
Antifungal Activity of Cell-Free Filtrate of Probiotic Bacteria Lactobacillus rhamnosus ATCC-7469 against Fungal Strains Isolated from a Historical Manuscript
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Historical Manuscript Studied
2.1.2. New Whatman Paper (Control)
2.2. Methods
2.2.1. Measurement of Deterioration Aspects
Visual Assessment
Environmental Scanning Electron Microscope (ESEM) Investigation
Measurement of Cellulose Crystallinity by XRD
2.2.2. Fungal Isolation and Identification
2.2.3. Enzyme Activity
2.2.4. Biocontrol of Fungal Growth by Probiotic Strain
Probiotic Bacterial Strain
Bacterial Growth and Extraction
Gas Chromatography–Mass Spectrometry (GC-MS) Analysis Conditions
Biocompatibility of Extracted Metabolites
Antifungal Activity of the Probiotic Crude Extract
2.2.5. Statistical Analysis
3. Results and Discussion
3.1. Analytical Techniques Used for Condition Assessment of the Studied Manuscript
3.1.1. Visual Assessment
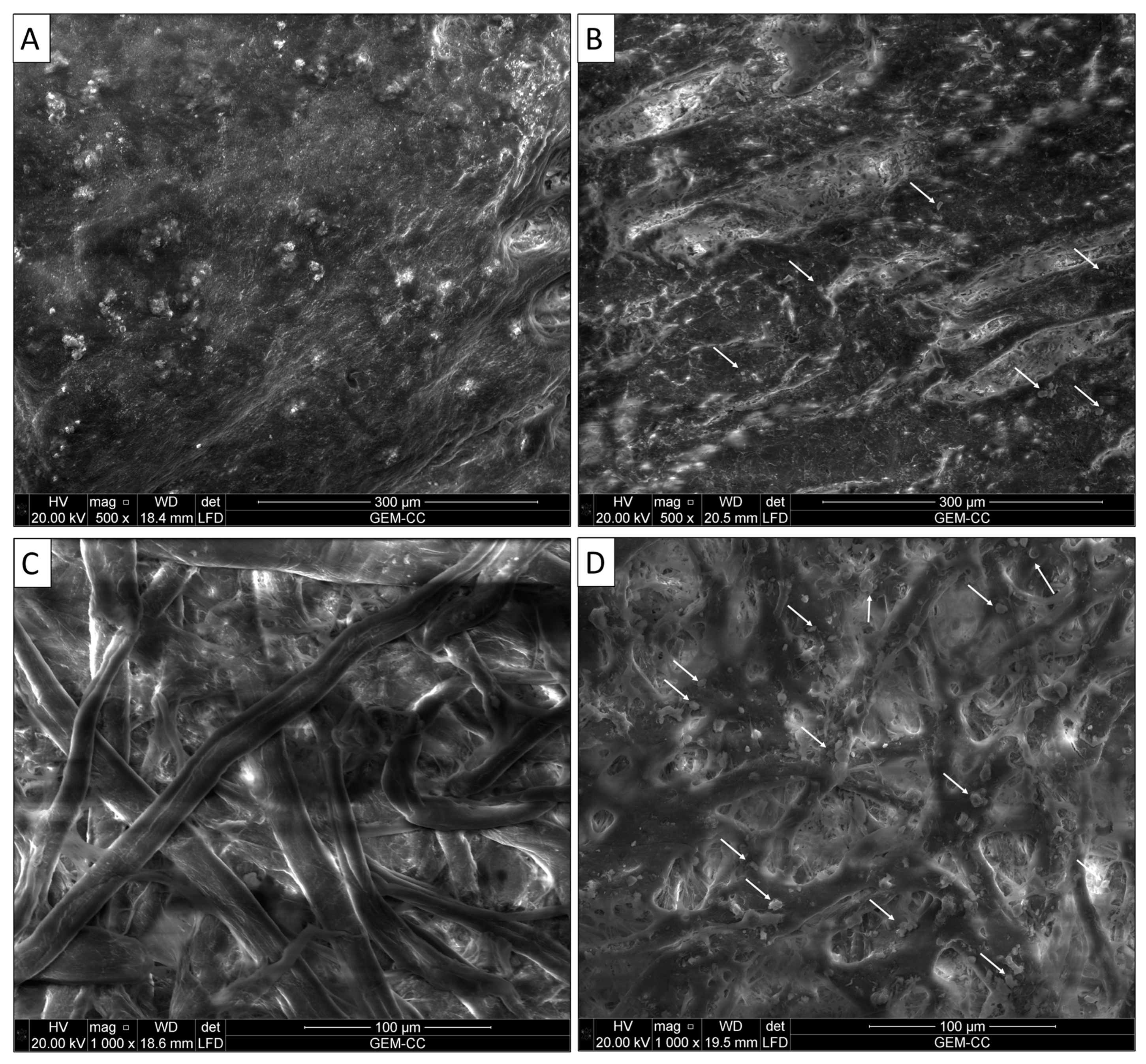
3.1.2. Investigation of the Surface Morphology by Scanning Electron Microscope (SEM)
3.1.3. X-ray Diffraction Analysis (XRD) for Measurement of Cellulose Crystallinity
3.2. Fungal Isolation and Identification
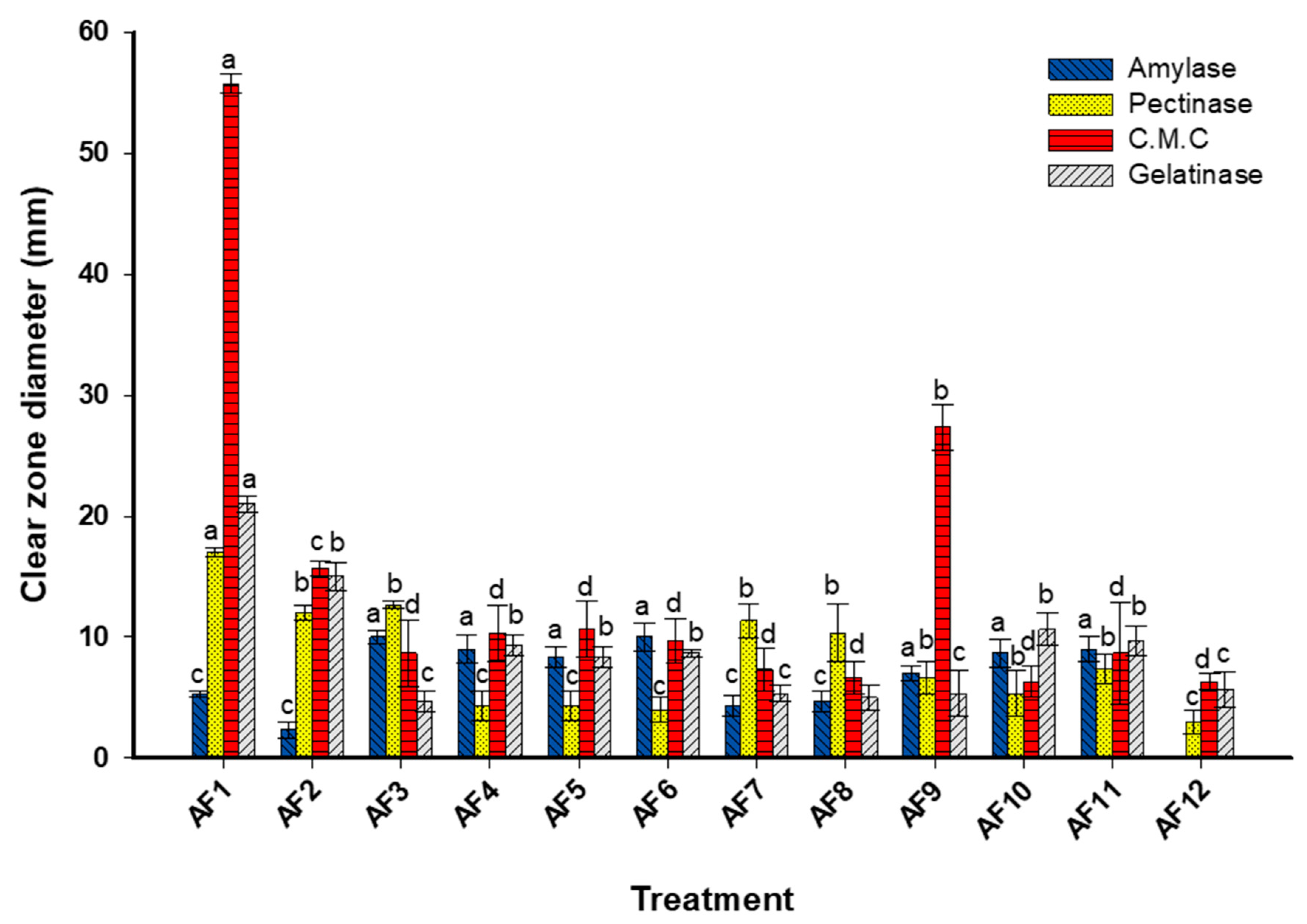
3.3. Enzymatic Activities
3.4. Biocontrol of Fungal Strains by Cell-Free Filtrate (CFF) of Probiotic Bacterial Strain L. rhamnosus
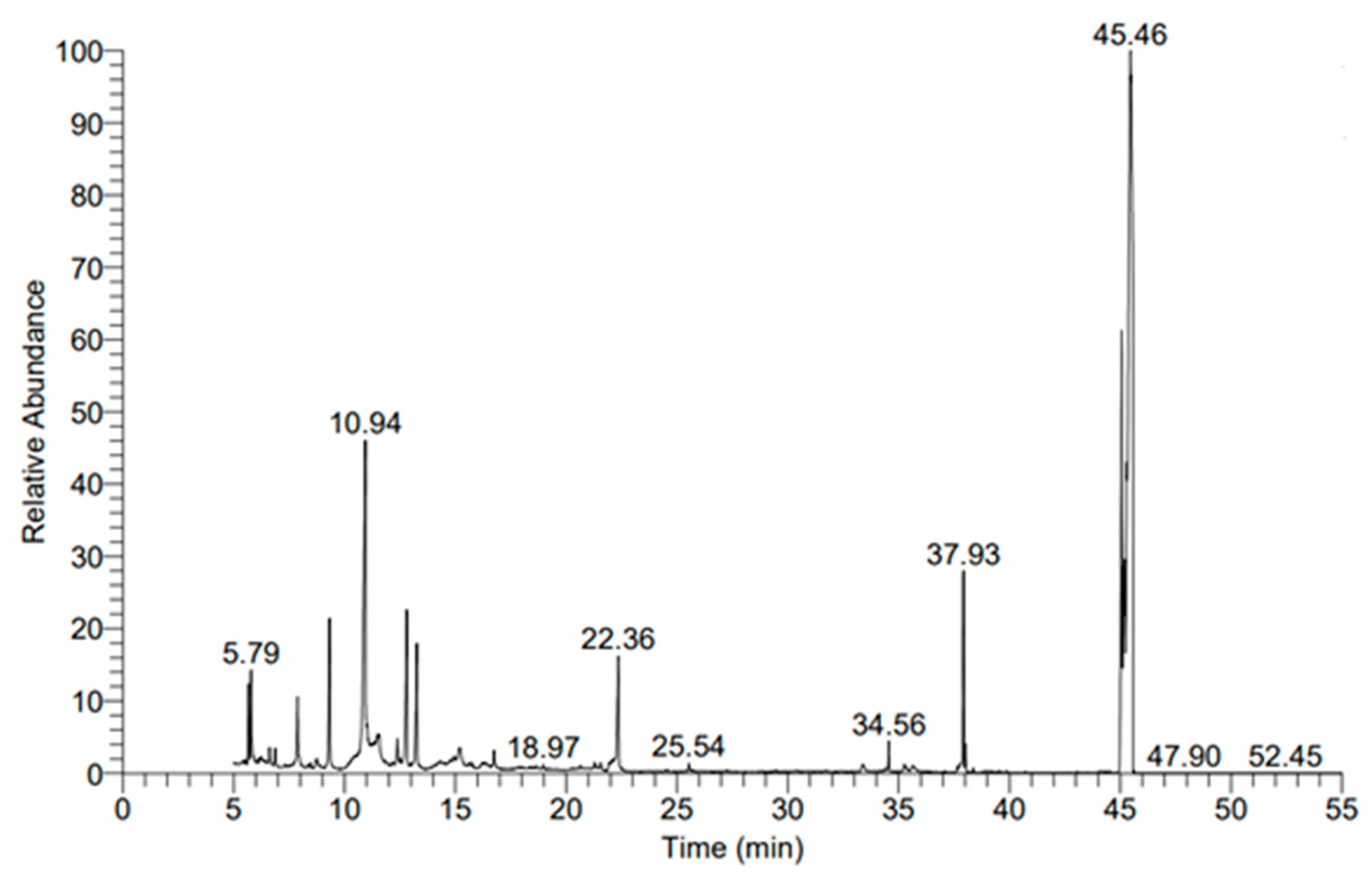
3.4.1. Metabolic Profile Analysis Using Gas Chromatography–Mass Spectrometry (GC-MS)
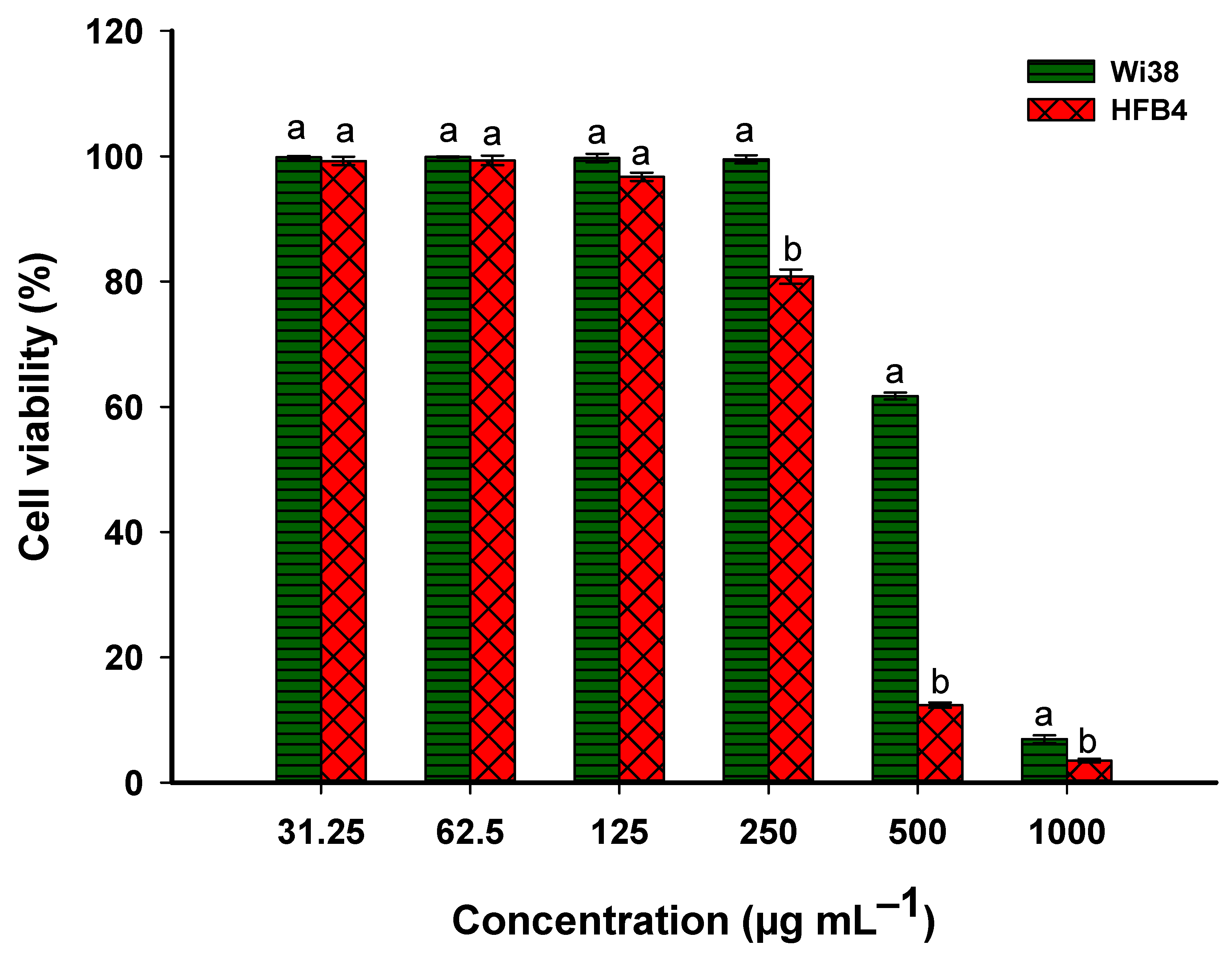
3.4.2. Biocompatibility of Metabolites of Probiotic Bacterial Strain
3.4.3. Antifungal Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Brand, A.J.; Quezada, V.; Gonzalez-Melo, C.; Bolaños-Barbosa, A.D.; Cruz, J.C.; Reyes, L.H. Novel Developments on Stimuli-Responsive Probiotic Encapsulates: From Smart Hydrogels to Nanostructured Platforms. Fermentation 2022, 8, 117. [Google Scholar] [CrossRef]
- Hossain, M.I.; Sadekuzzaman, M.; Ha, S.-D. Probiotics as potential alternative biocontrol agents in the agriculture and food industries: A review. Food Res. Int. 2017, 100, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Crook, B.; Burton, N.C. Indoor moulds, Sick Building Syndrome and building related illness. Fungal Biol. Rev. 2010, 24, 106–113. [Google Scholar] [CrossRef]
- Selim, M.T.; Salem, S.S.; Mohamed, A.A.; El-Gamal, M.S.; Awad, M.F.; Fouda, A. Biological Treatment of Real Textile Effluent Using Aspergillus flavus and Fusarium oxysporium and Their Consortium along with the Evaluation of Their Phytotoxicity. J. Fungi 2021, 7, 193. [Google Scholar] [CrossRef]
- Pinzari, F.; Gutarowska, B. Extreme Colonizers and Rapid Profiteers: The Challenging World of Microorganisms That Attack Paper and Parchment. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Joseph, E., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 79–113. [Google Scholar]
- Fouda, A.; Abdel-Nasser, M.; Khalil, A.M.A.; Hassan, S.E.-D.; Abdel-Maksoud, G. Investigate the role of fungal communities associated with a historical manuscript from the 17th century in biodegradation. NPJ Mater. Degrad. 2022, 6, 88. [Google Scholar] [CrossRef]
