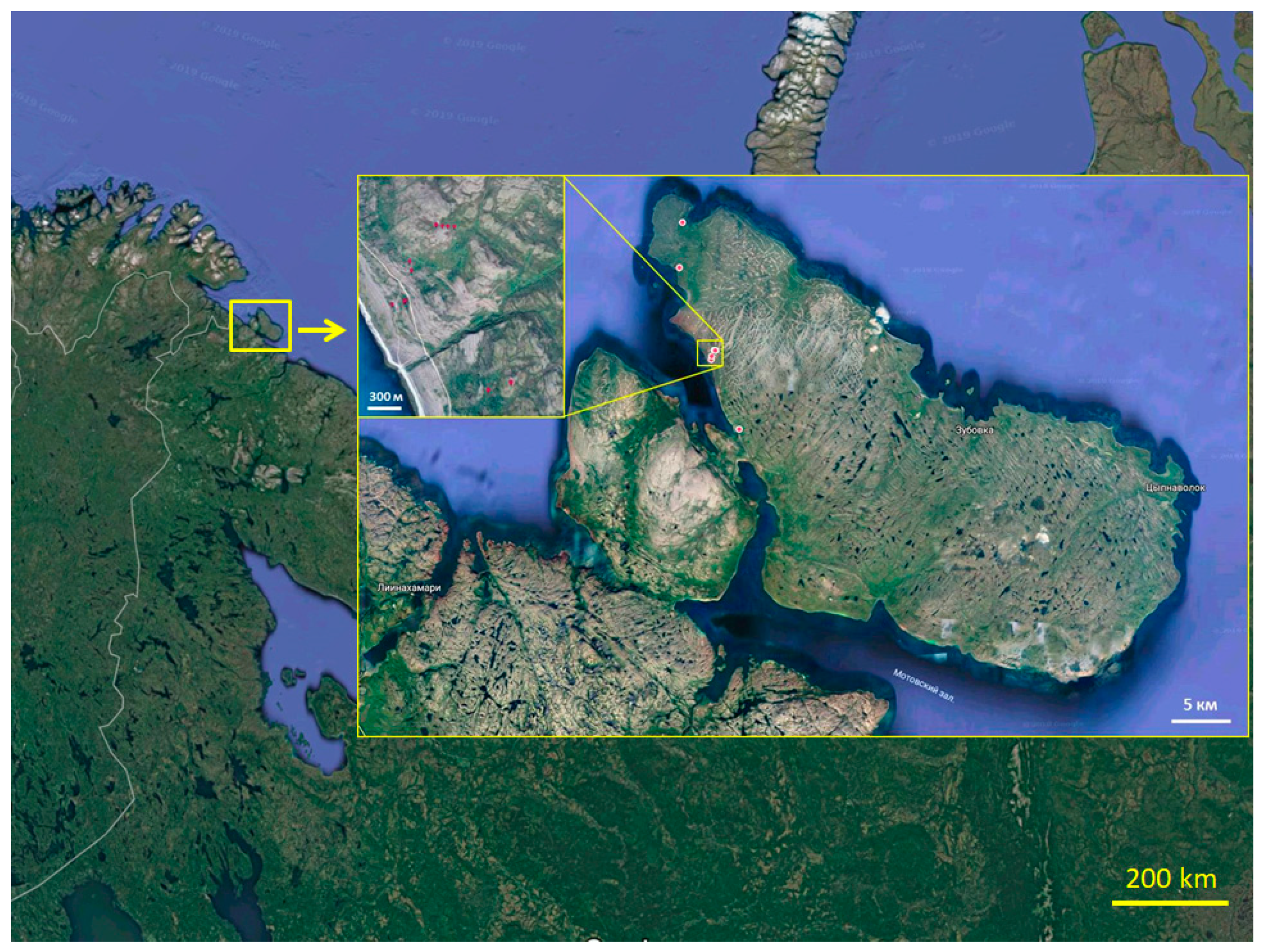
2.2. Sampling
2.3. Prokaryotic Biomass
2.4. Fungal Biomass
2.5. Number of Culturable Microorganisms
2.6. Enzymatic Activity of Soils
2.7. Soil pH and Total Organic Carbon (TOC)
2.8. Statistical Analysis
3. Results and Discussion
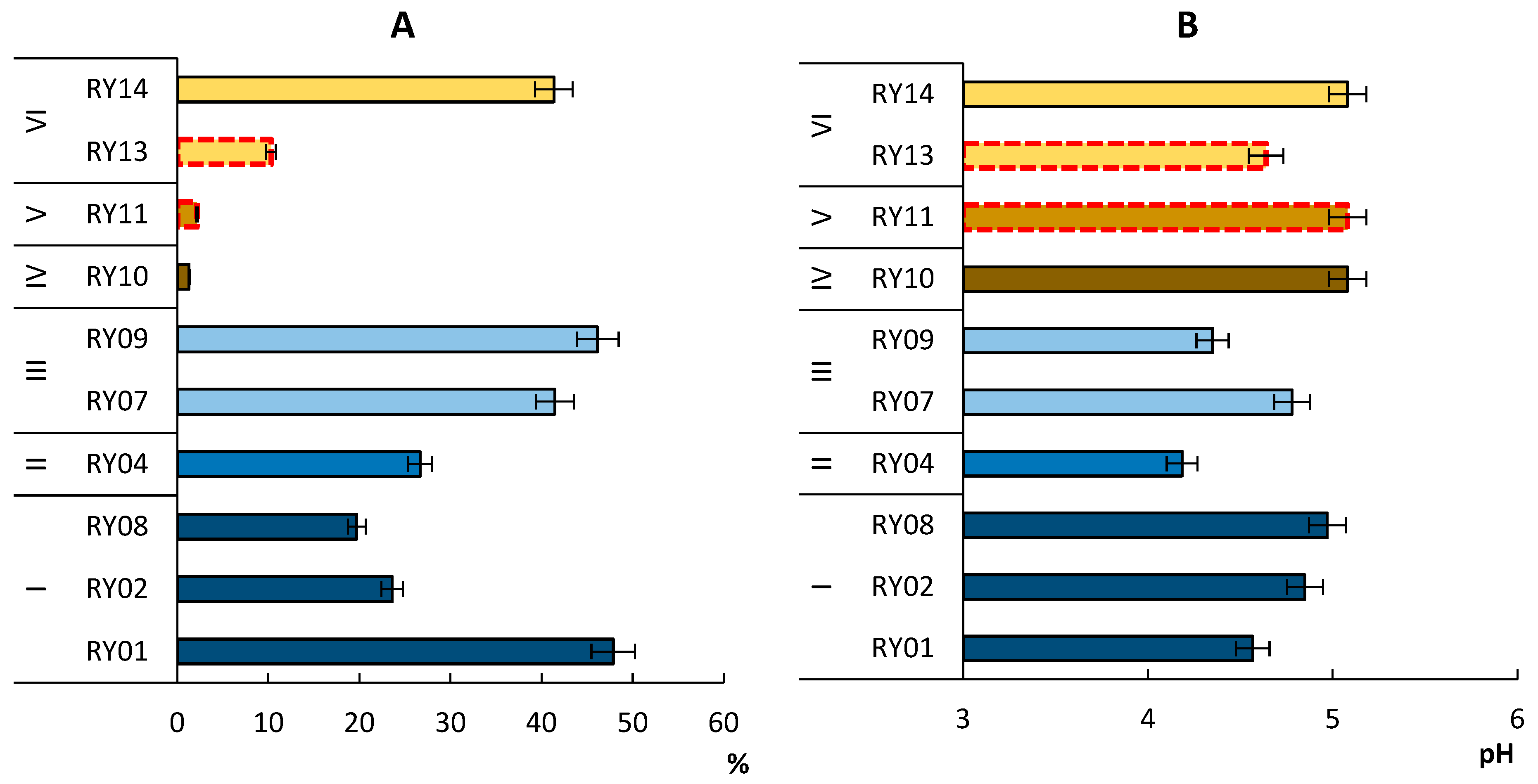
3.1. Soil pH and TOC Content
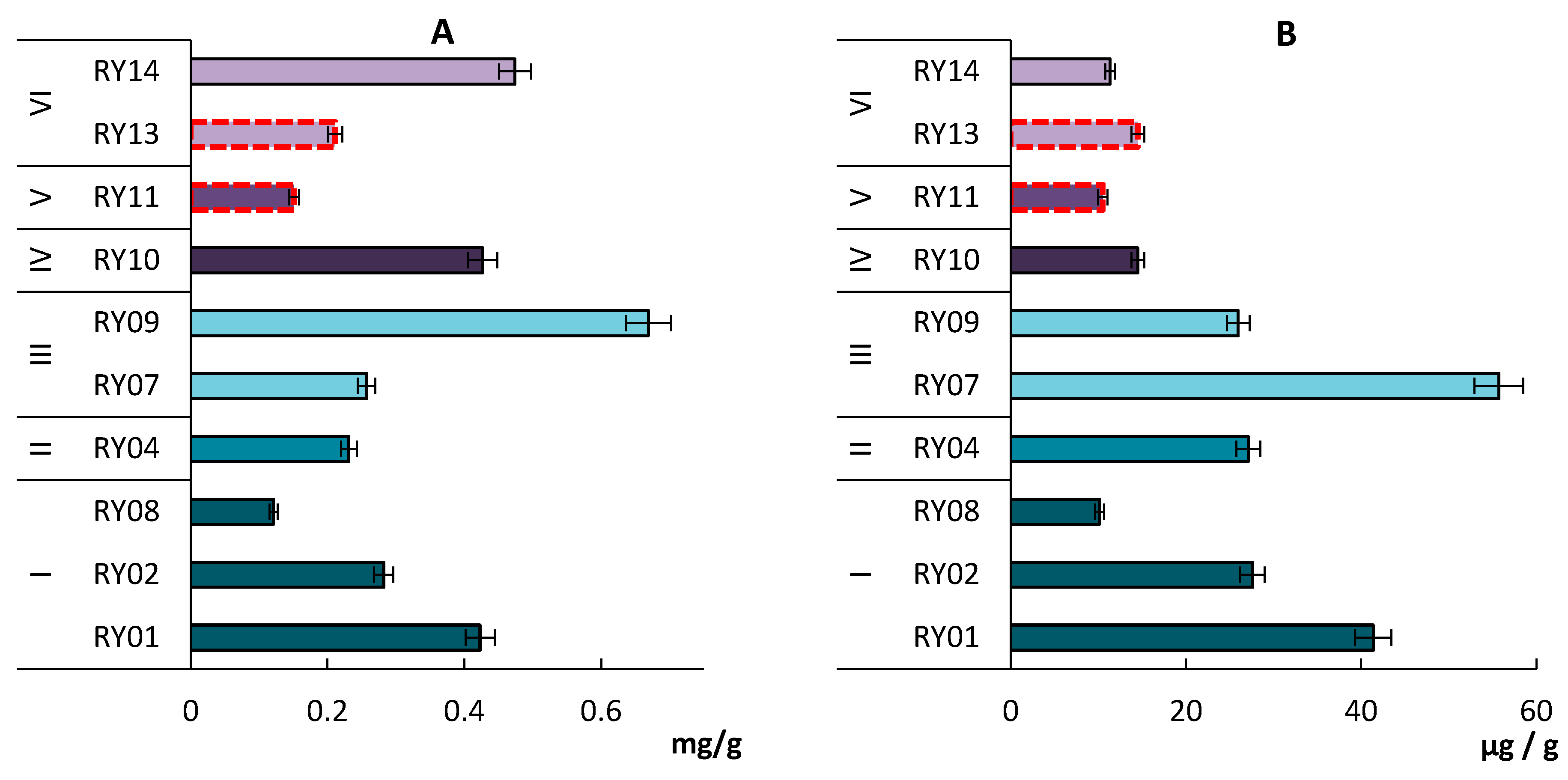
3.2. Fungal Biomass
3.3. Prokaryote Biomass
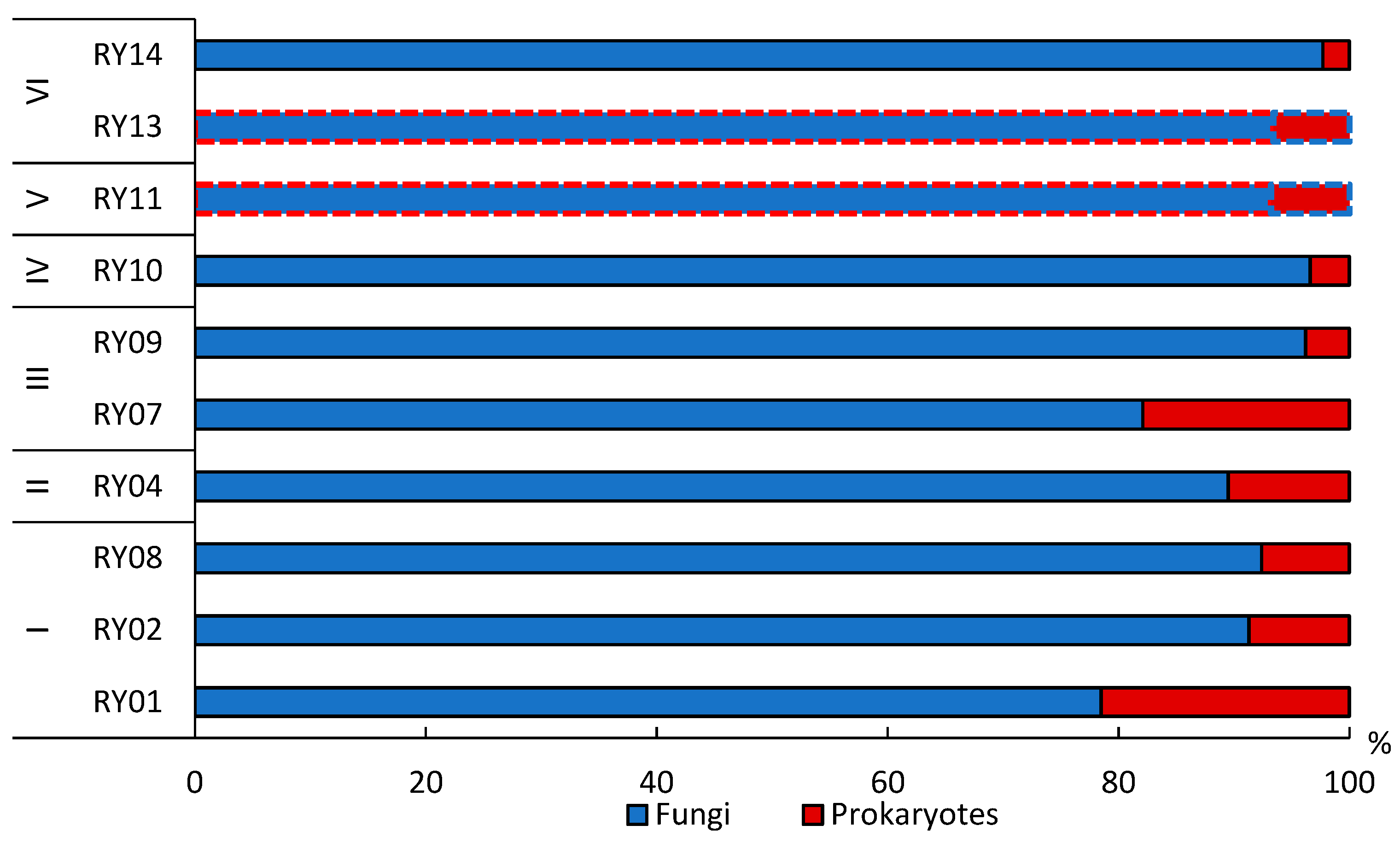
3.4. Total Microbial Biomass
3.5. CFU Number of Microfungi
3.6. Number of Bacteria
3.7. Enzymatic Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flocco, C.G.; Mac Cormack, W.P.; Smalla, K. Antarctic soil microbial communities in a changing environment: Their contributions to the sustainability of Antarctic Ecosystems and the bioremediation of anthropogenic pollution. In The Ecological Role of Microorganisms in the Antarctic Environment; Springer: Berlin/Heidelberg, Germany, 2019; Volume 2, pp. 133–161. [Google Scholar] [CrossRef]
- Dobrovol’skaya, T.G.; Zvyagintsev, D.G.; Chernov, I.Y.; Golovchenko, A.V.; Zenova, G.M.; Lysak, L.V.; Manucharova, N.A.; Marfenina, O.E.; Polyanskaya, L.M.; Stepanov, A.L.; et al. The role of microorganisms in the ecological functions of soils. Eurasian Soil Sci. 2015, 48, 959–967. [Google Scholar] [CrossRef]
- Malard, L.A.; Pearce, D.A. Microbial diversity and biogeography in Arctic soils. Environ. Microbiol. Rep. 2018, 10, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Sebastian, L.; Nicole, S.; Burkhard, B. Biodiversity of biological soil crusts from the Polar Regions revealed by metabarcoding. FEMS Microbiol. Ecol. 2018, 94, fiy036. [Google Scholar]
- Shivaji, S.; Chattopadhyay, M.K.; Reddy, G.S. Diversity of Bacteria from Antarctica, Arctic, Himalayan Glaciers and Stratosphere. Proc. Indian Natl. Sci. Acad. 2019, 85, 909–923. [Google Scholar] [CrossRef]
- Oh, Y.; Zhuang, Q.; Liu, L.; Welp, L.R.; Lau, M.C.; Onstott, T.C.; D’Imperio, L. Reduced net methane emissions due to microbial methane oxidation in a warmer Arctic. Nat. Clim. Chang. 2020, 10, 317–321. [Google Scholar] [CrossRef]
