
The Effects of Different Feeding Regimes on Body Composition, Gut Microbial Population, and Susceptibility to Pathogenic Infection in Largemouth Bass

Abstract
:1. Introduction
2. Materials and Methods
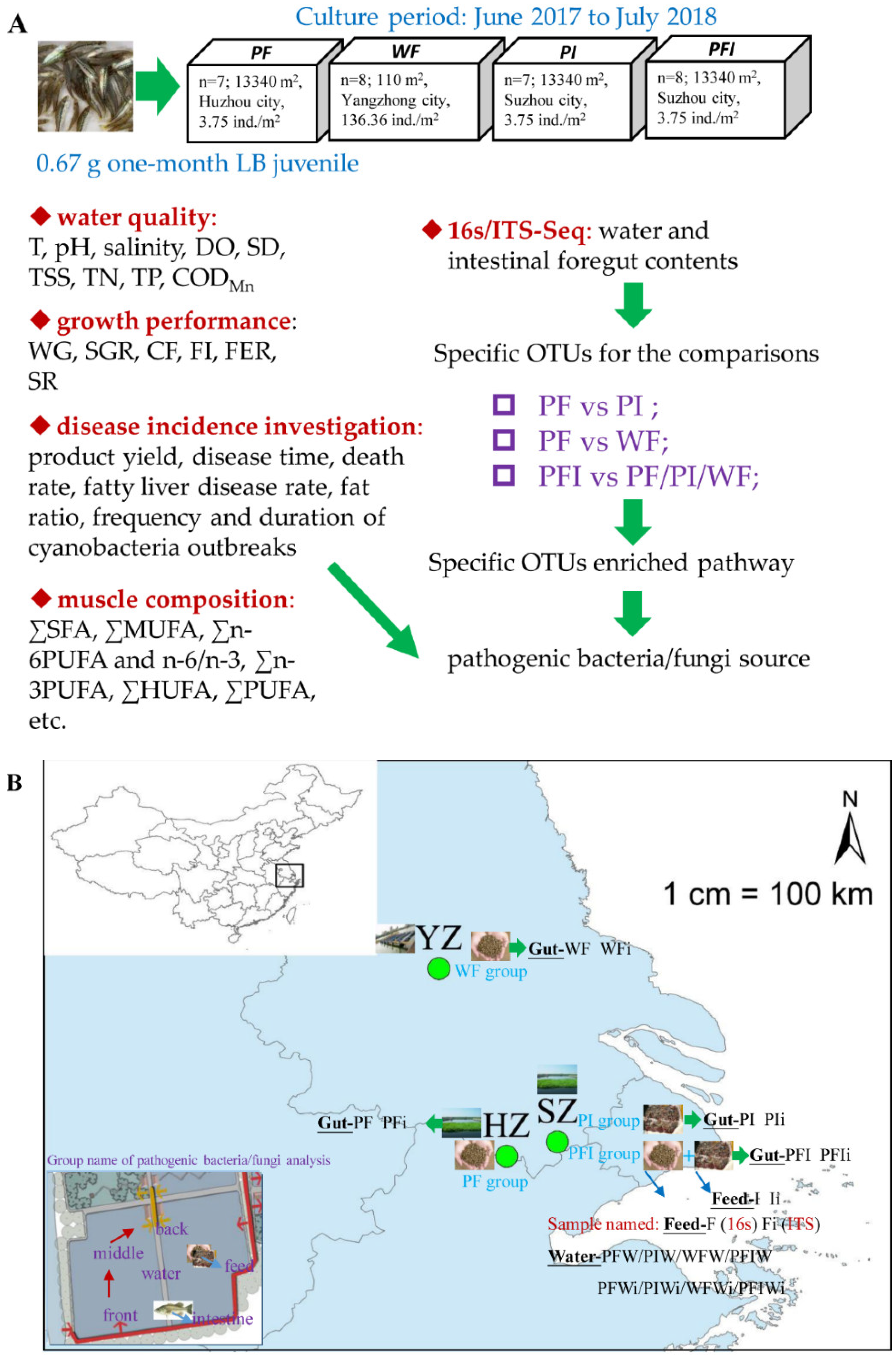
2.1. Experimental Design and Sample Collection
2.2. Water Quality and Epidemiological Survey
2.3. Growth Performance and Body Composition
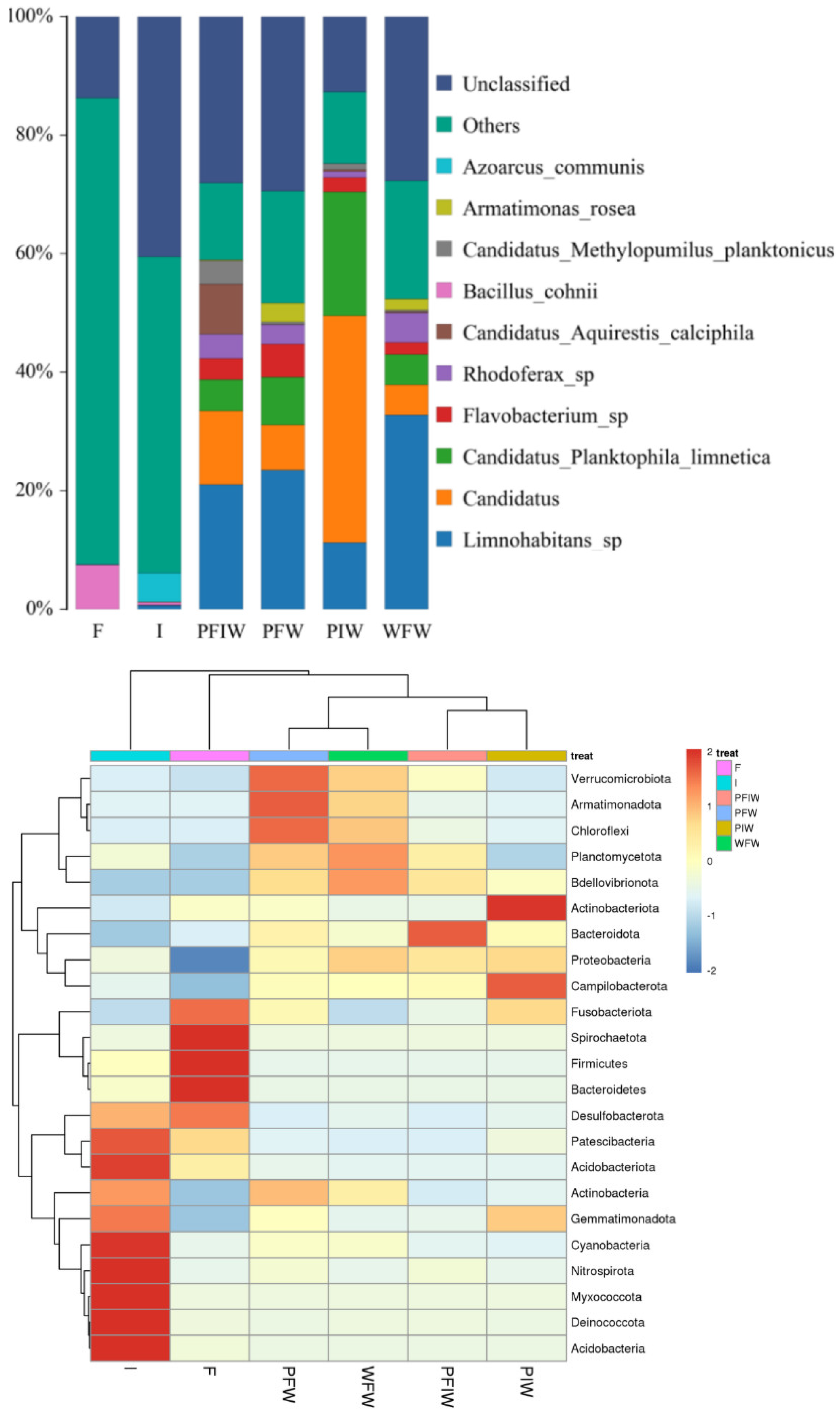
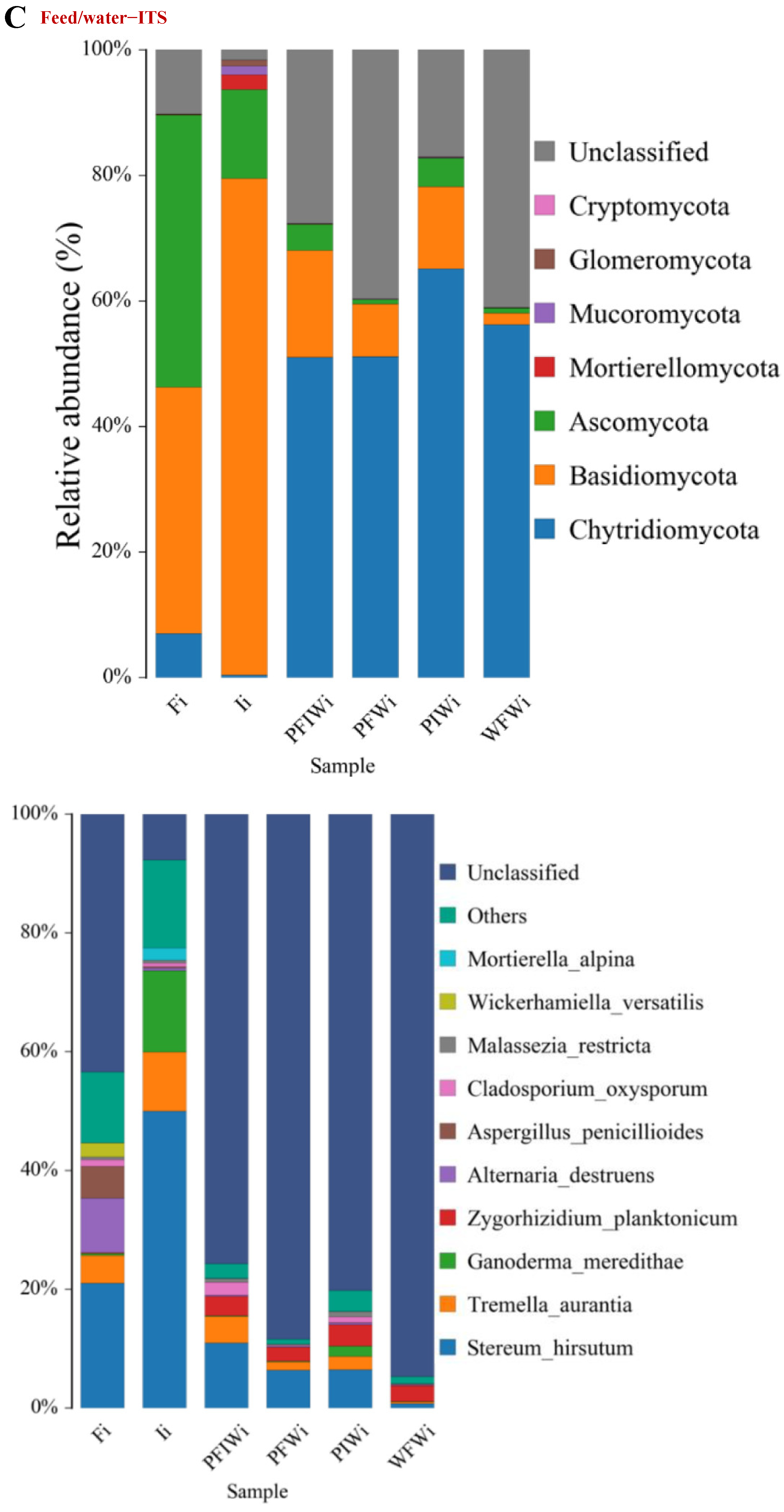
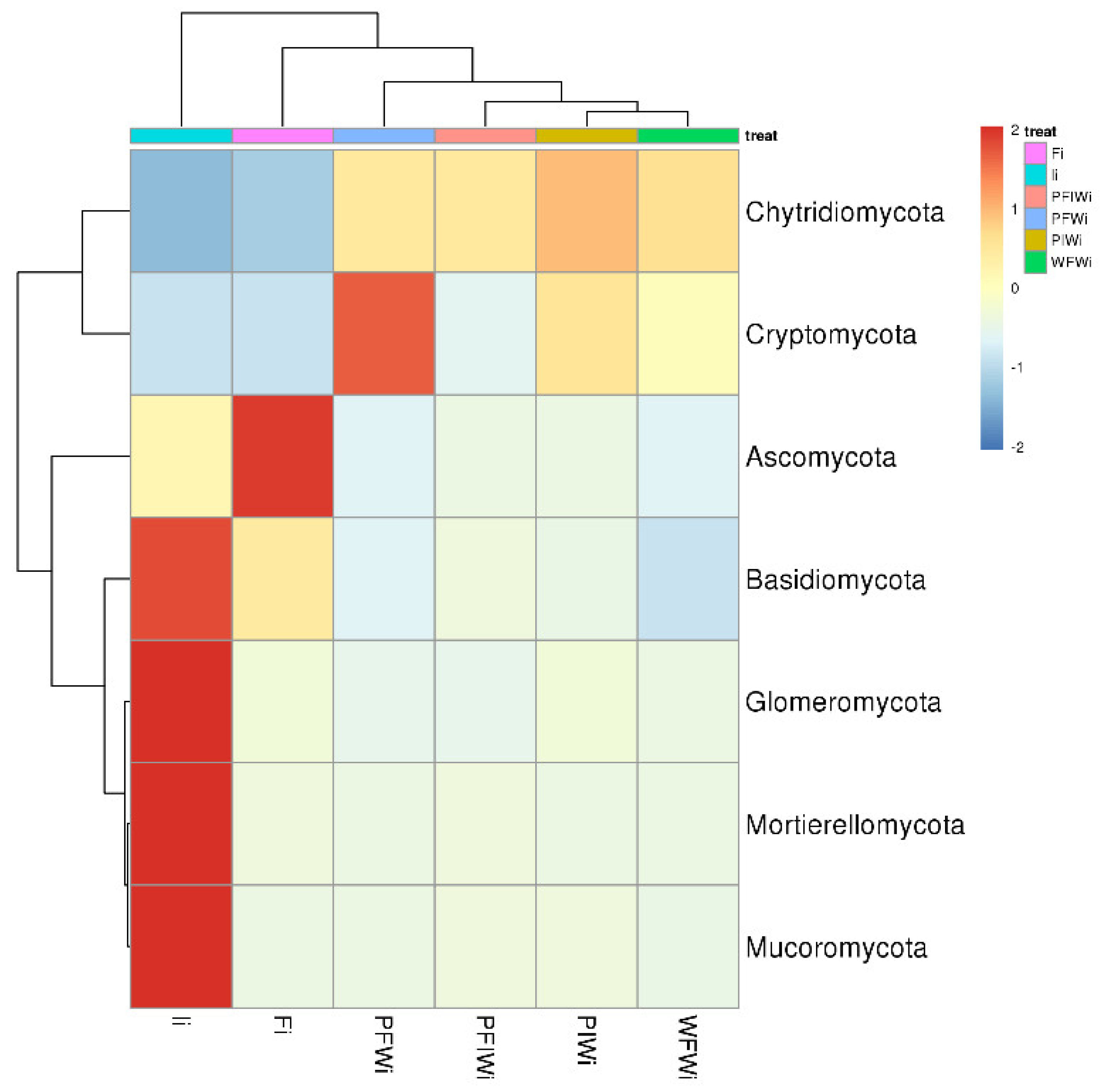
2.4. Sequencing and Taxonomy Classification
2.5. Change in Microbial Communities among Different Feeding Strategies
2.6. Data Analyses
3. Results
3.1. Water Quality and Disease Incidence
3.2. Growth Performance
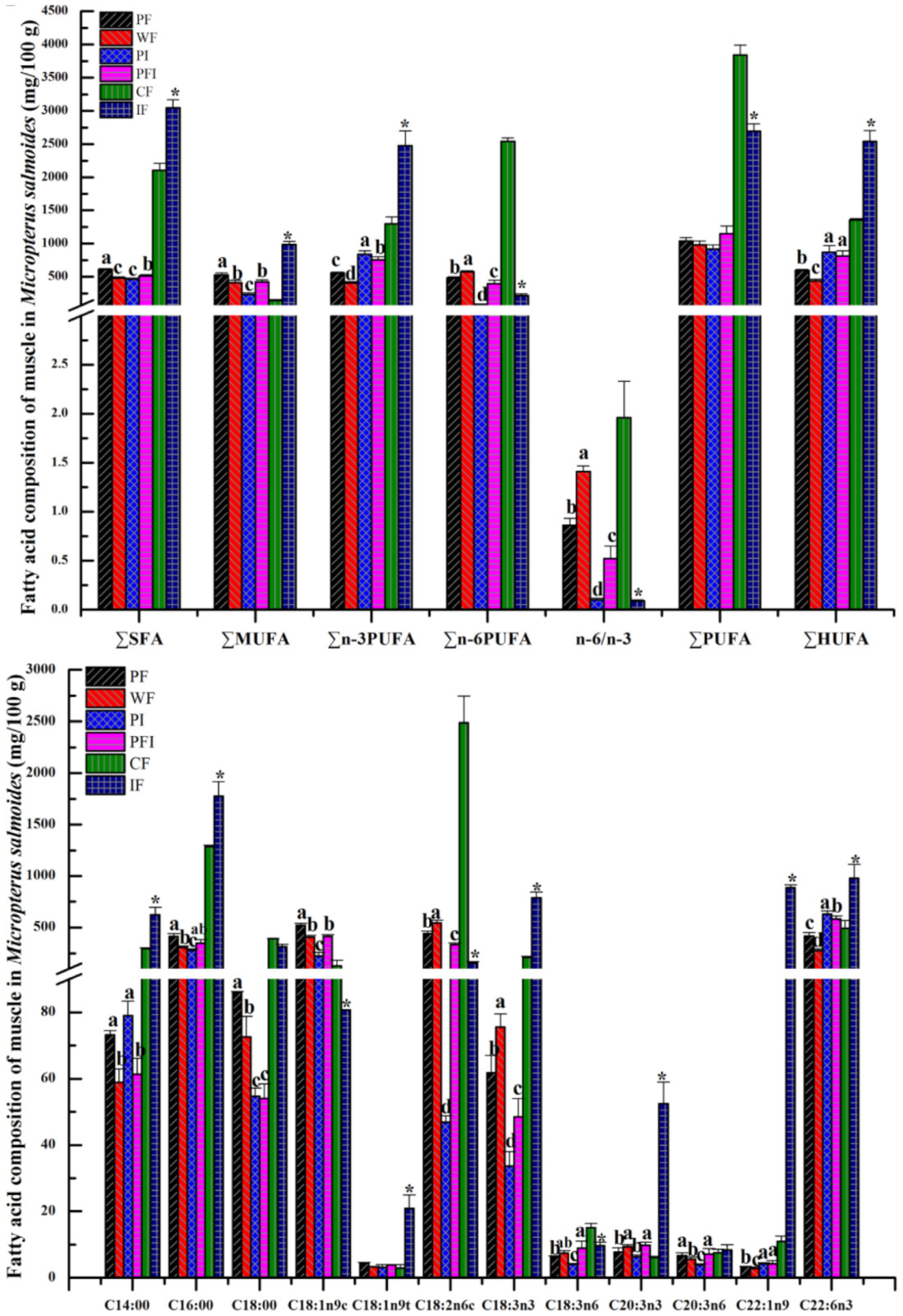
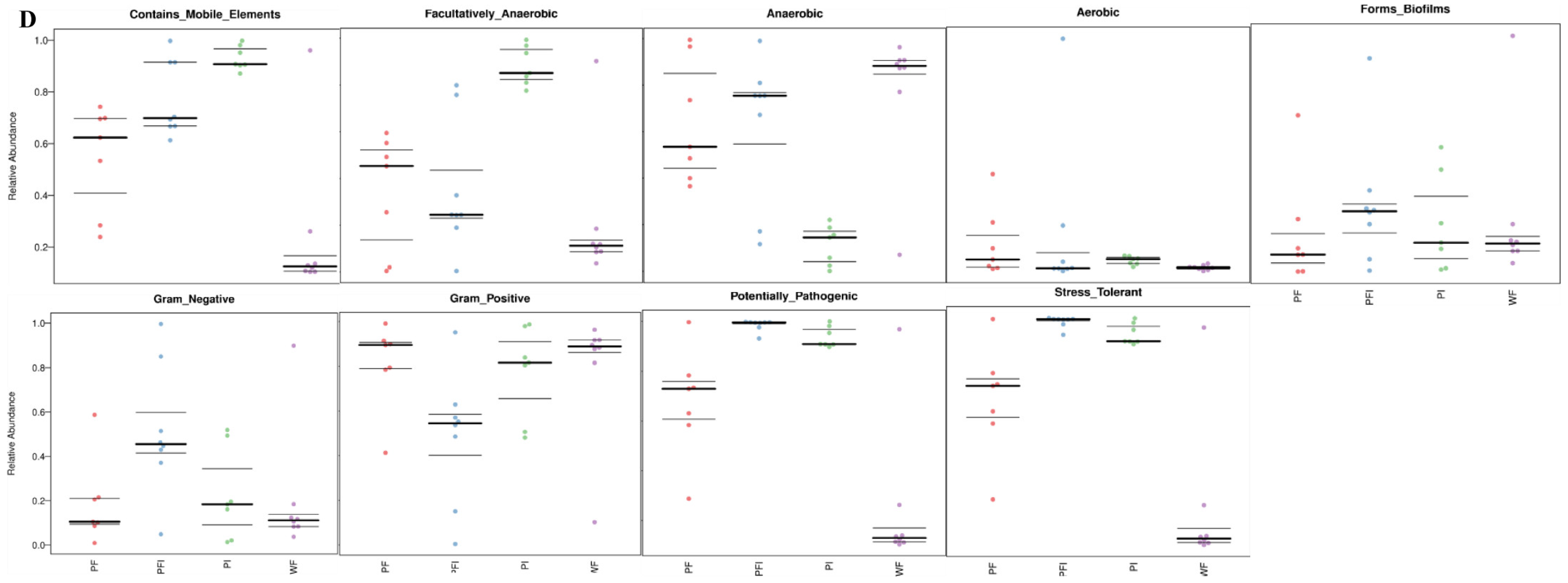
3.3. Body Composition
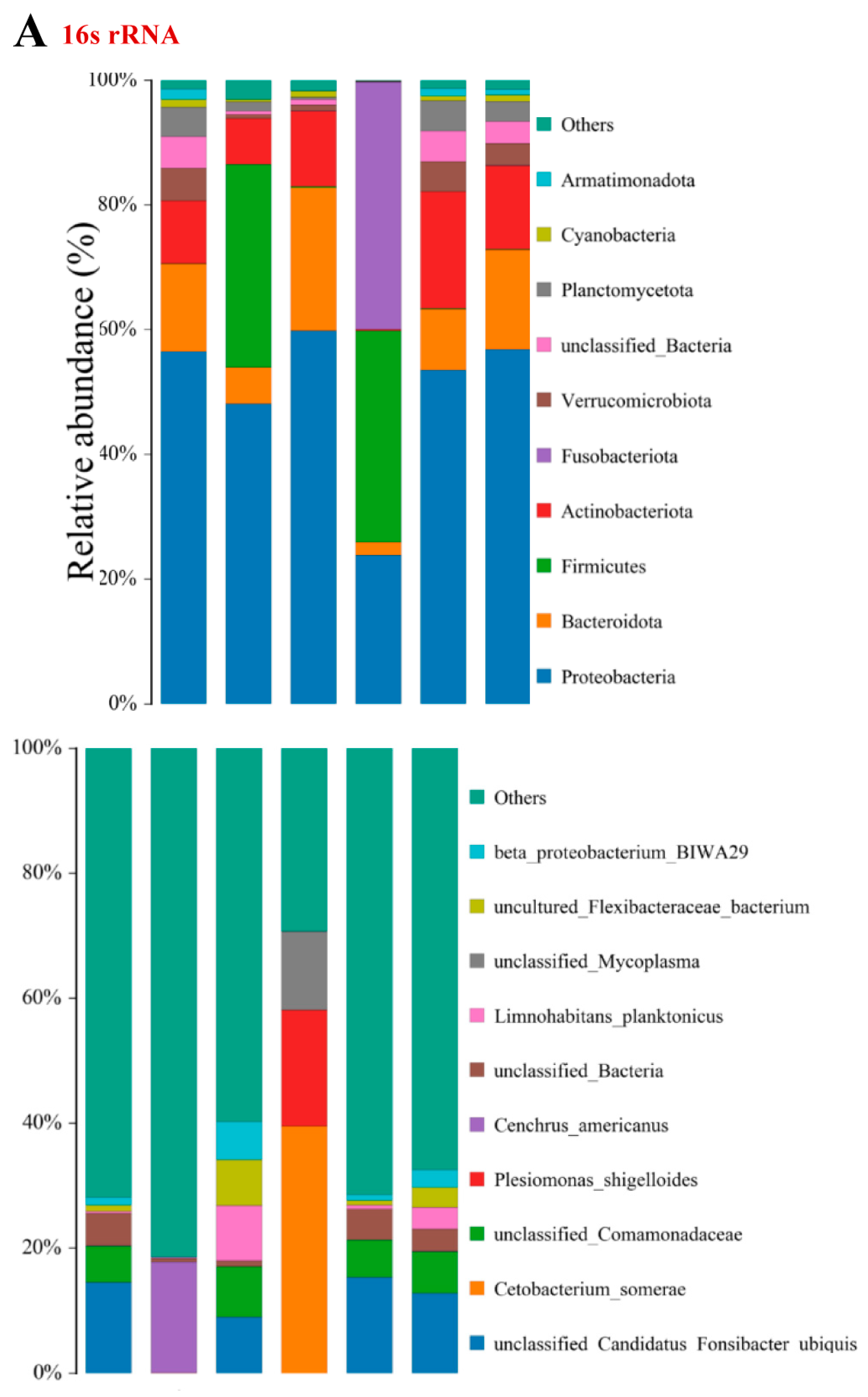
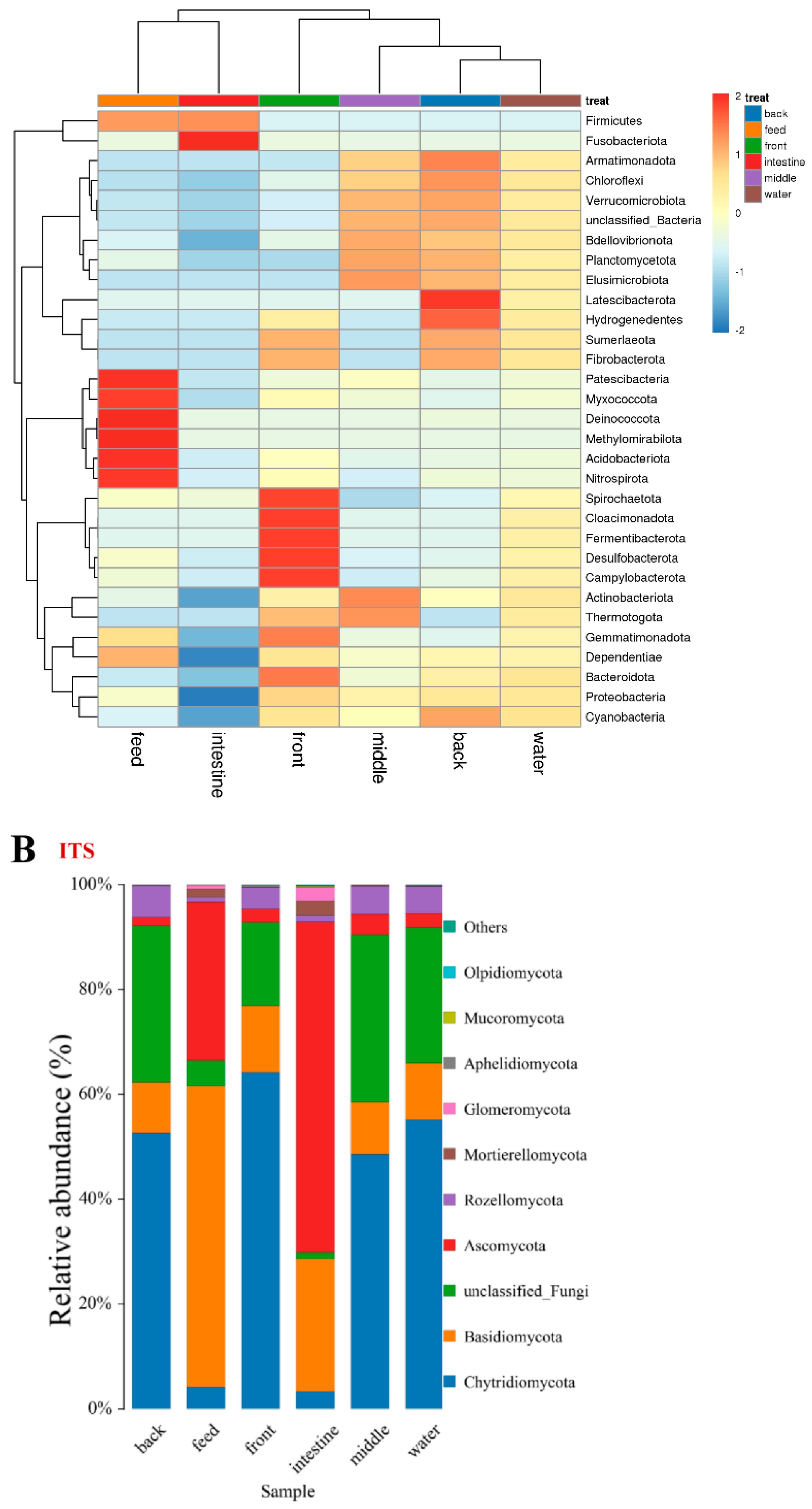
3.4. α- and β-Diversity
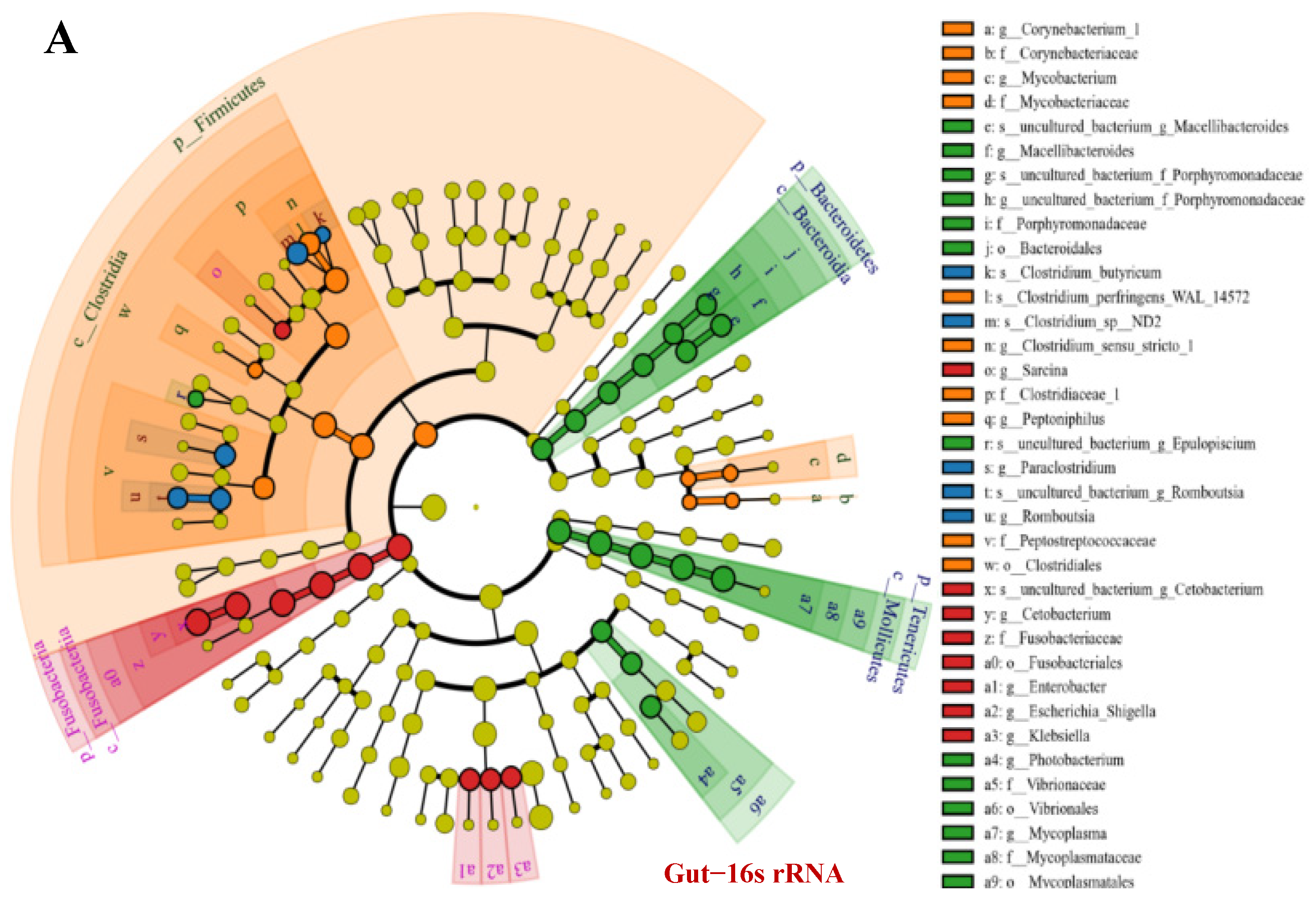
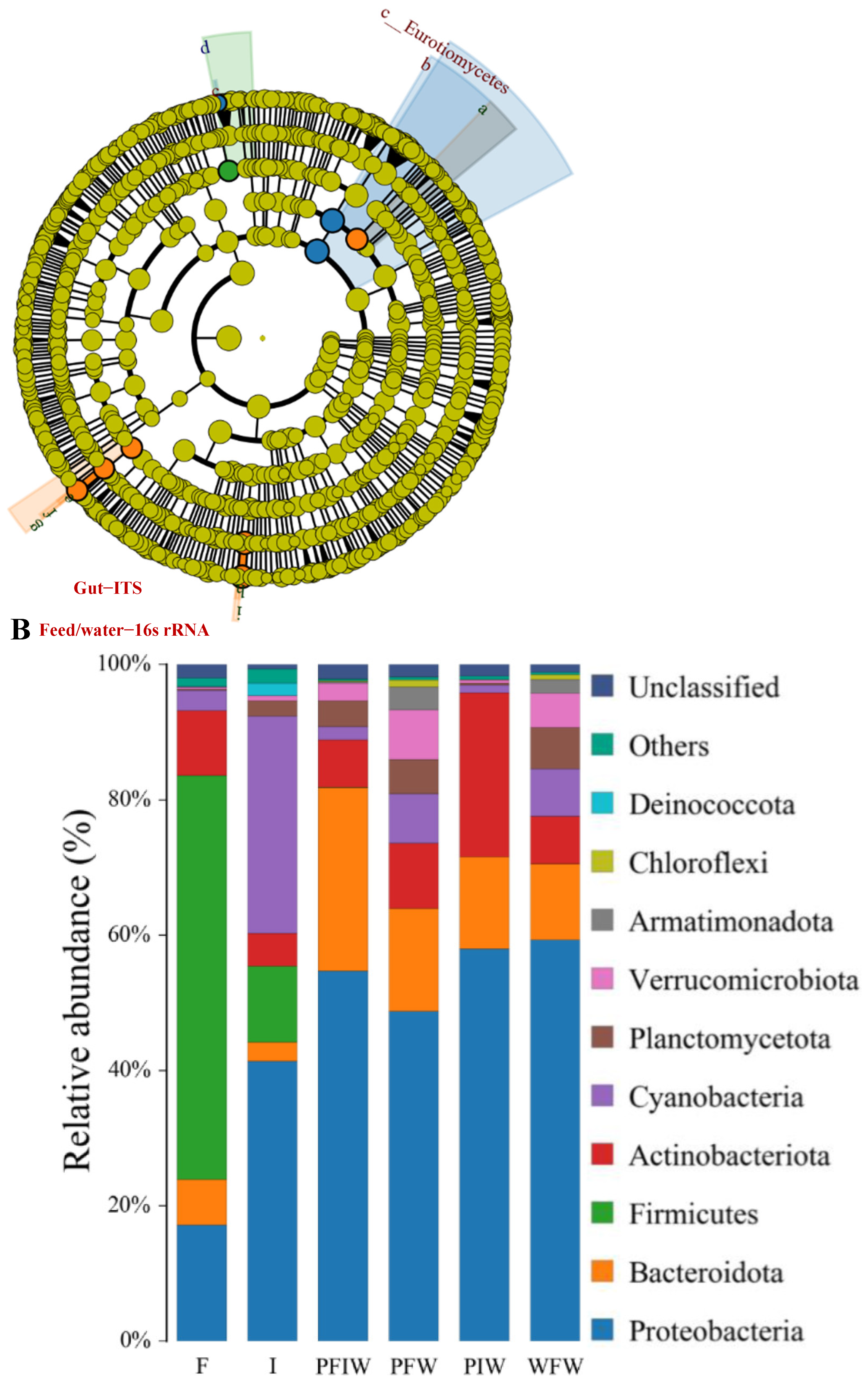
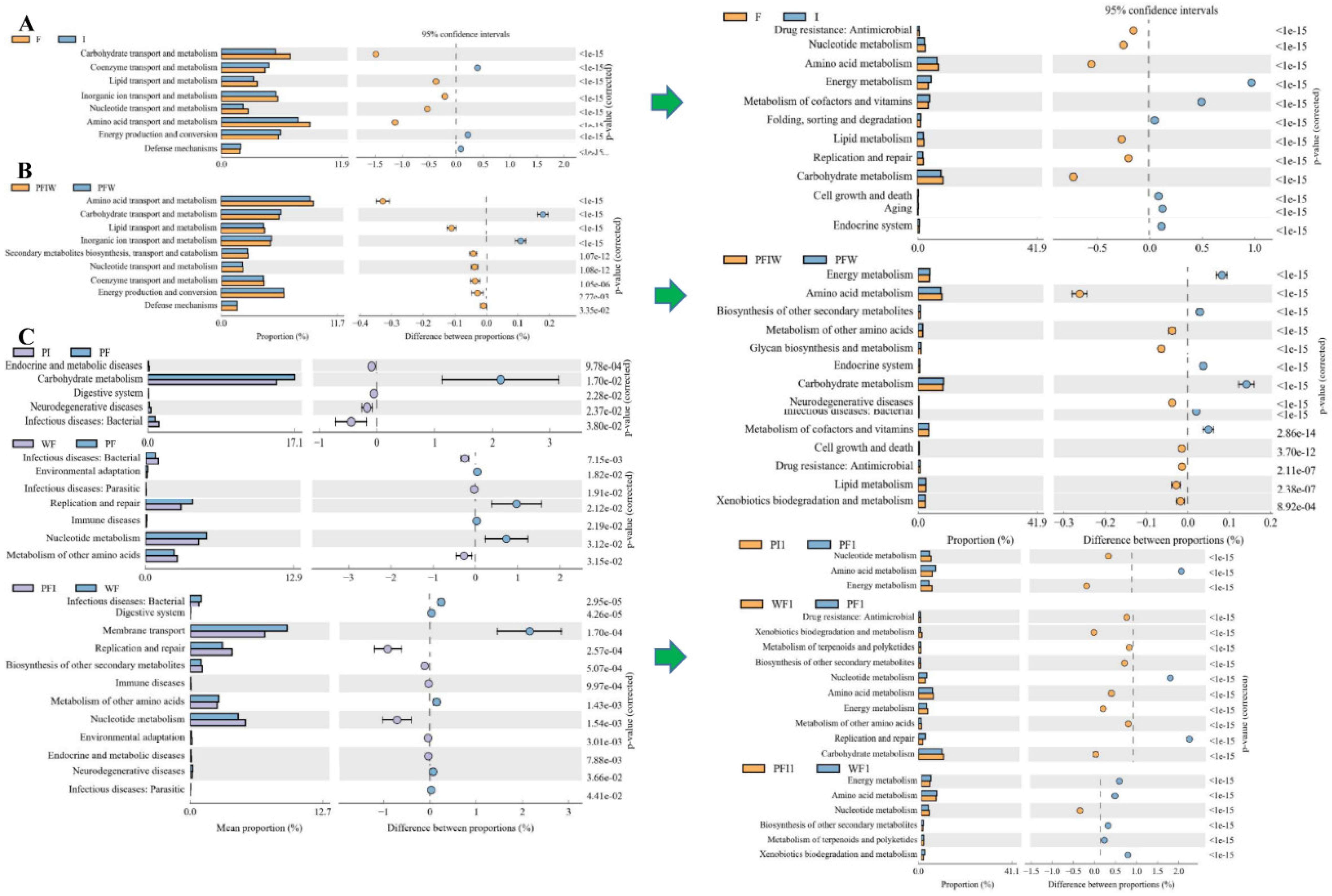
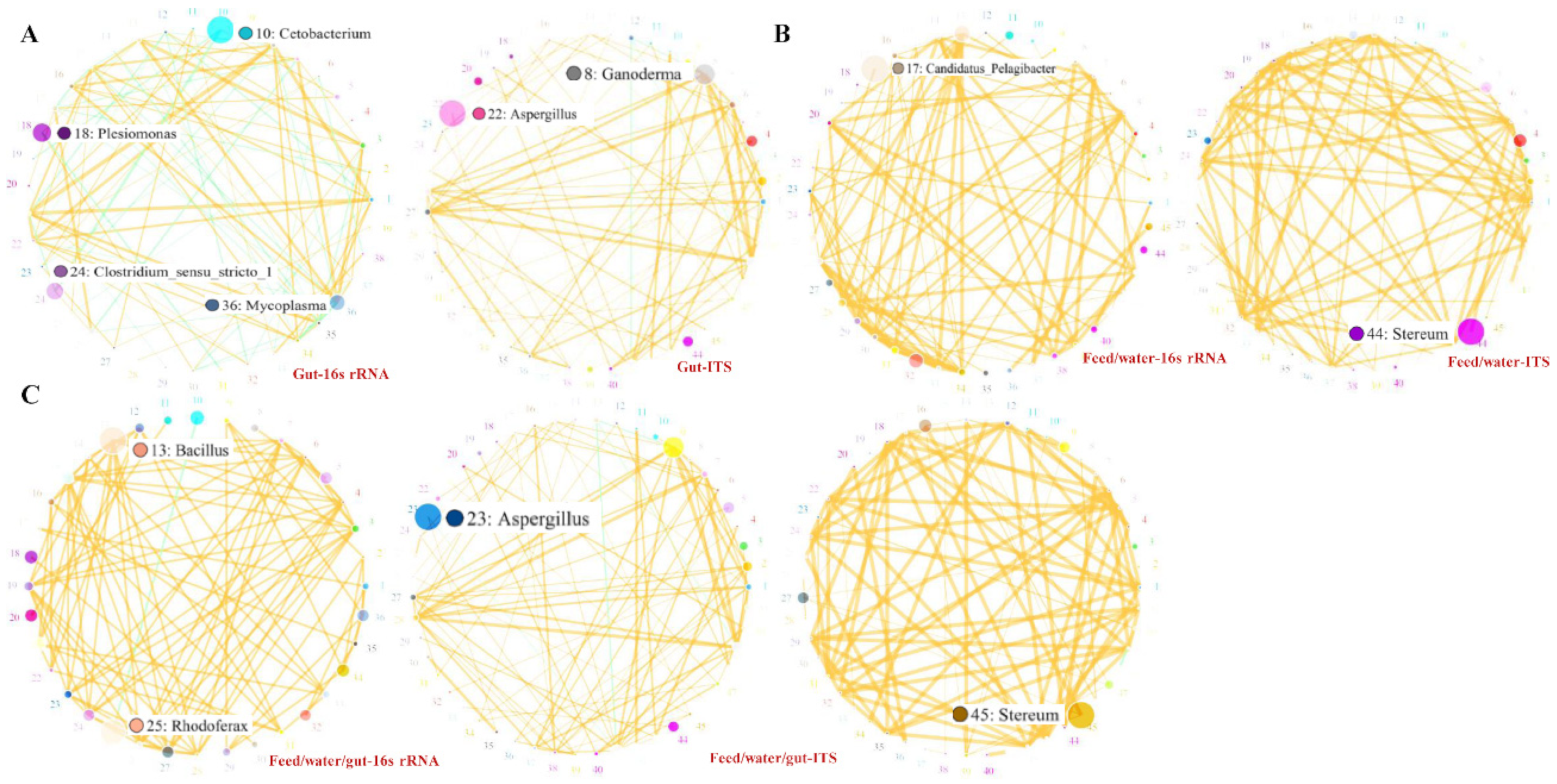
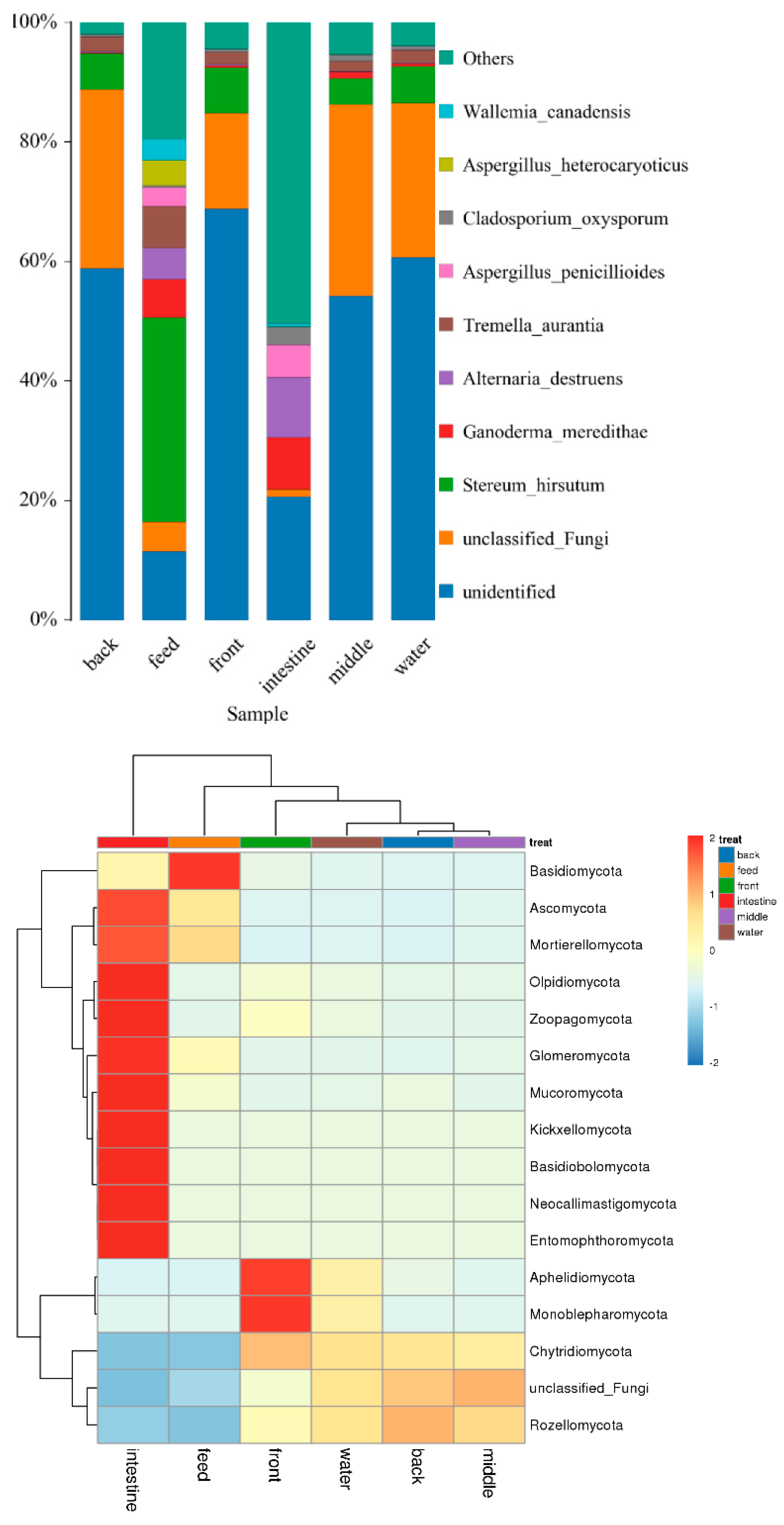
3.5. OTU Biomarkers and Their Related Analysis for the Associated Pathway
- (1)
- OTUs between pond commercial feed (PF) and iced fish feed (PI) groups
- (2)
- OTUs for using different culturing densities between pond (PF) and water tank commercial feed (WF) groups
- (3)
- OTUs by using different feeding strategies between pond commercial and iced fish combined feed (PFI), and pond commercial (PF) and iced fish (PI) feed, water tank (WF) groups
- (4)
- The pathogenic bacteria and fungi between groups and their related characteristic branches
- (5)
- The potential pathogenic bacteria/fungi source
4. Discussion
4.1. Changes for the Components of Fatty Acid
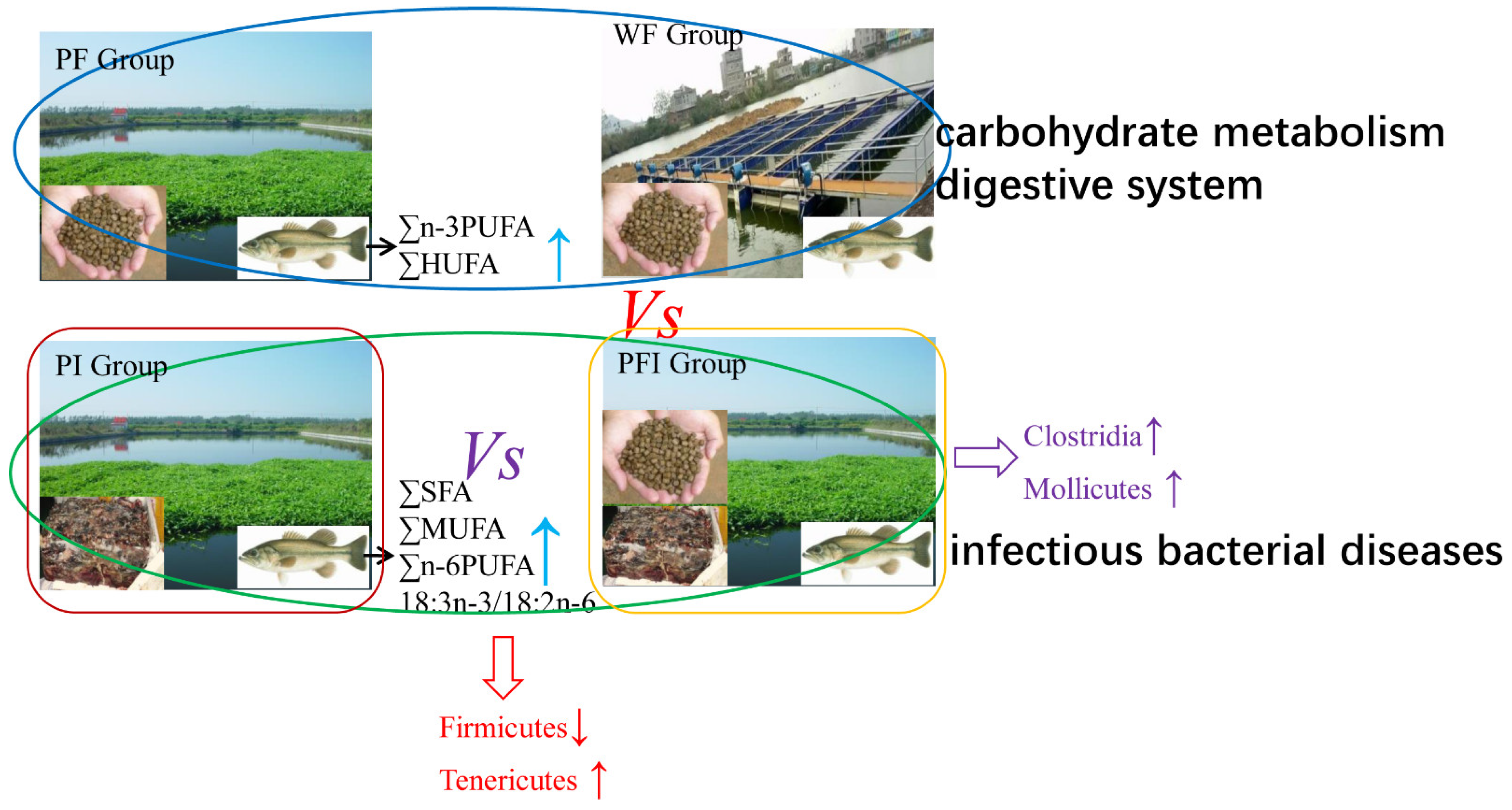
4.2. Different Feeding Strategies Shaping Body Composition, Gut Microbiota, and Enhancing Pathogenic Prevention
- (1)
- OTUs related to nutrient absorption efficiency between pond commercial feed (PF) and iced fish feed (PI) groups
- (2)