- Saada, N.S.; Abdel-Maksoud, G.; Abd El-Aziz, M.S.; Youssef, A.M. Evaluation and utilization of lemongrass oil nanoemulsion for disinfection of documentary heritage based on parchment. Biocatal. Agric. Biotechnol. 2020, 29, 101839. [Google Scholar] [CrossRef]
- Nitiu, D.S.; Mallo, A.C.; Saparrat, M.C.N. Fungal melanins that deteriorate paper cultural heritage: An overview. Mycologia 2020, 112, 859–870. [Google Scholar] [CrossRef]
- Oetari, A.; Natalius, A.; Komalasari, D.; Susetyo-Salim, T.; Sjamsuridzal, W. Fungal deterioration of old manuscripts of European paper origin. AIP Conf. Proc. 2018, 2023, 020156. [Google Scholar] [CrossRef]
- Ismail, Y.; Abdrabou, A.; Abdallah, M. A non-destructive analytical study and the conservation processes of Pharaoh Tutankhamun’s painted boat model. Int. J. Conserv. Sci. 2016, 7, 15–28. [Google Scholar]
- Beyene, D.; Chae, M.; Dai, J.; Danumah, C.; Tosto, F.; Demesa, A.G.; Bressler, D.C. Characterization of cellulase-treated fibers and resulting cellulose nanocrystals generated through acid hydrolysis. Materials 2018, 11, 1272. [Google Scholar] [CrossRef]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef] [PubMed]
- Diba, K.; Kordbacheh, P.; Mirhendi, S.; Rezaie, S.; Mahmoudi, M. Identification of Aspergillus species using morphological characteristics. Pak. J. Med. Sci. 2007, 23, 867. [Google Scholar]
- Frisvad, J.; Samson, R. Polyphasic taxonomy of Penicillium subgenus Penicillium. A guide to identif cation of food and airborne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 2004, 1–173. [Google Scholar]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Salem, S.S.; Hassan, S.E.-D.; El-Sadany, M.A.-H. Eco-friendly approach utilizing green synthesized nanoparticles for paper conservation against microbes involved in biodeterioration of archaeological manuscript. Int. Biodeterior. Biodegrad. 2019, 142, 160–169. [Google Scholar] [CrossRef]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.; Mahgoub, H.A.M.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; Elkelish, A.; et al. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E. Plant Growth-Promoting Endophytic Bacterial Community Inhabiting the Leaves of Pulicaria incisa (Lam.) DC Inherent to Arid Regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef]
- Vougiouklaki, D.; Tsironi, T.; Papaparaskevas, J.; Halvatsiotis, P.; Houhoula, D. Characterization of Lacticaseibacillus rhamnosus, Levilactobacillus brevis and Lactiplantibacillus plantarum Metabolites and Evaluation of Their Antimicrobial Activity against Food Pathogens. Appl. Sci. 2022, 12, 660. [Google Scholar] [CrossRef]
- Sharma, M.; Chandel, D.; Shukla, G. Antigenotoxicity and Cytotoxic Potentials of Metabiotics Extracted from Isolated Probiotic, Lactobacillus rhamnosus MD 14 on Caco-2 and HT-29 Human Colon Cancer Cells. Nutr. Cancer 2020, 72, 110–119. [Google Scholar] [CrossRef]
