
The Application of Sulfur Influences Microbiome of Soybean Rhizosphere and Nutrient-Mobilizing Bacteria in Andosol
 , , , , , and
, , , , , and 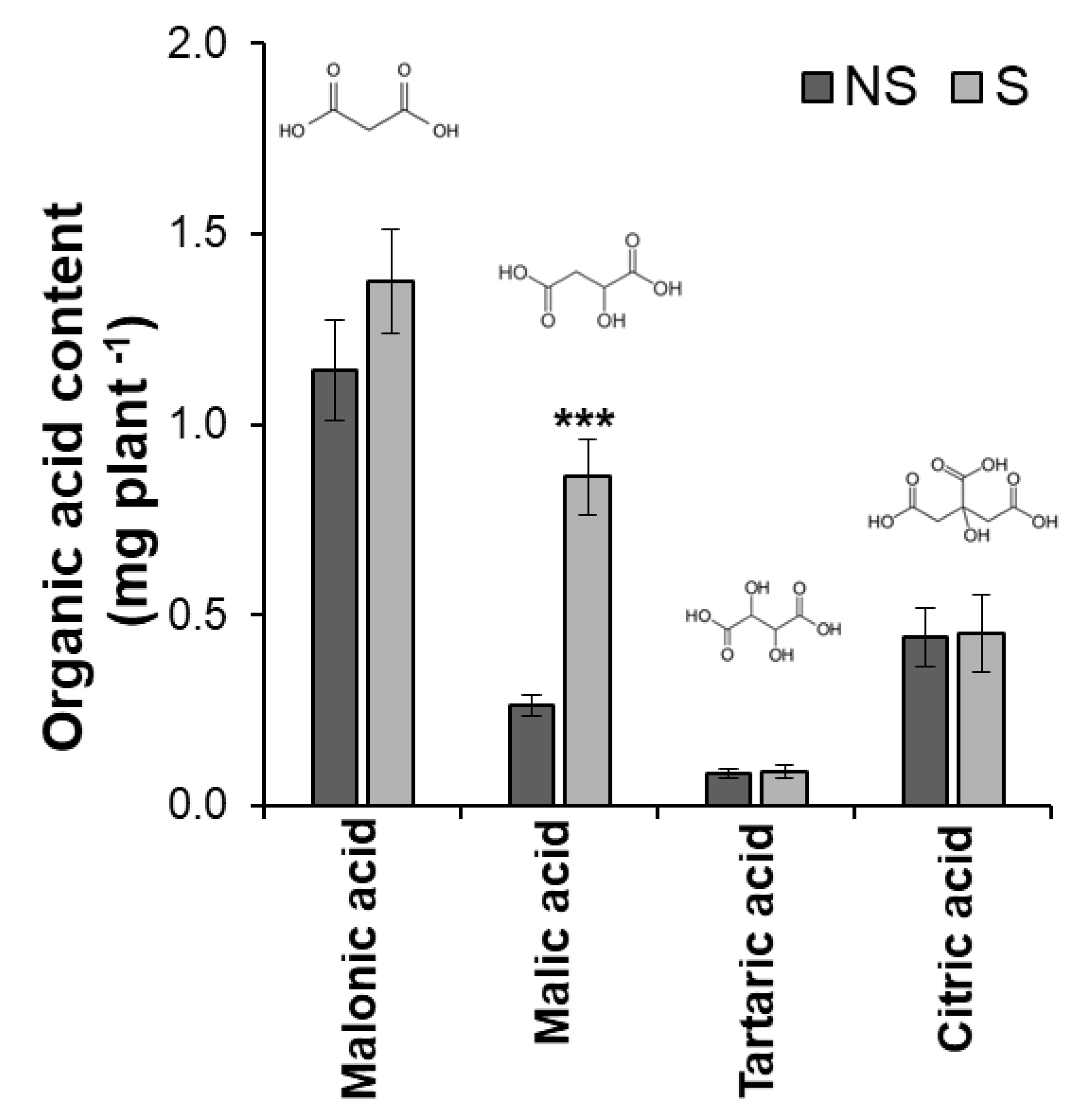{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Analysis
2.2. Soybean Pot Experiment
2.3. Plant P Content and Organic Acid Secretion from Roots
2.4. DNA Extraction and Amplicon Analysis
2.5. Isolation of Microorganism
2.6. PCR Amplification of 16S rRNA Gene Sequence and Phylogenetic Analysis
2.7. Plant Growth-Promoting (PGP) Characteristics
2.7.1. Phosphate-Solubilizing Bacteria (PSB)
2.7.2. Siderophore-Producing Bacteria (SPB)
2.7.3. Arylsulfatase-Producing Bacteria (APB)
2.7.4. Multiple Nutrient Mobilization Trait Screening
2.8. Statistical Analyses
2.9. Nucleotide Sequence Accession Numbers
3. Results
3.1. Influence of S Application on Soybean and Its Microbial Community
3.2. Soil Microbial Diversity When Affected Using Soybean Cultivation and S Application
3.3. Soybean Rhizosphere Microbial Community Composition between S Applications
3.4. Quantification of Cultivable Nutrient-Mobilizing Bacteria in the Rhizospheres of NS- and S-Applied Soybean
3.5. Diversity in Nutrient-Mobilizing Bacteria Isolated from NS- and S-fertilized Soybean
3.6. Evaluation of Nutrient Mobilizing Potential of the Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Skrumsager, I.; White, P. Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; pp. 135–189. [Google Scholar] [CrossRef]
- Dubuis, P.-H.; Marazzi, C.; Stadler, E.; Mauch, F. Sulphur Deficiency Causes a Reduction in Antimicrobial Potential and Leads to Increased Disease Susceptibility of Oilseed Rape. J. Phytopathol. 2005, 153, 27–36. [Google Scholar] [CrossRef]
- Meyer, A.J.; Hell, R. Glutathione homeostasis and redox-regulation by sulfhydryl groups. Photosynth. Res. 2005, 86, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Kertesz, M.A.; Fellows, E.; Schmalenberger, A. Rhizobacteria and Plant Sulfur Supply. Adv. Appl. Microbiol. 2007, 62, 235–268. [Google Scholar] [CrossRef] [PubMed]
- Mukwevho, E.; Ferreira, Z.; Ayeleso, A. Potential Role of Sulfur-Containing Antioxidant Systems in Highly Oxidative Environments. Molecules 2014, 19, 19376–19389. [Google Scholar] [CrossRef]
- Klotz, M.G.; Bryant, D.A.; Hanson, T.E. The Microbial Sulfur Cycle. Front. Microbiol. 2011, 2, 2011. [Google Scholar] [CrossRef]
- Engardt, M.; Simpson, D.; Schwikowski, M.; Granat, L. Deposition of sulphur and nitrogen in Europe 1900–2050. Model calculations and comparison to historical observations. Tellus B Chem. Phys. Meteorol. 2017, 69, 1328945. [Google Scholar] [CrossRef]
- Prasad, R. Major Sulphur Compounds in Plants and their Role in Human Nutrition and Health-An overview. In Proceedings of the Indian National Science Academy, New Delhi, India, 5 December 2014; Volume 80, p. 1045. [Google Scholar] [CrossRef]
- Bouranis, D.; Chorianopoulou, S.; Margetis, M.; Saridis, G.; Sigalas, P. Effect of Elemental Sulfur as Fertilizer Ingredient on the Mobilization of Iron from the Iron Pools of a Calcareous Soil Cultivated with Durum Wheat and the Crop’s Iron and Sulfur Nutrition. Agriculture 2018, 8, 20. [Google Scholar] [CrossRef]