- OTUs related to environmental adaption by different culturing density between pond (PF) and water tank commercial feed (WF) groups
- (3)
- The pathogenic bacteria and fungi OTUs by different feeding strategies between pond commercial and iced fish combined feed (PFI) and other groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, J.; Ni, M.; Liu, M.; Lou, J.; Mi, G.; Gu, Z. Stocking density alters growth performance, serum biochemistry, digestive enzymes, immune response, and muscle quality of largemouth bass (Micropterus salmoides) in in-pond raceway system. Fish Physiol. Biochem. 2021, 47, 1697. [Google Scholar] [CrossRef]
- Huang, X.; Zhong, L.; Kang, Q.; Liu, S.; Feng, Y.; Geng, Y.; Chen, D.; Ou, Y.; Yang, S.; Yin, L.; et al. A high starch diet alters the composition of the intestinal microbiota of largemouth bass Micropterus salmoides, which may be associated with the development of enteritis. Front. Microbiol. 2021, 12, 696588. [Google Scholar] [CrossRef]
- Lin, M.; Zeng, C.; Jia, X.; Zhai, S.; Li, Z.; Ma, Y. The composition and structure of the intestinal microflora of Anguilla marmorata at different growth rates: A deep sequencing study. J. Appl. Microbiol. 2019, 126, 1340–1352. [Google Scholar] [CrossRef]
- Shi, C.; Zhao, H.; Zhai, X.; Chen, Y.; Lin, S. Linseed oil can decrease liver fat deposition and improve antioxidant ability of juvenile largemouth bass, Micropterus salmoides. Fish Physiol. Biochem. 2019, 45, 1513–1521. [Google Scholar] [CrossRef]
- Estadella, D.; da Penha Oller do Nascimento, C.M.; Oyama, L.M.; Ribeiro, E.B.; Dâmaso, A.R.; de Piano, A. Lipotoxicity: Effects of dietary saturated and transfatty acids. Mediat. Inflamm. 2013, 2013, 137579. [Google Scholar] [CrossRef]
- Kühn, G.; Pallauf, K.; Schulz, C.; Birringer, M.; Diaz-Rica, B.; de Pascual-Teresa, S.; Rimbach, G. Resveratrol modulates desaturase expression and fatty acid composition of cultured hepatocytes. Front. Nutr. 2018, 5, 106. [Google Scholar] [CrossRef]
- Sun, S.; Hua, X.; Deng, Y.; Zhang, Y.; Li, J.; Wu, Z.; Limbu, S.M.; Lu, D.; Yin, H.; Wang, G.; et al. Tracking pollutants in dietary fish oil: From ocean to table. Environ. Pollut. 2018, 240, 733–744. [Google Scholar] [CrossRef]