- Tate, K.R. Microbial Biomass: A Paradigm Shift in Terrestrial Biogeochemistry; World Scientific Publ Co. Pte Ltd.: Singapore, 2017; pp. 1–5. [Google Scholar]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Cox, F.; Newsham, K.K.; Bol, R.; Dungait, J.A.; Robinson, C.H. Not poles apart: Antarctic soil fungal communities show similarities to those of the distant Arctic. Ecol. Lett. 2016, 19, 528–536. [Google Scholar] [CrossRef]
- Dobrovolskaya, T.G.; Golovchenko, A.V.; Pankratov, T.A.; Lysak, L.V.; Zvyagintsev, D.G. Assessment of bacterial diversity of soils: Evolution of approaches and methods. Pochvovedenie 2009, 10, 1222–1232. [Google Scholar] [CrossRef]
- Fanin, N.; Kardol, P.M.; Nilsson, M.C.; Gundale, M.J.; Wardle, D.A. The ratio of Gram-positive to Gram-negative bacterial PLFA markers as an indicator of carbon availability in organic soils. Soil Biol. Biochem. 2019, 128, 111–114. [Google Scholar] [CrossRef]
- Demyanyuk, O.; Symochko, L.; Shatsman, D. Structure and dynamics of soil microbial communities of natural and transformed ecosystems. Environ. Res. Eng. Manag. 2020, 76, 97–105. [Google Scholar] [CrossRef]
- Astaykina, A.A.; Streletskii, R.A.; Maslov, M.N.; Belov, A.A.; Gorbatov, V.S.; Stepanov, A.L. The impact of pesticides on the microbial community of agrosoddy-podzolic soil. Eurasian Soil Sci. 2020, 53, 696–706. [Google Scholar] [CrossRef]
- Korneykova, M.V.; Evdokimova, G.A. Microbiota of the ground air layers in natural and industrial zones of the Kola Arctic. J. Environ. Sci. Health. 2018, 53, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Popova, K.B. The Rybachy and Sredny Peninsulas vegetation dataset. Arctic Veg. Arch. Arct. Veg. Classif. 2019, 62, 909–923. [Google Scholar]
- Korneykova, M.V.; Redkina, V.V.; Shalygina, R.R. Algae, cyanobacteria, and microscopic fungi complexes in the Rybachy Peninsula soils, Russia. Czech Polar Rep. 2017, 7, 181–194. [Google Scholar] [CrossRef]
- Evdokimova, G.A.; Mozgova, N.P.; Myazin, V.A. Physicochemical and microbiological characteristics of tundra soils on the Rybachiy Peninsula. Pochvovedenie 2018, 1, 78–85. [Google Scholar] [CrossRef]