- Astolfi, S.; Celletti, S.; Vigani, G.; Mimmo, T.; Cesco, S. Interaction Between Sulfur and Iron in Plants. Front. Plant Sci. 2021, 12, 670308. [Google Scholar] [CrossRef]
- Sugiura, H.; Sugihara, S.; Kamiya, T.; Ramirez, M.D.A.; Miyatake, M.; Fujiwara, T.; Takuji, O.; Motobayashi, T.; Yokoyama, T.; Bellingrath-Kimura, S.D.; et al. Sulfur application enhances secretion of organic acids by soybean roots and solubilization of phosphorus in rhizosphere. Soil Sci. Plant Nutr. 2021, 67, 400–407. [Google Scholar] [CrossRef]
- Sugiura, H.; Miyaji, S.; Yamamoto, S.; Yasuda, M.; Damo, J.L.C.; Ramirez, M.D.A.; Agake, S.-I.; Kamiya, T.; Fujiwara, T.; Bellingrath-Kimura, S.D.; et al. Induction of citrate transporter gene expression in soybean roots by sulfur application. Soil Sci. Plant Nutr. 2022, 68, 547–552. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; aLey, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. In Proceedings of the National Academy of Sciences, Washington, DC, USA, 1 April 2013; Volume 110, pp. 6548–6553. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhaltnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. In Proceedings of the National Academy of Sciences, Washington, DC, USA, 20 January 2015; Volume 112. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddiquet, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering With Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Kumar, A.; Dubey, A. Rhizosphere microbiome: Engineering bacterial competitiveness for enhancing crop production. J. Adv. Res. 2020, 24, 337–352. [Google Scholar] [CrossRef]
- Arcand, M.M.; Schneider, K.D. Plant-and microbial-based mechanisms to improve the agronomic effectiveness of phosphate rock: A review. An. Acad Bras. Cienc. 2006, 78, 791–807. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319339. [Google Scholar] [CrossRef]
- Whitelaw, M. Growth promotion of plants inoculated with phosphate solubilizing fungi. Adv. Agron. 2000, 69, 99–151. [Google Scholar]
- Singh, S.K. Microbes for iron chlorosis remediation In Peach. In Biostimulants in Plant Science; Intech Open: London, UK, 2020. [Google Scholar]
- Cregut, M.; Piutti, S.; Slezack-Deschaumes, S.; Benizri, E. Compartmentalization and regulation of arylsulfatase activities in Streptomyces sp., Microbacterium sp. and Rhodococcus sp. soil isolates in response to inorganic sulfate limitation. Microbiol. Res. 2013, 168, 12–21. [Google Scholar] [CrossRef]
- Ahkami, A.H.; White, R.A.; Handakumbura, P.P.; Jansson, C. Rhizosphere engineering: Enhancing sustainable plant ecosystem productivity. Rhizosphere 2017, 3, 233–243. [Google Scholar] [CrossRef]
- Sugiyama, A.; Yazaki, K. Root Exudates of Legume Plants and Their Involvement in Interactions with Soil Microbes. In Secretions and Exudates in Biological Systems; Springer: Berlin/Heidelberg, Germany, 2012; pp. 27–48. [Google Scholar] [CrossRef]
- Currier, W.W.; Strobel, G.A. Chemotaxis of Rhizobium spp. to Plant Root Exudates. Plant Physiol. 1976, 57, 820–823. [Google Scholar] [CrossRef]