- Jiang, D.; Wu, Y.; Huang, D.; Ren, X.; Wang, Y. Effects of nutritional history on stress response in gibel carp (Carassius auratus gibelio) and largemouth bass (Micropterus salmoides). Comp. Biochem. Physiol. B 2017, 210, 9–17. [Google Scholar] [CrossRef]
- Zha, Y.; Eiler, A.; Johansson, F.; Svanbäck, R. Effects of predation stress and food ration on perch gut microbiota. Microbiome 2018, 6, 28. [Google Scholar] [CrossRef]
- Zheng, Y.; Wu, W.; Zhang, S.; Hu, G.; Qiu, L.; Bing, X.; Chen, J. Varieties of immunity activities and gut contents in tilapia with seasonal changes. Fish Shellfish Immunol. 2019, 90, 466–476. [Google Scholar] [CrossRef]
- Sutton, N.B.; Maphosa, F.; Morillo, J.A.; Abu Al-Soud, W.; Langenhoff, A.A.; Grotenhuis, T. Impact of long-term diesel contamination on soil microbial community structure. Appl. Environ. Microbiol. 2013, 79, 619–630. [Google Scholar] [CrossRef]
- Minamoto, T.; Uchii, K.; Takahara, T.; Kitayoshi, T.; Tsuji, S.; Yamanaka, H.; Doi, H. Nuclear internal transcribed spacer-1 as a sensitive genetic marker for environmental DNA studies in common carp Cyprinus carpio. Mol. Ecol. Resour. 2017, 17, 324–333. [Google Scholar] [CrossRef]
- Martín, M.; Almansa, E.; Cejas, J.; Bolaños, A.; Jerez, S.; Lorenzo, A. Effects of a diet lacking HUFA on lipid and fatty acid content of intestine and gills of male gilthead seabream (Sparus aurata L.) broodstock at different stages of the reproductive cycle. Fish Physiol. Biochem. 2011, 37, 935–949. [Google Scholar] [CrossRef]
- Li, Y.; Pang, Y.; Xiang, X.; Du, J.; Mai, K.; Ai, Q. Molecular cloning, characterization, and nutritional regulation of elovl6 in large yellow croaker (Larimichthys crocea). Int. J. Mol. Sci. 2019, 20, 1801. [Google Scholar] [CrossRef]
- Wei, J.; Guo, X.; Liu, H.; Chen, Y.; Wang, W. The variation profile of intestinal microbiota in blunt snout bream (Megalobrama amblycephala) during feeding habit transition. BMC Microbiol. 2018, 18, 99. [Google Scholar] [CrossRef]
