


Rhizobacterial Colonization and Management of Bacterial Speck Pathogen in Tomato by Pseudomonas spp.

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemotaxis Activity of PGPRs
2.1.1. Collecting Root Exudates
2.1.2. Chemotactic Response of Bacterial Isolates to Organic Acids in a “Drop” Assay
2.1.3. Chemotaxis Assay (Capillary Assay)
2.2. Induction Treatments with PGPRs
2.3. Assessment of Defense-Related Enzymes
2.4. Molecular Investigation
2.5. Plant Growth under Field Experiments
2.6. Statistical Analysis
3. Results
3.1. Chemotaxis Activity of PGPRs
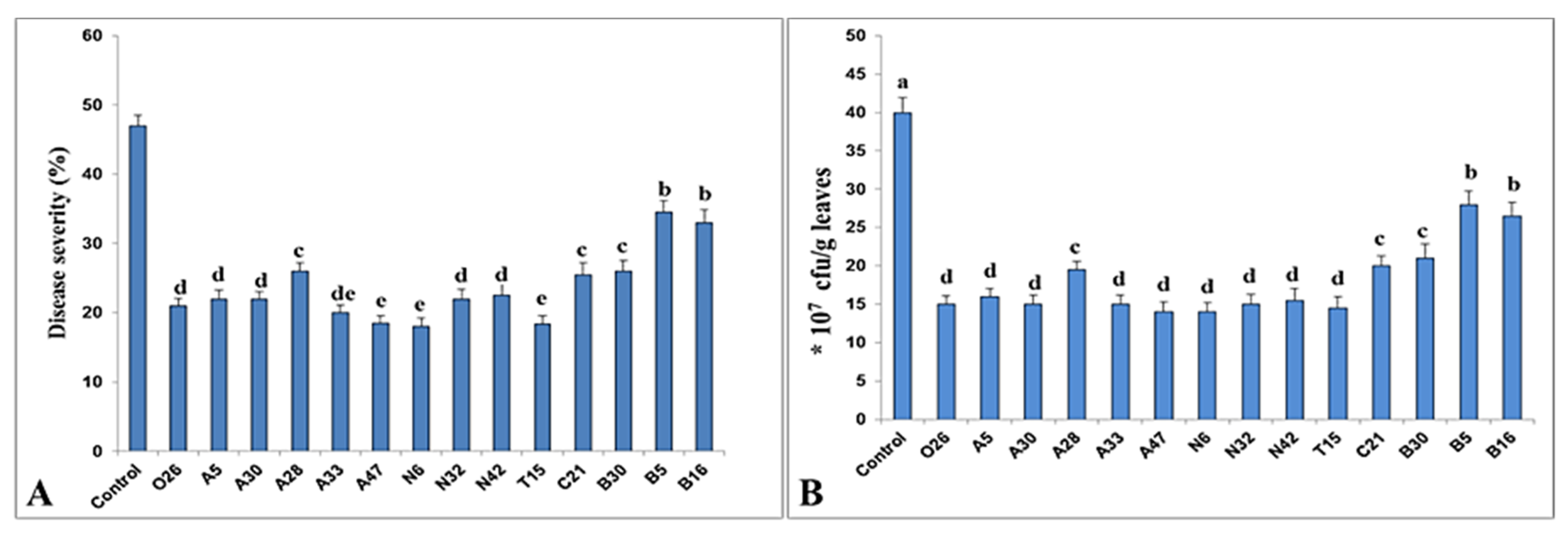
3.2. Induction of Systemic Resistance against Pst by PGPR
3.3. Effects of PGPR Treatments on Enzyme Activities in Plants Infected with Pst
3.4. Effects of PGPR Treatments on Plant Growth and Yield
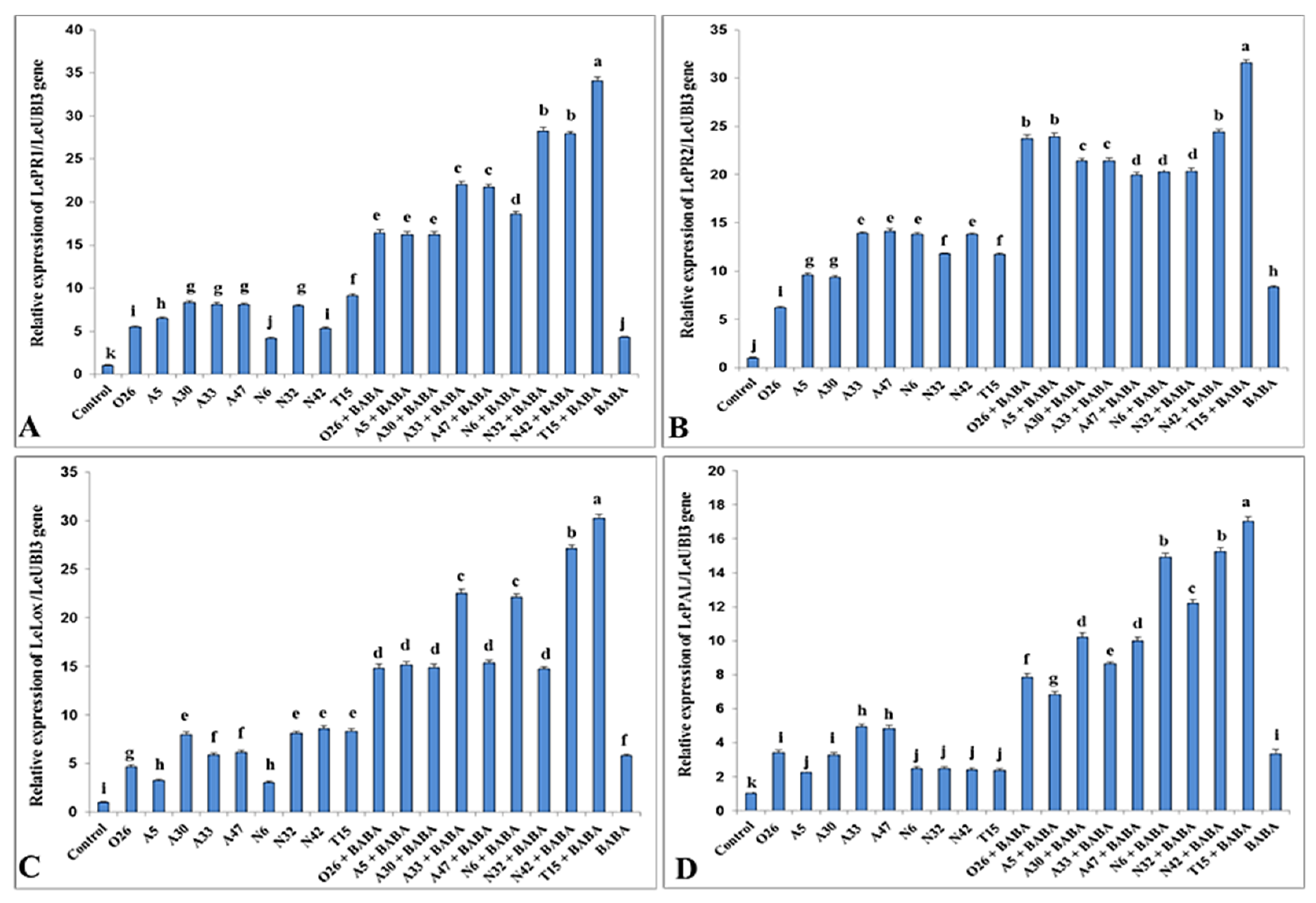
3.5. Molecular Analysis of Defense Genes by Real-Time PCR
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Elsawy, M.M.; Ismail, I.A. Mechanism of resistance to Cucumber mosaic virus elicited by inoculation with Bacillus subtilis subsp. subtilis. Pest Manag. Sci. 2021, 78, 86–94. [Google Scholar] [CrossRef] [PubMed]
- El-Sawah, A.M.; El-Keblawy, A.; Ali, D.F.I.; Ibrahim, H.M.; El-Sheikh, M.A.; Sharma, A.; Hamoud, Y.A.; Shaghaleh, H.; Brestic, M.; Skalicky, M.; et al. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Enhance Soil Key Enzymes, Plant Growth, Seed Yield, and Qualitative Attributes of Guar. Agriculture 2021, 11, 194. [Google Scholar] [CrossRef]
- Vocciante, M.; Grifoni, M.; Fusini, D.; Petruzzelli, G.; Franchi, E. The Role of Plant Growth-Promoting Rhizobacteria (PGPR) in Mitigating Plant’s Environmental Stresses. Appl. Sci. 2022, 12, 1231. [Google Scholar] [CrossRef]
- Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle. Biology 2021, 10, 158. [Google Scholar] [CrossRef]
- Oubohssaine, M.; Sbabou, L.; Aurag, J. Native Heavy Metal-Tolerant Plant Growth Promoting Rhizobacteria Improves Sulla spinosissima (L.) Growth in Post-Mining Contaminated Soils. Microorganisms 2022, 10, 838. [Google Scholar] [CrossRef]
- Gashash, E.A.; Osman, N.A.; Alsahli, A.A.; Hewait, H.M.; Ashmawi, A.E.; Alshallash, K.S.; El-Taher, A.M.; Azab, E.S.; El-Raouf, H.S.A.; Ibrahim, M.F.M. Effects of Plant-Growth-Promoting Rhizobacteria (PGPR) and Cyanobacteria on Botanical Characteristics of Tomato (Solanum lycopersicon L.) Plants. Plants 2022, 11, 2732. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, B.; Yu, Q. Genetic Engineering-Facilitated Coassembly of Synthetic Bacterial Cells and Magnetic Nanoparticles for Efficient Heavy Metal Removal. ACS Appl. Mater. Interfaces 2020, 12, 22948–22957. [Google Scholar] [CrossRef]