- Ding, M.; Wen, J.-H.; Guo, Z.-F.; Li, M.-N.; Wang, G.; Li, P.; Yang, H. Comprehensive chemical profiling and quantification of Shexiang Xintongning tablets by integrating liquid chromatography-mass spectrometry and gas chromatography-mass spectrometry. Arab. J. Chem. 2023, 16, 104527. [Google Scholar] [CrossRef]
- Hamza, M.F.; Fouda, A.; Wei, Y.; El Aassy, I.E.; Alotaibi, S.H.; Guibal, E.; Mashaal, N.M. Functionalized biobased composite for metal decontamination—Insight on uranium and application to water samples collected from wells in mining areas (Sinai, Egypt). Chem. Eng. J. 2022, 431, 133967. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Hamza, M.F.; Hamad, D.M.; Hamad, N.A.; Abdel-Rahman, A.A.H.; Fouda, A.; Wei, Y.; Guibal, E.; El-Etrawy, A.-A.S. Functionalization of magnetic chitosan microparticles for high-performance removal of chromate from aqueous solutions and tannery effluent. Chem. Eng. J. 2022, 428, 131775. [Google Scholar] [CrossRef]
- Abdel-Maksoud, G.; Abed al-Sameh Al-Shazly, E.E.; El-Amin, A.-R. Damage caused by insects during the mummification process: An experimental study. Archaeol. Anthropol. Sci. 2011, 3, 291–308. [Google Scholar] [CrossRef]
- Borrego, S.; Guiamet, P.; Vivar, I.; Battistoni, P. Fungi involved in biodeterioration of documents in paper and effect on substrate. Acta Microsc. 2018, 27, 37–44. [Google Scholar]
- Abdel-Maksoud, G.; Abdel-Nasser, M.; Sultan, M.H.; Eid, A.M.; Alotaibi, S.H.; Hassan, S.E.; Fouda, A. Fungal Biodeterioration of a Historical Manuscript Dating Back to the 14th Century: An Insight into Various Fungal Strains and Their Enzymatic Activities. Life 2022, 12, 1821. [Google Scholar] [CrossRef]
- Sebestyén, Z.; Badea, E.; Carsote, C.; Czégény, Z.; Szabó, T.; Babinszki, B.; Bozi, J.; Jakab, E. Characterization of historical leather bookbindings by various thermal methods (TG/MS, Py-GC/MS, and micro-DSC) and FTIR-ATR spectroscopy. J. Anal. Appl. Pyrolysis 2022, 162, 105428. [Google Scholar] [CrossRef]
- Carşote, C.; Budrugeac, P.; Decheva, R.; Haralampiev, N.S.; Miu, L.; Badea, E. Characterization of a byzantine manuscript by infrared spectroscopy and thermal analysis. Rev. Roum. Chim. 2014, 59, 429–436. [Google Scholar]
- Abdel-Maksoud, G.; Tharwat, N.; Gad, H. The role of fungi isolated from historical vegetable-tanned leather on the degradation of peptides and amino acids. J. Soc. Leather Technol. Chem 2014, 98, 1–9. [Google Scholar]
- Mosca Conte, A.; Pulci, O.; Knapik, A.; Bagniuk, J.; Del Sole, R.; Lojewska, J.; Missori, M. Role of cellulose oxidation in the yellowing of ancient paper. Phys. Rev. Lett. 2012, 108, 158301. [Google Scholar] [CrossRef]
- Abdel-Maksoud, G. Evaluation of wax or oil/fungicide formulations for preservation of vegetable-tanned leather artifacts. J. Soc. Leather Technol. Chem. 2006, 90, 58–67. [Google Scholar]
- Piñar, G.; Piombino-Mascali, D.; Maixner, F.; Zink, A.; Sterflinger, K. Microbial survey of the mummies from the Capuchin Catacombs of Palermo, Italy: Biodeterioration risk and contamination of the indoor air. FEMS Microbiol. Ecol. 2013, 86, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Jawahar, M.; Vani, K.; Babu, N.C. Leather species identification based on surface morphological characteristics using image analysis technique. J. Am. Leather Chem. Assoc. 2016, 111, 308–314. [Google Scholar]