- Li, X.; Yan, Q.; Ringø, E.; Wu, X.; He, Y.; Yang, D. The influence of weight and gender on intestinal bacterial community of wild largemouth bronze gudgeon (Coreius guichenoti, 1874). BMC Microbiol. 2016, 16, 191. [Google Scholar] [CrossRef]
- Yu, L.; Qiao, N.; Li, T.; Yu, R.; Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Dietary supplementation with probiotics regulates gut microbiota structure and function in Nile tilapia exposed to aluminum. PeerJ 2019, 7, e6963. [Google Scholar] [CrossRef]
- Li, T.; Long, M.; Ji, C.; Shen, Z.; Gatesoupe, F.J.; Zhang, X.; Zhang, Q.; Zhang, L.; Zhao, Y.; Liu, X.; et al. Alterations of the gut microbiome of largemouth bronze gudgeon (Coreius guichenoti) suffering from furunculosis. Sci. Rep. 2016, 6, 30606. [Google Scholar] [CrossRef]
- Pan, Z.; Li, L.; Shen, Z.; Chen, Y.; Li, M. Characterization of the microbiota in air- or vacuum-packed crisp grass carp (Ctenopharyngodon idella C. et V.) fillets by 16S rRNA PCR-denaturing gradient gel electrophoresis and high-throughput sequencing. J. Food Prot. 2018, 81, 1022–1029. [Google Scholar] [CrossRef]
- Campanaro, S.; Raga, R.; Squartini, A. Intermittent aeration of landfill simulation bioreactors: Effects on emissions and microbial community. Waste Manag. 2020, 117, 146–156. [Google Scholar] [CrossRef]
- Lu, X.; Han, B.; Deng, X.; Deng, S.; Zhang, Y.; Shen, P.; Hui, T.; Chen, R.; Li, X.; Zhang, Y. Pomegranate peel extract ameliorates the severity of experimental autoimmune encephalomyelitis via modulation of gut microbiota. Gut Microbes 2020, 12, 1857515. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, H.; Yang, H.; Li, J.; Xiao, S.; Hu, S.; Yan, F.; Xia, L.; Zhang, Y. Screening of a Plesiomonas shigelloides phage and study of the activity of its lysis system. Virus Res. 2021, 306, 198581. [Google Scholar] [CrossRef]
- Chen, W.; Chang, K.; Chen, J.; Zhao, X.; Gao, S. Dietary sodium butyrate supplementation attenuates intestinal inflammatory response and improves gut microbiota composition in largemouth bass (Micropterus salmoides) fed with a high soybean meal diet. Fish Physiol. Biochem. 2021, 47, 1805–1819. [Google Scholar] [CrossRef]