- Prokopenko, V.V.; Zenova, G.M.; Manucharova, N.A. Ecophysiological characteristics of psychrotolerant actinomycetes in tundra and forest landscapes. Eurasian Soil Sci. 2019, 52, 682–689. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214, Correction in: 2020, 7, 1–2. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base or Soil Resources 2014, Update 2015. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; p. 203. [Google Scholar]
- Bakermans, C.; Emili, L.A. Terrestrial systems of the Arctic as a model for growth and survival at low temperatures. In Model Ecosystems in Extreme Environments; Academic Press: Cambridge, MA, USA, 2019; pp. 1–21. [Google Scholar] [CrossRef]
- Zvyagintsev, D.G. Methods of Soil Microbiology and Biochemistry; MGU: Moscow, Russia, 1991; p. 304. (in Russian) [Google Scholar]
- Polyanskaya, L.M.; Zvyagintsev, D.G. The content and composition of microbial biomass as an index of the ecological status of soil. Eurasian Soil Sci. 2005, 38, 625–633. [Google Scholar]
- Hoffman, G.; Pallauf, J. Einekolorimetrische method zur Bestimmung der Saccharase-Aktivität von Boden. Z. Pfl. Düng. Bodenk. 1965, 110, 93–101. [Google Scholar]
- Mineev, V.G. Determination of Invertase Activity. In Workshop on Agrochemistry: Textbook Allowance, 2nd ed.; Publishing House of Moscow State University: Sankt Petersburg, Russia, 2001; pp. 346–348. [Google Scholar]
- Shcherbakova, T.A. Enzymatic Activity of Soils and the Transformation of Organic Matter (in Natural and Artificial Phytocenoses); Science and Technology: Minsk, Belarus, 1983; p. 222. [Google Scholar]
- Mineev, V.G. Determination of Dehydrogenase Activity. Workshop on Agrochemistry: Textbook Allowance, 2nd ed.; Publishing House of Moscow State University: Sankt Petersburg, Russia, 2001; pp. 330–332. [Google Scholar]
- Galstyan, A.S. Enzymatic Activity of Soils in Armenia; Yerevan, Hayastan Publ.: Armenia, Yerevan, 1974; p. 274. [Google Scholar]
- Mineev, V.G. Determination of Organic Carbon in the Soil by the Nikitin Method with A Colorimetric Ending According to Orlov-Grindel. Workshop on Agrochemistry: Textbook Allowance, 2nd ed.; Publishing House of Moscow State University: Sankt Petersburg, Russia, 2001; pp. 220–221. [Google Scholar]
- Mastitsky, S.E.; Shitikov, V.K. Statistical Analysis and Data Visualization with R. 2014. E-book. Available online: http://r-analytics.blogspot.com (accessed on 13 May 2023).