- Ebsen, J.A.; Haase, K.; Larsen, R.; Sommer, D.V.P.; Brandt, L.Ø. Identifying archaeological leather—Discussing the potential of grain pattern analysis and zooarchaeology by mass spectrometry (ZooMS) through a case study involving medieval shoe parts from Denmark. J. Cult. Herit. 2019, 39, 21–31. [Google Scholar] [CrossRef]
- Michaelsen, A.; Piñar, G.; Pinzari, F. Molecular and Microscopical Investigation of the Microflora Inhabiting a Deteriorated Italian Manuscript Dated from the Thirteenth Century. Microb. Ecol. 2010, 60, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Sandy, M.; Manning, A.; Bollet, F. Changes in the crystallinity of cellulose in response to changes in relative humidity and acid treatment. Restaurator 2010, 31, 1–18. [Google Scholar] [CrossRef]
- Ioelovich, M. Adjustment of Hydrophobic Properties of Cellulose Materials. Polymers 2021, 13, 1241. [Google Scholar] [CrossRef] [PubMed]
- Boukir, A.; Fellak, S.; Doumenq, P. Structural characterization of Argania spinosa Moroccan wooden artifacts during natural degradation progress using infrared spectroscopy (ATR-FTIR) and X-Ray diffraction (XRD). Heliyon 2019, 5, e02477. [Google Scholar] [CrossRef]
- Samsudin, N.A.; Low, F.W.; Yusoff, Y.; Shakeri, M.; Tan, X.Y.; Lai, C.W.; Asim, N.; Oon, C.S.; Newaz, K.S.; Tiong, S.K.; et al. Effect of temperature on synthesis of cellulose nanoparticles via ionic liquid hydrolysis process. J. Mol. Liq. 2020, 308, 113030. [Google Scholar] [CrossRef]
- Lintang, W.; Susetyo-Salim, T.; Oetari, A.; Sjamsuridzal, W. Isolation and characterization of fungi from deteriorated old manuscripts from Banyumas, collection of Library of Universitas Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2021, 948, 012031. [Google Scholar] [CrossRef]
- Hassan, R. Analytical study of a manuscript,’Tafsir Al khazin’-the seventeenth century AD. Curr. Sci. Int. 2015, 4, 196–207. [Google Scholar]
- Shamsian, S.; Fata, A.; Mohajeri, M.; Ghazvini, K. Fungal Contaminations in Historical Manuscripts at Astan Quds Museum Library, Mashhad, Iran. Int. J. Agric. Biol. 2006, 8, 420–422. [Google Scholar]
- Oetari, A.; Susetyo-Salim, T.A.; Sjamsuridzal, W.; Suherman, E.A.; Monica, M.; Wongso, R.; Fitri, R.; Nurlaili, D.G.; Ayu, D.C.; Teja, T.P. Occurrence of fungi on deteriorated old dluwang manuscripts from Indonesia. Int. Biodeterior. Biodegrad. 2016, 114, 94–103. [Google Scholar] [CrossRef]
- Rachmania, M.; Oetari, A.; Fitri, R.; Susetyo-Salim, T.; Sjamsuridzal, W. Isolation and morphological characterization of fungi from deteriorated old Chinese manuscripts from Central Library Universitas Indonesia. AIP Conf. Proc. 2018, 2023, 020144. [Google Scholar]