- Kent, M.L.; Wall, E.S.; Sichel, S.; Watral, V.; Stagaman, K.; Sharpton, T.J.; Guillemin, K. Pseudocapillaria tomentosa, Mycoplasma spp.; and intestinal lesions in experimentally infected zebrafish Danio rerio. Zebrafish 2021, 18, 207–220. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Q.; Lin, Y.; Hao, J.; Wang, S.; Zhang, J.; Li, A. Taxonomic and functional characteristics of the gill and gastrointestinal microbiota and its correlation with intestinal metabolites in new GIFT strain of farmed adult nile tilapia (Oreochromis niloticus). Microorganisms 2021, 9, 617. [Google Scholar] [CrossRef]
- Nguyen, C.D.H.; Amoroso, G.; Ventura, T.; Minich, J.J.; Elizur, A. Atlantic salmon (Salmo salar L., 1758) gut microbiota profile correlates with flesh pigmentation: Cause or effect? Mar. Biotechnol. 2020, 22, 786–804. [Google Scholar] [CrossRef]
- Wang, A.; Zhang, Z.; Ding, Q.; Yang, Y.; Bindelle, J.; Ran, C.; Zhou, Z. Intestinal Cetobacterium and acetate modify glucose homeostasis via parasympathetic activation in zebrafish. Gut Microbes 2021, 13, 1–15. [Google Scholar] [CrossRef]
- Ray, K. Gut microbiota: Married to our gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 555. [Google Scholar] [CrossRef]
- Ornelas-García, P.; Pajares, S.; Sosa-Jiménez, V.M.; Rétaux, S.; Miranda-Gamboa, R.A. Microbiome differences between river-dwelling and cave-adapted populations of the fish Astyanax mexicanus (De Filippi, 1853). PeerJ 2018, 6, e5906. [Google Scholar] [CrossRef]
- Xu, G.; Xing, W.; Li, T.; Xue, M.; Ma, Z.; Jiang, N.; Luo, L. Comparative study on the effects of different feeding habits and diets on intestinal microbiota in Acipenser baeri Brandt and Huso huso. BMC Microbiol. 2019, 19, 297. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Y.; Ringø, E.; Wang, X.; Gong, J.; Yang, D. Intestinal microbiome and its potential functions in bighead carp (Aristichthys nobilis) under different feeding strategies. PeerJ 2018, 6, e6000. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Y.; Yan, Q.; Ringø, E.; Yang, D. Do the intestinal microbiotas differ between paddlefish (Polyodon spathala) and bighead carp (Aristichthys nobilis) reared in the same pond? J. Appl. Microbiol. 2014, 117, 1245–1252. [Google Scholar] [CrossRef]