- Korneykova, M.V.; Nikitin, D.A.; Myazin, V.A. Qualitative and quantitative characteristics of soil microbiome of Barents Sea coast, Kola Peninsula. Microorganisms 2021, 9, 2126. [Google Scholar] [CrossRef]
- Korneykova, M.V. Comparative analysis of the number and structure of the complexes of microscopic fungi in tundra and taiga soils in the north of the Kola Peninsula. Eurasian Soil Sci. 2018, 51, 89–95. [Google Scholar] [CrossRef]
- Evdokimova, G.A.; Mozgova, N.P. Comparative characterization of microbial biomass in Al–Fe-humus podzols of the Kola Peninsula. Eurasian Soil Sci. 2001, 34, 1305–1312. [Google Scholar]
- Korneikova, M.V.; Nikitin, D.A.; Dolgikh, A.V.; Soshina, A.S. Soil mycobiota of the city of Apatity (Murmansk region). Mikol. Fitopatol. 2020, 54, 264–277. [Google Scholar] [CrossRef]
- Bogorodskaya, A.V. Microbiological assessment of the state of anthropogenically disturbed forest ecosystems of central Siberia. Sib. Forest J. 2016, 2, 71–84. [Google Scholar]
- Fomicheva, O.A.; Polyanskaya, L.M.; Nikonov, V.V.; Lukina, N.V.; Orlova, M.A.; Isaeva, L.G.; Zvyagintsev, D.G. Population and biomass of soil microorganisms in old-growth primary spruce forests in the Northern Taiga. Eurasian Soil Sci. 2006, 39, 1323–1331. [Google Scholar] [CrossRef]
- Schmidt, N.; Bölter, M. Fungal and bacterial biomass in tundra soils along an arctic transect from Taimyr Peninsula, central Siberia. Polar Biol. 2002, 25, 871–877. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Lysak, L.V.; Mergelov, N.S.; Dolgikh, A.V.; Zazovskaya, E.P.; Goryachkin, S.V. Microbial biomass, carbon stocks, and CO2 emission in soils of franz Josef Land: High-Arctic tundra or Polar Deserts. Eurasian Soil Sci. 2020, 53, 467–484. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Semenov, M.V.; Semikolennykh, A.A.; Kachalkin, A.V.; Ivanova, A.E. Biomass of fungi and species diversity of the cultivated mycobiota of soils and substrates in Northbrook Island (Franz Josef Land). Mikol. Fitopatol. 2019, 53, 210–222. [Google Scholar] [CrossRef]