- Kumar, R.; Swapnil, P.; Meena, M.; Selpair, S.; Yadav, B.G. Plant Growth-Promoting Rhizobacteria (PGPR): Approaches to Alleviate Abiotic Stresses for Enhancement of Growth and Development of Medicinal Plants. Sustainability 2022, 14, 15514. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, Y.; Guo, L.; Yang, L.; Wang, J.; Liang, C.; Huang, J. The Effect of Banana Rhizosphere Chemotaxis and Chemoattractants on Bacillus velezensis LG14-3 Root Colonization and Suppression of Banana Fusarium Wilt Disease. Sustainability 2023, 15, 351. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Srivastava, A.K.; Rajput, V.D.; Chauhan, P.K.; Bhojiya, A.A.; Jain, D.; Chaubey, G.; Dwivedi, P.; Sharma, B.; Minkina, T. Root Exudates: Mechanistic Insight of Plant Growth Promoting Rhizobacteria for Sustainable Crop Production. Front. Microbiol. 2022, 13, 916488. [Google Scholar] [CrossRef]
- Feng, H.; Fu, R.; Hou, X.; Lv, Y.; Zhang, N.; Liu, Y.; Xu, Z.; Miao, Y.; Krell, T.; Shen, Q.; et al. Chemotaxis of Beneficial Rhizobacteria to Root Exudates: The First Step towards Root–Microbe Rhizosphere Interactions. Int. J. Mol. Sci. 2021, 22, 6655. [Google Scholar] [CrossRef] [PubMed]
- Tsotetsi, T.; Nephali, L.; Malebe, M.; Tugizimana, F. Bacillus for Plant Growth Promotion and Stress Resilience: What Have We Learned? Plants 2022, 11, 2482. [Google Scholar] [CrossRef] [PubMed]
- Jamil, F.; Mukhtar, H.; Fouillaud, M.; Dufossé, L. Rhizosphere Signaling: Insights into Plant–Rhizomicrobiome Interactions for Sustainable Agronomy. Microorganisms 2022, 10, 899. [Google Scholar] [CrossRef]
- Antoszewski, M.; Mierek-Adamska, A.; Dąbrowska, G.B. The Importance of Microorganisms for Sustainable Agriculture—A Review. Metabolites 2022, 12, 1100. [Google Scholar] [CrossRef]
- De Weert, S.; Vermeiren, H.; Mulders, I.H.M.; Kuiper, I.; Hendrickx, N.; Bloemberg, G.V.; Vanderleyden, J.; De Mol, R.; Lug-tenberg, B.J.J. Flagella-driven chemotaxis towards exudate components is an important trait for tomato root colonization by Pseudomonas fluorescens. Mol. Plant Microbe. Interact. 2002, 15, 1173–1180. [Google Scholar] [CrossRef]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-Secreted Malic Acid Recruits Beneficial Soil Bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef]