- Padarik, E.T.; Oetari, A.; Fitri, R.; Susetyo-Salim, T.; Sjamsuridzal, W. Diversity of fungi from old Dluwang manuscripts from Mertasinga, Cirebon. AIP Conf. Proc. 2016, 1729, 020068. [Google Scholar]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Koul, B.; Upadhyay, H. Fungi-Mediated Biodeterioration of Household Materials, Libraries, Cultural Heritage and Its Control. In Fungi and Their Role in Sustainable Development: Current Perspectives; Gehlot, P., Singh, J., Eds.; Springer: Singapore, 2018; pp. 597–615. [Google Scholar]
- Sequeira, S.O.; Carvalho, H.P.d.; Mesquita, N.; Portugal, A.; Macedo, M.F. Fungal stains on paper: Is what you see what you get? Conserv. Património 2018, 32, 18–27. [Google Scholar] [CrossRef]
- Manente, S.; Micheluz, A.; Ganzerla, R.; Ravagnan, G.; Gambaro, A. Chemical and biological characterization of paper: A case study using a proposed methodological approach. Int. Biodeterior. Biodegrad. 2012, 74, 99–108. [Google Scholar] [CrossRef]
- Pinniger, D.B. Managing Pests in Paper-Based Collections; Preservation Advisory Centre, The British Library: London, UK, 2012. [Google Scholar]
- Taniwaki, M.H.; Pitt, J.I.; Magan, N. Aspergillus species and mycotoxins: Occurrence and importance in major food commodities. Curr. Opin. Food Sci. 2018, 23, 38–43. [Google Scholar] [CrossRef]
- Ma, X.; Hu, J.; Yu, Y.; Wang, C.; Gu, Y.; Cao, S.; Huang, X.; Wen, Y.; Zhao, Q.; Wu, R.; et al. Assessment of the pulmonary adaptive immune response to Cladosporium cladosporioides infection using an experimental mouse model. Sci. Rep. 2021, 11, 909. [Google Scholar] [CrossRef]
- Ferrándiz-Pulido, C.; Martin-Gomez, M.T.; Repiso, T.; Juárez-Dobjanschi, C.; Ferrer, B.; López-Lerma, I.; Aparicio, G.; González-Cruz, C.; Moreso, F.; Roman, A.; et al. Cutaneous infections by dematiaceous opportunistic fungi: Diagnosis and management in 11 solid organ transplant recipients. Mycoses 2019, 62, 121–127. [Google Scholar] [CrossRef]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Eid, A.M.; Barghoth, M.G.; El-Sadany, M.A.-H. Monitoring the effect of biosynthesized nanoparticles against biodeterioration of cellulose-based materials by Aspergillus niger. Cellulose 2019, 26, 6583–6597. [Google Scholar] [CrossRef]
- Sterflinger, K.; Pinzari, F. The revenge of time: Fungal deterioration of cultural heritage with particular reference to books, paper and parchment. Environ. Microbiol. 2012, 14, 559–566. [Google Scholar] [CrossRef] [PubMed]
- El Bergadi, F.; Laachari, F.; Elabed, S.; Mohammed, I.H.; Ibnsouda, S.K. Cellulolytic potential and filter paper activity of fungi isolated from ancients manuscripts from the Medina of Fez. Ann. Microbiol. 2014, 64, 815–822. [Google Scholar] [CrossRef]
- Anaya, M.; Borrego, S.F.; Gámez, E.; Castro, M.; Molina, A.; Valdés, O. Viable fungi in the air of indoor environments of the National Archive of the Republic of Cuba. Aerobiologia 2016, 32, 513–527. [Google Scholar] [CrossRef]
- Savković, Ž.; Stupar, M.; Unković, N.; Ivanović, Ž.; Blagojević, J.; Vukojević, J.; Ljaljević Grbić, M. In vitro biodegradation potential of airborne Aspergilli and Penicillia. Sci. Nat. 2019, 106, 8. [Google Scholar] [CrossRef]