- Polyanskaya, L.M.; Sukhanova, N.I.; Chakmazyan, K.V.; Zvyagintsev, D.G. Changes in the structure of soil microbial biomass under fallow. Eurasian Soil Sci. 2012, 45, 710–716. [Google Scholar] [CrossRef]
- Khabibullina, F.M.; Kuznetsova, E.G.; Vaseneva, I.Z. Micromycetes in podzolic and bog-podzolic soils in the middle taiga subzone of northeastern European Russia. Eurasian Soil Sci. 2014, 47, 1027–1032. [Google Scholar] [CrossRef]
- Dubrova, M.S.; Lubsanova, D.A.; Makarova, E.P.; Kozhevin, P.A.; Manucharova, N.A.; Zenova, G.M. Psychrotolerant actinomycetes in soils of the tundra and northern taiga. Mosc. Univ. Soil Sci. Bull. 2011, 66, 45–49. [Google Scholar] [CrossRef]
- Millán-Aguiñaga, N.; Soldatou, S.; Brozio, S.; Munnoch, J.T.; Howe, J.; Hoskisson, P.A.; Duncan, K.R. Awakening ancient polar Actinobacteria: Diversity, evolution and specialized metabolite potential. Microbiology 2019, 165, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Kudinova, A.G.; Lysak, L.V.; Soina, V.S.; Mergelov, N.S.; Dolgikh, A.V.; Shorkunov, I.G. Bacterial communities in the soils of cryptogamic barrens of East Antarctica (the Larsemann Hills and Thala Hills oases). Eurasian Soil Sci. 2015, 48, 276–287. [Google Scholar] [CrossRef]
- Lysak, L.V.; Lapygina, E.V. The diversity of bacterial communities in urban soils. Eurasian Soil Sci. 2018, 51, 1050–1056. [Google Scholar] [CrossRef]
- Artamonova, V.S. Microbiological features of anthropogenically transformed soils of Western Siberia. V.S.-vo SB RAS. 2002, 225. [Google Scholar]
- Gerasimova, M.; Stroganova, M.; Mozharova, N.; Prokofieva, T. Anthropogenic Soils; Oekumena: Moscow, Russia, 2003; p. 268. [Google Scholar]
- Oleinik, G.N. Bacterioplankton and bacteriobenthos in ecotone ecosystems. Gidrobiol. Mag. 1997, 33, 51–62. [Google Scholar] [CrossRef]
- Akulova, A.Y. The current state of heterotrophic bacteriocenoses of the Kosinsky Trekhozerye. Dis. Cand. Biol. Sci. Mosc. 2017, 260. [Google Scholar]
- Kukharenko, O.S.; Pavlova, N.S.; Dobrovolskaya, T.G.; Golovchenko, A.B.; Pochatkova, T.N.; Zenova, G.M.; Zvyagintsev, D.G. Effect of aeration and temperature on the structure of bacterial complexes of high-moor peat bogs. Soil Sci. 2010, 5, 614–620. [Google Scholar]