- Lin, S.; Jiang, Y.; Chen, Y.; Luo, L.; Doolgindachbaporn, S.; Yuangsoi, B. Effects of Astragalus polysaccharides (APS) and chitooligosaccharides (COS) on growth, immune response and disease resistance of juvenile largemouth bass, Micropterus salmoides. Fish Shellfish Immunol. 2017, 70, 40–47. [Google Scholar] [CrossRef]
- Molnár, T.; Csuvár, A.; Benedek, I.; Molnár, M.; Kabai, P. Domestication affects exploratory behaviour of pikeperch (Sander lucioperca L.) during the transition to pelleted food. PLoS ONE 2018, 13, e0196118. [Google Scholar] [CrossRef]
- Deng, Y.; Cheng, C.; Xie, J.; Liu, S.; Ma, H.; Feng, J.; Su, Y.; Guo, Z. Coupled changes of bacterial community and function in the gut of mud crab (Scylla paramamosain) in response to Baimang disease. AMB Express 2019, 9, 18. [Google Scholar] [CrossRef]
- Romero, J.; Navarrete, P. 16S rDNA-based analysis of dominant bacterial populations associated with early life stages of coho salmon (Oncorhynchus kisutch). Microb. Ecol. 2006, 51, 422–430. [Google Scholar] [CrossRef]
- McIlroy, S.J.; Starnawska, A.; Starnawski, P.; Saunders, A.M.; Nierychlo, M.; Nielsen, P.H.; Nielsen, J.L. Identification of active denitrifiers in full-scale nutrient removal wastewater treatment systems. Environ. Microbiol. 2016, 18, 50–64. [Google Scholar] [CrossRef]
- Chen, Z.; Ai, L.; Wang, J.; Ren, L.; Yu, Y.; Xu, J.; Chen, H.; Yu, J.; Li, M.; Qin, W.; et al. Probiotics Clostridium butyricum and Bacillus subtilis ameliorate intestinal tumorigenesis. Future Microbiol. 2015, 10, 1433–1445. [Google Scholar] [CrossRef]
- Wu, H.N.; Kawaguchi, C.; Nakane, D.; Miyata, M. “Mycoplasmal antigen modulation”, a novel surface variation suggested for a lipoprotein specifically localized on Mycoplasma mobile. Curr. Microbiol. 2012, 64, 433–440. [Google Scholar] [CrossRef]