- Micallef, S.A.; Channer, S.; Shiaris, M.P.; Colón-Carmona, A. Plant age and genotype impact the progression of bacterial community succession in the Arabidopsis rhizosphere. Plant Signal. Behav. 2009, 4, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Huang, R.; Wang, J.; Wang, C.; Liu, R.; Zhang, H.; Deng, M.; Li, S.; Li, X.; Tang, R.; et al. Increase in Cd Tolerance through Seed-Borne Endophytic Fungus Epichloë gansuensis Affected Root Exudates and Rhizosphere Bacterial Community of Achnatherum inebrians. Int. J. Mol. Sci. 2022, 23, 13094. [Google Scholar] [CrossRef]
- Broz, A.K.; Manter, D.K.; Vivanco, J.M. Soil fungal abundance and diversity: Another victim of the invasive plant Centaurea maculosa. ISME J. 2007, 1, 763–765. [Google Scholar] [CrossRef]
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Rhizosphere Colonization Determinants by Plant Growth-Promoting Rhizobacteria (PGPR). Biology 2021, 10, 475. [Google Scholar] [CrossRef]
- Xu, Z.; Shao, J.; Li, B.; Yan, X.; Shen, Q.; Zhang, R. Contribution of Bacillomycin D in Bacillus amyloliquefaciens SQR9 to Antifungal Activity and Biofilm Formation. Appl. Environ. Microbiol. 2013, 79, 808–815. [Google Scholar] [CrossRef]
- Zhang, N.; Wu, K.; He, X.; Li, S.-Q.; Zhang, Z.-H.; Shen, B.; Yang, X.-M.; Zhang, R.-F.; Huang, Q.-W.; Shen, Q.-R. A new bioorganic fertilizer can effectively control banana wilt by strong colonization with Bacillus subtilis N11. Plant Soil 2011, 344, 87–97. [Google Scholar] [CrossRef]
- Butsenko, L.; Pasichnyk, L.; Kolomiiets, Y.; Kalinichenko, A. The Effect of Pesticides on the Tomato Bacterial Speck Disease Pathogen Pseudomonas Syringae pv. Tomato. Appl. Sci. 2020, 10, 3263. [Google Scholar] [CrossRef]
- Bazzarelli, F.; Mazzei, R.; Papaioannou, E.; Giannakopoulos, V.; Roberts, M.R.; Giorno, L. Biorefinery of Tomato Leaves by Integrated Extraction and Membrane Processes to Obtain Fractions That Enhance Induced Resistance against Pseudomonas syringae Infection. Membranes 2022, 12, 585. [Google Scholar] [CrossRef] [PubMed]
- Nabi, R.B.S.; Rolly, N.K.; Tayade, R.; Khan, M.; Shahid, M.; Yun, B.-W. Enhanced Resistance of atbzip62 against Pseudomonas syringae pv. tomato Suggests Negative Regulation of Plant Basal Defense and Systemic Acquired Resistance by AtbZIP62 Transcription Factor. Int. J. Mol. Sci. 2021, 22, 11541. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.P.; Coacker, G. Genome sequences of two Pseudomonas syringae pv. tomato race 1 strains, isolated from tomato fields in California. Genome Announc. 2016, 4, e01671-15. [Google Scholar] [CrossRef]