- Gaponyuk, E.I.; Malakhov, S.V. Complex system of indicators of environmental monitoring of soils. Migration of pollutants in soils and adjacent environments. Gidrometeoizdat 1985, 5, 3–10. [Google Scholar]
- Ferraz de Almeida, R.; Rezende Naves, E.; Pinheiro da Mota, R. Soil quality: Enzymatic activity of soil β-glucosidase. Glob. J. Agric. Res. Rev. 2015, 3, 146–150. [Google Scholar]
- Kovylina, O.P.; Kovylin, N.V. Activity of soil enzymes of gray forest soils in clearings. Actual Probl. For. Complex 2006, 15, 85–87. [Google Scholar]
- Turusov, V.I.; Cheverdin, Y.I.; Titova, T.V.; Bespalov, V.A. Influence of physical properties of chernozem soils on the activity of soil enzymes. Int. J. Humanit. Nat. Sci. 2019, 4, 15–18. [Google Scholar]
- Romanov, V.N.; Zaushintsena, A.V.; Kozhevnikov, N.V. The use of enzyme activity indicators for operational diagnostics of the ecological state of agrogenic soils. Achiev. Sci. Technol. Agro-Ind. Complex 2019, 33, 44–47. [Google Scholar] [CrossRef]
- Uligova, T.S.; Khezheva, F.V.; Takh, I.P. Enzymatic activity of leached chernozems in the Northwest and Central Caucasus. Bull. Maikop State Technol. Univ. 2010, 3, 31–37. [Google Scholar]
- Orlova, I.V.; Shvakova, E.V. Urease activity of soils in the Arctic. In The Collection: Papanin Readings. In Proceedings of the Articles of the Participants of the International Youth Scientific Conference, Moscow, Russia, 18–20 April 2017; pp. 123–126. [Google Scholar]
- Titova, K.V.; Shvakova, E.V.; Popov, S.S. Enzymatic activity of soils of coastal and island territories of the White and Barents Seas. In Arctic Research: From Extensive Development to Integrated Development. In Proceedings of the International Scientific and Practical Conference, Arkhangelsk, Russia, 23–28 April 2018; pp. 199–204.
- Khaziev, F.K. Ecological relations of the enzymatic activity of soils. Ecobiotech 2018, 1, 80–92. [Google Scholar] [CrossRef]
- Tovstik, E.V.; Olkova, A.S. Assessment of the influence of factors of abiotic nature on the enzymatic activity of the soil. Ecobiotech 2021, 4, 128–134. [Google Scholar]
- Walls-Thumma, D. Dehydrogenase Activity in Soil Bacteria. 2000. Available online: https://www.gardenguides.com/138996-types-soil-bacteria.html (accessed on 13 May 2023).