- Iwashita, M.K.; Nakandakare, I.B.; Terhune, J.S.; Wood, T.; Ranzani-Paiva, M.J. Dietary supplementation with Bacillus subtilis, Saccharomyces cerevisiae and Aspergillus oryzae enhance immunity and disease resistance against Aeromonas hydrophila and Streptococcus iniae infection in juvenile tilapia Oreochromis niloticus. Fish Shellfish Immunol. 2015, 43, 60–66. [Google Scholar] [CrossRef]
- Shi, F.; Zi, Y.; Lu, Z.; Li, F.; Yang, M.; Zhan, F.; Li, Y.; Li, J.; Zhao, L.; Lin, L.; et al. Bacillus subtilis H2 modulates immune response, fat metabolism and bacterial flora in the gut of grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2020, 106, 8–20. [Google Scholar] [CrossRef]
- Minich, J.J.; Zhu, Q.; Xu, Z.Z.; Amir, A.; Ngochera, M.; Simwaka, M.; Allen, E.; Zidana, H.; Knight, R. Microbial effects of livestock manure fertilization on freshwater aquaculture ponds rearing tilapia (Oreochromis shiranus) and North African catfish (Clarias gariepinus). Microbiologyopen 2018, 7, e00716. [Google Scholar] [CrossRef]
- Feng, W.; Feng, W.; Ge, J.; Li, J.; Su, S.; Jia, R.; Yu, J.; Xu, P.; Tang, Y. Alterations of amino acid metabolism and intestinal microbiota in Chinese mitten crab (Eriocheir sinensis) fed on formulated diet and iced trash fish. Comp. Biochem. Physiol. D 2021, 40, 100924. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Name | PF (n = 7) | WF (n = 8) | PI (n = 7) | PFI (n = 8) | PI:PF (p) | WF:PF (p) | PFI:PF (p) | PFI:PI (p) | PFI:WF (p) |
|---|---|---|---|---|---|---|---|---|---|---|
| Phylum | Firmicutes | 0.2577 ± 0.3333 | 0.0240 ± 0.0180 | 0.0345 ± 0.0497 | 0.5600 ± 0.3001 | ## | ## | |||
| Fusobacteria | 0.5014 ± 0.2783 | 0.1553 ± 0.0908 | 0.8297 ± 0.2920 | 0.0373 ± 0.0820 | * | * | ** | ** | ||
| Tenericutes | 0.0575 ± 0.0634 | 0.4069 ± 0.3253 | 0.0004 ± 0.0004 | 0.0553 ± 0.1481 | ** | ** | ** | |||
| Class | Clostridia | 0.2481 ± 0.3294 | 0.0228 ± 0.0178 | 0.0117 ± 0.0113 | 0.5523 ± 0.3116 | ## | ## | |||
| Fusobacteria | 0.5014 ± 0.2783 | 0.1553 ± 0.0908 | 0.8297 ± 0.2920 | 0.0373 ± 0.0820 | * | * | ** | * | ||
| Mollicutes | 0.0575 ± 0.0634 | 0.4069 ± 0.3253 | 0.0004 ± 0.0004 | 0.0553 ± 0.1481 | ** | ** | ** | |||
| Order | Clostridiales | 0.2481 ± 0.3294 | 0.0228 ± 0.0178 | 0.0117 ± 0.0113 | 0.5523 ± 0.3116 | ## | ## | |||