- Nicoletti, R.; Petriccione, M.; Curci, M.; Scortichini, M. Hazelnut-Associated Bacteria and Their Implications in Crop Management. Horticulturae 2022, 8, 1195. [Google Scholar] [CrossRef]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef]
- Martinez, C.; Blanc, F.; Le Claire, E.; Besnard, O.; Nicole, M.; Baccou, J.-C. Salicylic Acid and Ethylene Pathways Are Differentially Activated in Melon Cotyledons by Active or Heat-Denatured Cellulase from Trichoderma longibrachiatum. Plant Physiol. 2001, 127, 334–344. [Google Scholar] [CrossRef]
- Nguyen, Q.-M.; Iswanto, A.B.B.; Son, G.H.; Kim, S.H. Recent Advances in Effector-Triggered Immunity in Plants: New Pieces in the Puzzle Create a Different Paradigm. Int. J. Mol. Sci. 2021, 22, 4709. [Google Scholar] [CrossRef]
- Wang, S.; Wu, H.; Qiao, J.; Ma, L.; Liu, J.; Xia, Y.; Gao, X. Molecular mechanism of plant growth promotion and induced systemic resistance to Tobacco mosaic virus by Bacillus spp. J. Microbiol. Biotechnol. 2009, 19, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Sharma, M.; Saleh, D.; Charron, J.-B.; Jabaji, S. A Crosstalk Between Brachypodium Root Exudates, Organic Acids, and Bacillus velezensis B26, a Growth Promoting Bacterium. Front. Microbiol. 2020, 11, 575578. [Google Scholar] [CrossRef]
- Grimm, A.C.; Harwood, C.S. Chemotaxis of Pseudomonas spp. to the polyaromatic hydrocarbon naphthalene. Appl. Environ. Microbiol. 1997, 63, 4111–4115. [Google Scholar] [CrossRef] [PubMed]
- Adler, J. A Method for Measuring Chemotaxis and Use of the Method to Determine Optimum Conditions for Chemotaxis by Escherichia coli. J. Gen. Microbiol. 1973, 74, 77–91. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Khedr, A.A.; Mehiar, F.; El-Kady, E.M.; Baazeem, A.; Shimizu, M. Suppression of Pseudomonas syringae pv. tomato infection by rhizosphere fungi. Pest Manag. Sci. 2021, 77, 4350–4356. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Shivanna, M.B.; Meera, M.S.; Hyakumachi, M. Mechanism of induced systemic resistance against anthracnose disease in cucumber by plant growth-promoting fungi. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2015, 65, 287–299. [Google Scholar] [CrossRef]