- Utobo, E.B.; Tewari, L. Soil enzymes as bioindicators of soil ecosystem status. Appl. Ecol. Environ. Res. 2014, 13, 147–169. [Google Scholar] [CrossRef]
- Ivashchenko, K.V.; Sushko, S.V.; Selezneva, A.E.; Ananyeva, N.D.; Zhuravleva, A.I.; Kudeyarov, V.N.; Makarov, M.I.; Blagodatsky, S.A. Soil microbial activity along an altitudinal gradient: Vegetation as a main driver beyond topographic and edaphic factors. Appl. Soil Ecol. 2021, 168, 104197. [Google Scholar] [CrossRef]
- Kussainova, M.; Durmuş, M.; Erkoçak, A.; Kızılkaya, R. Soil dehydrogenase activity of natural macro aggregates in a toposequence of forest soil. Eurasian J. Soil Sci. 2013, 2, 69–75. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
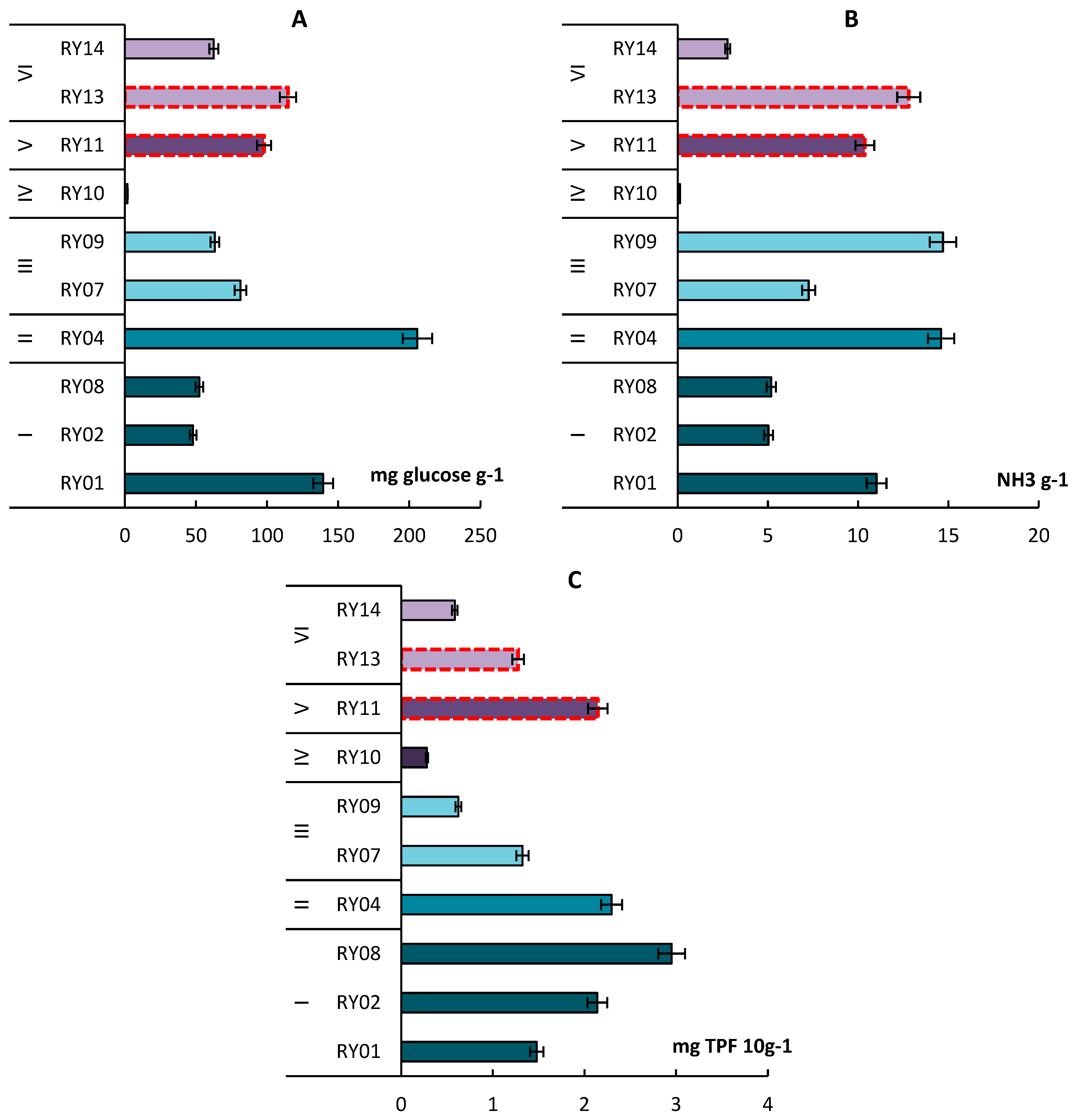{kind=link}
| Soil pit | Coordinates and Height above Sea Level | Type of Soil [20] | Horizons (Depth, cm) | Description of the Site |
|---|---|---|---|---|
| RY01 | 69°49′18.0″ N, 32°02′07.8″ E 35 m | Skeletic Folic Leptic Entic Podzol (Arenic) | O (0–20) Bs (20–40) BC (40–45) | Moss–lichen shrub tundra |
| RY02 | 69°49′18.3″ N 32°02′08.6″ E 34 m | Skeletic Folic Leptic Entic Podzol (Arenic) | O (0–15) Bs (15–45) BC (4–565) | Crooked birch forest shrub–forb |
| RY08 | 69°49′43.8″ N 32°02′37.0″ E 152 m | Skeletic Folic Leptic Entic Podzol (Arenic) | Oa (0–4) Bs (4–30) BC (30–50) | Moss and lichen tundra at the top of the hill |
| RY04 | 69°49′28.0″ N 32°02′12.0″ E 55 m | Dystric Rheic Leptic Rockic Histosol | H (0–35) | Eutrophic swamp |
| RY07 | 69°49′44.3″ N 32°02′31.5″ E 146 m | Skeletic Folic Leptic Albic Podzol (Arenic) | O (0–8) E (8–10) Bs (10–25) BC (25–50) | Shrub tundra, cryogenic crack |
| RY09 | 69°49′43.5″ N 32°02′47.6″ E 150 m | Skeletic Folic Leptic Albic Podzol (Arenic) | O (0–10) Oa (10–20) E (20–35) Bs (35–60) | Shrub tundra, cryogenic hillock |
| RY10 | 69°49′43.6″ N 32°02′42.0″ E 152 m | Skeletic Leptic Entic Podzol (Arenic) | Bs (0–7) BC (7–15) | Cryogenic spot, lichens and cereals |
| RY11 | 69°55′44.7″ N 31°58′08.5″ E 15 m | Dystric Leptic Hemic Folic Histosol (Technic) | Ou (0–10) BC (20–25) | Ruderal tall grass meadow, abandoned village, anthropogenic modified soils (chemical pollution and human impact) |
| RY13 | 69°53′37.4″ N 31°57′44.0″ E 6 m | Dystric Leptic Hemic Folic Histosol | O (0–10) BC (15–25) | Seaside forb meadow, anthropogenic modified soils (attempts at growing crops) |
| RY14 | 69°46′02.5″ N 32°05′56.8″ E 37 m | Dystric Leptic Hemic Folic Histosol | O (0–20) | Crooked birch forest |