| Fusobacteriales | 0.5014 ± 0.2783 | 0.1553 ± 0.0908 | 0.8297 ± 0.2920 | 0.0373 ± 0.0820 | * | * | ** | * | ||
| Mycoplasmatales | 0.0575 ± 0.0634 | 0.4069 ± 0.3253 | 0.0004 ± 0.0004 | 0.0553 ± 0.1481 | ** | ** | ** | |||
| Family | Clostridiaceae_1 | 0.1679 ± 0.3511 | 0.0202 ± 0.0167 | 0.0027 ± 0.0034 | 0.4681 ± 0.3159 | ** | ** | |||
| Fusobacteriaceae | 0.5014 ± 0.2783 | 0.1553 ± 0.0908 | 0.8297 ± 0.2920 | 0.0373 ± 0.0820 | * | * | ** | ** | ** | |
| Mycoplasmataceae | 0.0575 ± 0.0634 | 0.4069 ± 0.3253 | 0.0004 ± 0.0004 | 0.0553 ± 0.1481 | ** | ** | ## | |||
| Genus | Cetobacterium | 0.5014 ± 0.2783 | 0.1551 ± 0.0906 | 0.8297 ± 0.2919 | 0.0373 ± 0.0820 | * | * | ** | * | |
| Clostridium_sensu_stricto_1 | 0.1556 ± 0.3557 | 0.0182 ± 0.0160 | 0.0012 ± 0.0022 | 0.4659 ± 0.3193 | ** | ** | ||||
| Mycoplasma | 0.0575 ± 0.0634 | 0.4069 ± 0.3253 | 0.0004 ± 0.0004 | 0.0553 ± 0.1481 | ** | ** | ** | |||
| Species | Clostridium_perfringens_WAL-14572 | 0.0000 ± 0.0001 | 0.0013 ± 0.0028 | 0.0005 ± 0.0013 | 0.0807 ± 0.0661 | ## | ## | ## | ||
| uncultured_bacterium_g_Cetobacterium | 0.5014 ± 0.2783 | 0.1551 ± 0.0906 | 0.8297 ± 0.2919 | 0.0373 ± 0.0820 | * | * | ** | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Zhu, H.; Li, Q.; Xu, G. The Effects of Different Feeding Regimes on Body Composition, Gut Microbial Population, and Susceptibility to Pathogenic Infection in Largemouth Bass. Microorganisms 2023, 11, 1356. https://doi.org/10.3390/microorganisms11051356
Zheng Y, Zhu H, Li Q, Xu G. The Effects of Different Feeding Regimes on Body Composition, Gut Microbial Population, and Susceptibility to Pathogenic Infection in Largemouth Bass. Microorganisms. 2023; 11(5):1356. https://doi.org/10.3390/microorganisms11051356
Chicago/Turabian StyleZheng, Yao, Haojun Zhu, Quanjie Li, and Gangchun Xu. 2023. "The Effects of Different Feeding Regimes on Body Composition, Gut Microbial Population, and Susceptibility to Pathogenic Infection in Largemouth Bass" Microorganisms 11, no. 5: 1356. https://doi.org/10.3390/microorganisms11051356