- Kosslak, R.M.; Bookland, R.; Barkei, J.; Paaren, H.E.; Appelbaum, E.R. Induction of Bradyrhizobium japonicum common nod genes by isoflavones isolated from Glycine max. In Proceedings of the National Academy of Sciences, Washington, DC, USA, 1 November 1987; Volume 84, pp. 7428–7432. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef]
- Van Reeuwijk, L. (Ed.) Procedures for Soil Analysis, 6th ed.; Technical Paper; International Soil Reference and Information Centre: Wageningen, The Netherlands, 2002. [Google Scholar]
- Miyatake, M.; Ohyama, T.; Yokoyama, T.; Sugihara, S.; Motobayashi, T.; Kamiya, T.; Fujiwara, T.; Yuan, K.; Bellingrath-Kimura, S.D.; Ohkalma-Ohtsu, N. Effects of deep placement of controlled-release nitrogen fertilizer on soybean growth and yield under sulfur deficiency. Soil Sci. Plant Nutr. 2019, 65, 259–266. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Fontes, M.P.F.; Weed, S.B. Phosphate adsorption by clays from Brazilian Oxisols: Relationships with specific surface area and mineralogy. Geoderma 1996, 72, 37–51. [Google Scholar] [CrossRef]
- Noulas, C.; Karyotis, T.; Charoulis, A.; Massas, I. Red Mediterranean Soils: Nature, Properties, and Management of Rhodoxeralfs in Northern Greece. Commun. Soil Sci. Plant Anal. 2009, 40, 633–648. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- Imai, K.; Sugihara, S.; Wasaki, J.; Tanaka, H. Effects of white lupin and groundnut on fractionated rhizosphere soil P of different P-limited soil types in Japan. Agronomy 2019, 9, 68. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Gadagi, R.S.; Sa, T. New isolation method for microorganisms solubilizing iron and aluminum phosphates using dyes. Soil Sci. Plant Nutr. 2002, 48, 615–618. [Google Scholar] [CrossRef]
- Arora, N.K.; Verma, M. Modified microplate method for rapid and efficient estimation of siderophore produced by bacteria. Biotech 2017, 7, 381. [Google Scholar] [CrossRef]
- Bouranis, D.L.; Venieraki, A.; Chorianopoulou, S.N.; Katinakis, P. Impact of elemental sulfur on the rhizospheric bacteria of durum wheat crop cultivated on a calcareous soil. Plants 2019, 8, 379. [Google Scholar] [CrossRef]
- Gregersen, T. Rapid method for distinction of gram-negative from gram-positive bacteria. Eur. J. Appl. Microbiol. Biotechnol. 1978, 5, 123–127. [Google Scholar] [CrossRef]
- Habibi, S.; Djedidi, S.; Prongjunthuek, K.; Mortuzal, F.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyoma, T. Physiological and genetic characterization of rice nitrogen fixer PGPR isolated from rhizosphere soils of different crops. Plant Soil 2014, 379, 51–66. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Miwa, H.; Obirih-Opareh, J.; Suzaki, T.; Yasuda, M.; Okazaki, S. Novel rhizobia exhibit superior nodulation and biological nitrogen fixation even under high nitrate concentrations. FEMS Microbiol. Ecol. 2020, 96, fiz184. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Premono Edi, M.; Moawad, A.M.; Vlek, P.L.G. Effect of phosphate solubilizing Pseudomonas putida on the growth of maize and its survival in the rhizosphere. Indones. J. Crop Sci. 1996, 11, 13–23. [Google Scholar]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Zelenev, V.V.; van Bruggen, A.H.C.; Semenov, A.M. Modelling wave-like dynamics of oligotrophic andcopiotrophic bacteria along wheat roots in response to nutrient input from a growing root tip. Ecol. Model. 2005, 188, 404–417. [Google Scholar] [CrossRef]