- Shehata, M.G.; Badr, A.N.; El Sohaimy, S.A.; Asker, D.; Awad, T.S. Characterization of antifungal metabolites produced by novel lactic acid bacterium and their potential application as food biopreservatives. Ann. Agric. Sci. 2019, 64, 71–78. [Google Scholar] [CrossRef]
- Valerio, F.; Di Biase, M.; Lattanzio, V.M.; Lavermicocca, P. Improvement of the antifungal activity of lactic acid bacteria by addition to the growth medium of phenylpyruvic acid, a precursor of phenyllactic acid. Int. J. Food Microbiol. 2016, 222, 1–7. [Google Scholar] [CrossRef]
- Avand, A.; Akbari, V.; Shafizadegan, S. In Vitro Cytotoxic Activity of a Lactococcus lactis Antimicrobial Peptide Against Breast Cancer Cells. Iran. J. Biotechnol. 2018, 16, e1867. [Google Scholar] [CrossRef]
- Oliveira Silva, E.; Cruz de Carvalho, T.; Parshikov, I.A.; Alves dos Santos, R.; Silva Emery, F.; Jacometti Cardoso Furtado, N.A. Cytotoxicity of lapachol metabolites produced by probiotics. Lett. Appl. Microbiol. 2014, 59, 108–114. [Google Scholar] [CrossRef]
- Celebioglu, H.U.; Erden, Y.; Ozel, H.B. In vitro cytotoxic effects of lactobacilli grown with lime honey on human breast and colon cancer cells. Food Biosci. 2021, 41, 101020. [Google Scholar] [CrossRef]
- Dobrzyńska, I.; Skrzydlewska, E.; Figaszewski, Z.A. Changes in electric properties of human breast cancer cells. J. Membr. Biol. 2013, 246, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Yamaji-Hasegawa, A.; Tsujimoto, M. Asymmetric distribution of phospholipids in biomembranes. Biol. Pharm. Bull. 2006, 29, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Eckert, T.; Lutteke, T.; Hanstein, S.; Scheidig, A.; Bonvin, A.M.; Nifantiev, N.E.; Kozar, T.; Schauer, R.; Enani, M.A.; et al. Structure-Function Relationships of Antimicrobial Peptides and Proteins with Respect to Contact Molecules on Pathogen Surfaces. Curr. Top. Med. Chem. 2016, 16, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic Acid Bacteria as Antifungal and Anti-Mycotoxigenic Agents: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef]
- Thery, T.; Lynch, K.M.; Arendt, E.K. Natural Antifungal Peptides/Proteins as Model for Novel Food Preservatives. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1327–1360. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Algboory, H.L.; Kadum, H.; Mohammed, N.K.; Saari, N.; Hassan, Z.; Meor Hussin, A.S. Antifungal activity determination for the peptides generated by Lactobacillus plantarum TE10 against Aspergillus flavus in maize seeds. Food Control 2020, 109, 106898. [Google Scholar] [CrossRef]
- Quattrini, M.; Liang, N.; Fortina, M.G.; Xiang, S.; Curtis, J.M.; Gänzle, M. Exploiting synergies of sourdough and antifungal organic acids to delay fungal spoilage of bread. Int. J. Food Microbiol. 2019, 302, 8–14. [Google Scholar] [CrossRef]
- Poornachandra Rao, K.; Deepthi, B.V.; Rakesh, S.; Ganesh, T.; Achar, P.; Sreenivasa, M.Y. Antiaflatoxigenic Potential of Cell-Free Supernatant from Lactobacillus plantarum MYS44 Against Aspergillus parasiticus. Probiotics Antimicrob. Proteins 2019, 11, 55–64. [Google Scholar] [CrossRef]