| Soil Type | Soil Pit | Horizon | Fungi | Bacteria | |
|---|---|---|---|---|---|
| Copiotrophic | Oligotrophic | ||||
| Skeletic Folic Leptic Entic Podzol | RY01 | O (0–10) | 13.93 ± 3.5 | 814.3 ± 372.9 | 3310.3 ± 843.3 |
| O (10–20) | 6.36 ± 2.5 | 993.4 ± 520.6 | 2254.6 ± 728.3 | ||
| Bs | 1.77 ± 0.2 | 74.0 ± 30 | 342.0 ± 283.0 | ||
| BC | 0.25 ± 0.03 | 8.6 ± 0.6 | 195.7 ± 26.0 | ||
| RY 08 | Oa | 5.9 ± 1.2 | 258.1 ± 113.9 | 672.5 ± 208.5 | |
| Bs | 2.87 ± 0.5 | 385.4 ± 174.7 | 781.5 ± 241.8 | ||
| BC | 0.38 ± 0.03 | 74 ± 11.4 | 199.0 ± 84.8 | ||
| Rheic Leptic Rockic Histosol | RY 04 | H | 0.53 ± 0.05 | 513 ± 146.0 | 736.0 ± 266.9 |
| Skeletic Folic Leptic Albic Podzol | RY 07 | litter | 1.8 ± 0.03 | 513.9 ± 220.0 | 804.4 ± 344.0 |
| O | 0.9 ± −0.005 | 159 ± 3.3 | 219.9 ± 48.4 | ||
| E | 0.31 ± 0.04 | 5.5 ± 11.6 | 36.7 ± 49.1 | ||
| Bs | 0.25 ± 0.02 | 10.2 ± 2.2 | 38.5 ± 17.5 | ||
| BC | 0.04 ± 0.005 | 17.87 ± 5.1 | 17.3 ± 8.9 | ||
| Skeletic Leptic Entic Podzol—cryogenic spot | RY 10 | Bs | 1.12 ± 0.4 | 41.8 ± 11.5 | 77.9 ± 22.7 |
| BC | 0.5 ± 0.02 | 35.4 ± 11.9 | 161.5 ± 78.6 | ||
| Leptic Hemic Folic Histosol (Technic) anthropogenic modified soils (chemical pollution and human impact) | RY 11 | Ou | 0.53 ± 0.10 | 5551.3 ± 627.7 | 12,059.6 ± 1029.5 |
| BC | 0.05 ± 0.005 | 338.3 ± 95.3 | 376.8 ± 163.4 | ||
| Leptic Hemic Folic Histosol anthropogenic modified soils (attempts at growing crops) | RY 13 | O | 0.53 ± 0.09 | 210 ± 98.8 | 422.7 ± 53.0 |
| BC | 0.09 ± 0.003 | 46.65 ± 16.7 | 58.3 ± 28.3 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korneykova, M.V.; Myazin, V.A.; Fokina, N.V.; Chaporgina, A.A.; Nikitin, D.A.; Dolgikh, A.V. Structure of Microbial Communities and Biological Activity in Tundra Soils of the Euro-Arctic Region (Rybachy Peninsula, Russia). Microorganisms 2023, 11, 1352. https://doi.org/10.3390/microorganisms11051352
Korneykova MV, Myazin VA, Fokina NV, Chaporgina AA, Nikitin DA, Dolgikh AV. Structure of Microbial Communities and Biological Activity in Tundra Soils of the Euro-Arctic Region (Rybachy Peninsula, Russia). Microorganisms. 2023; 11(5):1352. https://doi.org/10.3390/microorganisms11051352
Chicago/Turabian StyleKorneykova, Maria V., Vladimir A. Myazin, Nadezhda V. Fokina, Alexandra A. Chaporgina, Dmitry A. Nikitin, and Andrey V. Dolgikh. 2023. "Structure of Microbial Communities and Biological Activity in Tundra Soils of the Euro-Arctic Region (Rybachy Peninsula, Russia)" Microorganisms 11, no. 5: 1352. https://doi.org/10.3390/microorganisms11051352