- Hirsch, P.R.; Miller, A.J.; Dennis, P.G. Do root exudates exert more influence on rhizosphere bacterial community structure than other rhizodeposits. In Molecular Microbial Ecology of the Rhizosphere; Wiley Press: Hoboken, NJ, USA, 2013; pp. 229–242. [Google Scholar]
- Igiehon, N.O.; Babalola, O.O.; Cheseto, X.; Torto, B. Effects of rhizobia and arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress. Microbiol. Res. 2021, 242, 126640. [Google Scholar] [CrossRef]
- Zeng, W.; Bai, X.; Zhang, L.; Wang, A.; Peng, Y. Population dynamics of nitrifying bacteria for nitritation achieved in Johannesburg (JHB) process treating municipal wastewater. Bioresour. Technol. 2014, 162, 30–37. [Google Scholar] [CrossRef]
- Yoneda, Y.; Yamamoto, K.; Makino, A.; Tanaka, Y.; Meng, X.-Y.; Hashimoto, J.; Shin-Ya, K.; Satoh, N.; Fujie, M.; Toyama, T.; et al. Novel Plant-Associated Acidobacteria Promotes Growth of Common Floating Aquatic Plants, Duckweeds. Microorganisms 2021, 9, 1133. [Google Scholar] [CrossRef]
- Hausmann, B.; Pelikan, C.; Herbold, C.W.; Köstlbacher, S.; Albertsen, M.; Eichorst, S.A.; del Rio, T.G.; Huemer, M.; Nielsen, P.H.; Rattei, T.; et al. Peatland Acidobacteria with a dissimilatory sulfur metabolism. ISME J. 2018, 12, 1729–1742. [Google Scholar] [CrossRef]
- Tang, X.; Li, L.; Wu, C.; Khan, M.I.; Manzoor, M.; Zou, L.; Shi, J. The response of arsenic bioavailability and microbial community in paddy soil with the application of sulfur fertilizers. Environ. Pollut. 2020, 264, 114679. [Google Scholar] [CrossRef]
- Sun, L.; Song, K.; Shi, L.; Duan, D.; Zhang, H.; Sun, Y.; Qin, Q. Influence of elemental sulfur on cadmium bioavailability, microbial community in paddy soil and Cd accumulation in rice plants. Sci. Rep. 2021, 11, 11468. [Google Scholar] [CrossRef]
- Mondal, S.; Pramanik, K.; Panda, D.; Dutta, D.; Karmakar, S.; Bose, B. Sulfur in Seeds: An Overview. Plants 2022, 11, 450. [Google Scholar] [CrossRef]
- Takahashi, T.; Mitamura, A.; Ito, T.; Ito, K.; Nanzyo, M.; Saigusa, M. Aluminum solubility of strongly acidified allophanic Andosols from Kagoshima Prefecture, southern Japan. Soil Sci. Plant Nutr. 2008, 54, 362–368. [Google Scholar] [CrossRef]
- Vigani, G.; Pii, Y.; Celletti, S.; Maver, M.; Mimmo, T.; Cesco, S.; Astolfi, S. Mitochondria dysfunctions under Fe and S deficiency: Is citric acid involved in the regulation of adaptive responses? Plant Physiol. Biochem. 2018, 126, 86–96. [Google Scholar] [CrossRef]
- Jones, D.L.; Darrah, P.R. Role of root derived organic acids in the mobilization of nutrients from the rhizosphere. Plant Soil 1994, 166, 247–257. [Google Scholar] [CrossRef]
- Drábek, O.; Kipkoech Kiplagat, I.; Komárek, M.; Tejnecký, V.; Borůvka, L. Study of interactions between relevant organic acids and aluminium in model solutions using HPLC and IC. Soil Water Res. 2015, 10, 172–180. [Google Scholar] [CrossRef]