- Ryan, L.A.M.; Zannini, E.; Dal Bello, F.; Pawlowska, A.; Koehler, P.; Arendt, E.K. Lactobacillus amylovorus DSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food Microbiol. 2011, 146, 276–283. [Google Scholar] [CrossRef]
- Deepthi, B.V.; Poornachandra Rao, K.; Chennapa, G.; Naik, M.K.; Chandrashekara, K.T.; Sreenivasa, M.Y. Antifungal Attributes of Lactobacillus plantarum MYS6 against Fumonisin Producing Fusarium proliferatum Associated with Poultry Feeds. PLoS ONE 2016, 11, e0155122. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [PubMed]
- Nader-Macías, M.E.F.; De Gregorio, P.R. Chapter 18—Probiotics and urogenital health. In Probiotics; Brandelli, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 355–388. [Google Scholar]
- Goh, Y.J.; Klaenhammer, T.R. Genetic mechanisms of prebiotic oligosaccharide metabolism in probiotic microbes. Annu. Rev. Food Sci. Technol. 2015, 6, 137–156. [Google Scholar] [CrossRef] [PubMed]
- Iñigo, M.; Del Pozo, J.L. Fungal biofilms: From bench to bedside. Rev. Esp. Quimioter. 2018, 31 (Suppl. 1), 35–38. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT | Area % | Probability | Compound Name | Molecular Weight | Molecular Formula | Library |
|---|---|---|---|---|---|---|
| 5.67 | 1.37 | 79.78 | Acetic acid, butyl ester (CAS) | 116 | C4H10O2 | Wiley9 |
| 5.79 | 1.87 | 43.80 | 1,3(R)Dihydroxybutane | 90 | C4H10O2 | Wiley9 |
| 8.66 | Diethylene glycol | 106 | C4H10O3 | nist_ms ms | ||
| 7.65 | 2Ethoxyethanol | 90 | C4H10O2 | nist_ms ms | ||
| 6.87 | 0.45 | 85.41 | 3,3Dimethoxy2butan one | 132 | C6H12O3 | Wiley9 |
| 7.88 | 1.78 | 10.63 | N-Acetyl glycine | 117 | C4H7NO3 | nist_ms ms |
| 10.22 | D-(-) Lactic acid | 90 | C3H6O3 | |||
| L-(+)-Lactic acid | ||||||
| 9.32 | 3.59 | 70.99 | Mono-Ethyl-malonate | 132 | C5H8O4 | nist_ms ms |
| 9.32 | 3.59 | 14.44 | Oxalacetic acid | 132 | C4H4O5 | |
| 10.93 | 9.58 | 41.53 | 2Butanone, 3hydroxy-(CAS) | 88 | C4H8O2 | Wiley9 |
| 8.68 | Methyl glyoxal | 72 | C3H4O2 | mainlib | ||
| 11.57 | 0.96 | 24.73 | Propanoic acid, 2-hydroxy-(CAS) | 90 | C3H6O3 | Wiley9 |
| (S)-2-Hydroxypropanoic acid | ||||||
| L-Lactic acid | mainlib | |||||
| 12.39 | 0.83 | 35.52 | 3-(3-chlorobutyl) isoquinolin-1(2H)-one | 235 | C13H14ClNO | Wiley9 |
| 22.23 | 3-Pentanone, 1,5dimethoxy-(CAS) | 146 | C7H14O3 | |||
| 3-Pentanone, 1,5-dimethoxy- | mainlib | |||||
| 12.81 | 4.22 | 79.32 | 3-(t-Butoxy)-2-methylpropan-1-ol | 144 | C8H16O2 | Wiley9 |
| 5.49 | 2(S)-acetoxy-succinic anhydride | 158 | C6H6O5 | |||
| 5.28 | (Z)-3-Tridecenal Diisopropyl Acetal | 298 | C19H38O2 | |||
| 13.26 | 3.30 | 18.91 | 2-Butanone, 4-(acetyloxy)-(CAS) | 130 | C6H10O3 | Wiley9 |
| 16.71 | 2-Butanol, 3-Methyl, Acetate | C7H14O2 | ||||
| 7.10 | 5-Hexen-2-one (CAS) | 98 | C6H10O | |||
| 15.20 | 0.66 | 16.61 | Propanoic acid,2-ydroxy-, Methyl ester (CAS) | 104 | C4H8O3 | Wiley9 |
| 11.40 | Acetic acid, methoxy-, ethyl ester (CAS) | 118 | C5H10O3 | |||
| 5.87 | 15-Crown-5 | 220 | C10H20O5 | mainlib | ||
| 16.75 | 0.48 | 1.85 | Phenyllacetic acid | 136 | C8H8O2 | nist_ms ms |
| 1.35 | 2,6, Dimethyl-6-(phenyl methoxy)-2-heptene | 232 | C16H24O | Wiley9 | ||
| 4-benzyloxy-3,5-dichloro-2-hydroxy-6-pentyl-benzoic acid | 347 | C19H20ClO4 | ||||
| 22.35 | 3.11 | 36.33 | Benzene-propanoic acid, à-hydroxy-, methyl ester | 180 | C10H12O3 | Wiley9 |
| Benzene-propanoic acid, à-hydroxy-, methyl ester (CAS) | ||||||
| 34.56 | 0.63 | 40.79 | Hexadecanoic acid, methyl ester (CAS) | 270 | C17H34O2 | Wiley9 |
| 37.93 | 3.58 | 10.03 | cis-13-Octadecenoic acid, methyl ester | 296 | C19H36O2 | mainlib |
| 8.86 | 9-Octadecenoic acid (Z)-, methyl ester | |||||
| 7.48 | trans-13-Octadecenoic acid, methyl ester | |||||
| 45.06 | 9.91 | 31.27 | 1,3-Benzenediamine, 2-methyl-5-nitro-(CAS) | 167 | C7H9N3O2 | Wiley9 |
| 11.42 | 4-(2,2-Dimethylpropyl)-2,2-dimethyl-3-methyl-netetrahydrofuran | 182 | C12H22O | |||
| 10.53 | 7-Chloro-2-methylfuro [3,2-b] pyridine | 167 | C8H6ClNO | |||
| 45.14 | 2.63 | 27.62 | 9,9-Dimethyl-8, 10-diox-apentacyclo [5.3.0.0(2,5).0(3,5).0(3,6)] decane | 166 | C10H14O2 | Wiley9 |
| 12.58 | 4-(2-Propen-1-yloxy) benzene amine | 149 | C9H11NO | |||
| 6.11 | 3-(5-’Formyl-2-furyl)-2-propenal | 150 | C8H6O3 | |||
| 45.20 | 2.28 | 18.26 | 9,9-Dimethyl-8,10-diox apenta-cyclo [5.3.0.0(2,5).0(3,5).0(3,6)]decane | 166 | C10H14O2 | Wiley9 |
| 16.85 | Methyl 1,3dihydro2Hisobenz ofuran-4-carboxylate | 178 | C10H10O3 | |||
| 12.57 | Benzoic acid, 4-formyl-, ethyl ester (CAS) | |||||
| 45.29 | 0.99 | 32.71 | Pyrido [2,1-a] isoindolium Chloride | 203 | C12H10ClN | Wiley9 |
| Pyrido [2,1-a] isoindolium Bromide | 247 | C12H10BrN | ||||
| 6.78 | 2-Carboxy-4-methylbicyclo [2.2.2]oct-2-en-1-ol | 182 | C10H14O3 | |||
| 45.49 | 47.78 | 81.24 | 8-Phenylacenaphtho [1,2-b] pyridine | 279 | C21H13N | Wiley9 |
| 7.01 | 4-Amino5-cyano-6-(4′methoxyphenyl)-1-methyl-2,3-dihydropyrrolo [2,3-b] pyridine | 280 | C16H16N4O | |||
| 10-Iodo-nido-7,8,9-phosphadicarbaborane | C2H10B8IP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Nasser, M.; Abdel-Maksoud, G.; Eid, A.M.; Hassan, S.E.-D.; Abdel-Nasser, A.; Alharbi, M.; Elkelish, A.; Fouda, A. Antifungal Activity of Cell-Free Filtrate of Probiotic Bacteria Lactobacillus rhamnosus ATCC-7469 against Fungal Strains Isolated from a Historical Manuscript. Microorganisms 2023, 11, 1104. https://doi.org/10.3390/microorganisms11051104
Abdel-Nasser M, Abdel-Maksoud G, Eid AM, Hassan SE-D, Abdel-Nasser A, Alharbi M, Elkelish A, Fouda A. Antifungal Activity of Cell-Free Filtrate of Probiotic Bacteria Lactobacillus rhamnosus ATCC-7469 against Fungal Strains Isolated from a Historical Manuscript. Microorganisms. 2023; 11(5):1104. https://doi.org/10.3390/microorganisms11051104
Chicago/Turabian StyleAbdel-Nasser, Mahmoud, Gomaa Abdel-Maksoud, Ahmed M. Eid, Saad El-Din Hassan, Aya Abdel-Nasser, Maha Alharbi, Amr Elkelish, and Amr Fouda. 2023. "Antifungal Activity of Cell-Free Filtrate of Probiotic Bacteria Lactobacillus rhamnosus ATCC-7469 against Fungal Strains Isolated from a Historical Manuscript" Microorganisms 11, no. 5: 1104. https://doi.org/10.3390/microorganisms11051104