- Somers, E.; Vanderleyden, J.; Srinivasan, M. Rhizosphere Bacterial Signalling: A Love Parade Beneath Our Feet. Crit. Rev. Microbiol. 2004, 30, 205–240. [Google Scholar] [CrossRef]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef]
- Rekha, K.; Baskar, B.; Srinath, S.; Usha, B. Plant-growth-promoting rhizobacteria Bacillus subtilis RR4 isolated from rice rhizosphere induces malic acid biosynthesis in rice roots. Can. J. Microbiol. 2018, 64, 20–27. [Google Scholar] [CrossRef]
- Gahan, J.; Schmalenberger, A. The role of bacteria and mycorrhiza in plant sulfur supply. Front Plant Sci. 2014, 5, 723. [Google Scholar] [CrossRef]
- Okazaki, K.; Tsurumaru, H.; Hashimoto, M.; Takahashi, H.; Okubo, T.; Ohwada, T.; Minamisawa, K. Community Analysis-based Screening of Plant Growth-promoting Bacteria for Sugar Beet. Microbes Environ. 2021, 36, ME20137. [Google Scholar] [CrossRef]
- Martín-Mora, D.; Ortega, Á.; Pérez-Maldonado, F.J.; Krell, T.; Matilla, M.A. The activity of the C4-dicarboxylic acid chemoreceptor of Pseudomonas aeruginosa is controlled by chemoattractants and antagonists. Sci. Rep. 2018, 8, 2102. [Google Scholar] [CrossRef]
- Oku, S.; Komatsu, A.; Nakashimada, Y.; Tajima, T.; Kato, J. Identification of Pseudomonas fluorescens chemotaxis sensory proteins for malate, succinate, and fumarate, and their involvement in root colonization. Microbes Environ. 2014, 29, 413–419. [Google Scholar] [CrossRef]
- Falagán, C.; Johnson, D.B. Acidibacter ferrireducens gen. nov., sp. nov.: An acidophilic ferric iron-reducing gammaproteobacterium. Extremophiles 2014, 18, 1067–1073. [Google Scholar] [CrossRef]
- Damo, J.L.C.; Ramiretz, M.D.A.; Agake, S.-I.; Pedro, M.; Brown, M.; Sekimoto, H.; Yokoyama, T.; Sugihara, S.; Okazaki, S.; Ohkama-Ohtsu, N. Isolation and Characterization of Phosphate Solubilizing Bacteria from Paddy Field Soils in Japan. Microbes Environ. 2022, 37, ME21085. [Google Scholar] [CrossRef]
- Weisskopf, L.; Heller, S.; Eberl, L. Burkholderia Species Are Major Inhabitants of White Lupin Cluster Roots. Appl. Environ. Microbiol. 2011, 77, 7715–7720. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damo, J.L.C.; Shimizu, T.; Sugiura, H.; Yamamoto, S.; Agake, S.-i.; Anarna, J.; Tanaka, H.; Sugihara, S.; Okazaki, S.; Yokoyama, T.; et al. The Application of Sulfur Influences Microbiome of Soybean Rhizosphere and Nutrient-Mobilizing Bacteria in Andosol. Microorganisms 2023, 11, 1193. https://doi.org/10.3390/microorganisms11051193
Damo JLC, Shimizu T, Sugiura H, Yamamoto S, Agake S-i, Anarna J, Tanaka H, Sugihara S, Okazaki S, Yokoyama T, et al. The Application of Sulfur Influences Microbiome of Soybean Rhizosphere and Nutrient-Mobilizing Bacteria in Andosol. Microorganisms. 2023; 11(5):1193. https://doi.org/10.3390/microorganisms11051193
Chicago/Turabian StyleDamo, Jean Louise Cocson, Takashi Shimizu, Hinako Sugiura, Saki Yamamoto, Shin-ichiro Agake, Julieta Anarna, Haruo Tanaka, Soh Sugihara, Shin Okazaki, Tadashi Yokoyama, and et al. 2023. "The Application of Sulfur Influences Microbiome of Soybean Rhizosphere and Nutrient-Mobilizing Bacteria in Andosol" Microorganisms 11, no. 5: 1193. https://doi.org/10.3390/microorganisms11051193