- Sanchez-Ferrer, A.; Bru, R.; Cabanes, J.; Garcia-Carmona, F. Characterization of catacholase and cresolase activities of Monastrell grape polyphenol oxidase. Phytochemistry 1988, 27, 319–321. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Yaryura, P.M.; León, M.; Correa, O.S.; Kerber, N.L.; Pucheu, N.L.; García, A.F. Assessment of the Role of Chemotaxis and Biofilm Formation as Requirements for Colonization of Roots and Seeds of Soybean Plants by Bacillus amyloliquefaciens BNM339. Curr. Microbiol. 2008, 56, 625–632. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced Systemic Resistance and Promotion of Plant Growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Viswanathan, R.; Raguchander, T.; Prakasam, V.; Samiyappan, R. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop. Prot. 2001, 20, 1–11. [Google Scholar] [CrossRef]
- Vale, F.X.R.D.; Parlevliet, J.E.; Zambolim, L. Concepts in plant disease resistance. Fitopatol. Bras. 2001, 26, 577–589. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van Wees, S.C.; Hoffland, E. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 1996, 8, 1225–1237. [Google Scholar] [PubMed]
- De Meyer, G.; Audenaert, K.; Höfte, M. Pseudomonas aeruginosa 7NSK2-induced Systemic Resistance in Tobacco Depends on in planta Salicylic Acid Accumulation but is not Associated with PR1a Expression. Eur. J. Plant Pathol. 1999, 105, 513–517. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Size | Accession Number |
|---|---|---|---|---|
| LePR1 | GCCAAGCTATAACTACGCTACCAAC | GCAAGAAATGAACCACCATCC | 139 | DQ159948 |
| LePR2 | GGACACCCTTCCGCTACTCTT | TGTTCCTGCCCCTCCTTTC | 81 | M80604 |
| LeLOX | ATCTCCCAAGTGAAACACCACA | TCATAAACCCTGTCCCATTCTTC | 109 | U13681 |
| LePAL | CTGGGGAAGCTTTTCAGAATC | TGCTGCAAGTTACAAATCCAGAG | 150 | AW035278 |
| LeUBI3 | TCCATCTCGTGCTCCGTCT | GAACCTTTCCAGTGTCATCAACC | 144 | X58253 |
| Strain | Genus Species | Host Plant | Chemotactic Activity | |
|---|---|---|---|---|
| Malic Acid | Citric Acid | |||
| N2 | Pseudomonas brassicacearum | Chinese chive | ++ | ++ |
| N6 | Pseudomonas brassicacearum | Chinese chive | ++ | +++ |
| N12 | Pseudomonas brassicacearum | Chinese chive | ++ | ++ |
| N16 | Pseudomonas brassicacearum | Chinese chive | +++ | +++ |
| N22 | Pseudomonas brassicacearum | Chinese chive | ++ | + |
| N27 | Pseudomonas brassicacearum | Chinese chive | ++ | +++ |
| N32 | Pseudomonas brassicacearum | Chinese chive | +++ | +++ |
| N37 | Pseudomonas delhiensis | Chinese chive | ++ | ++ |
| N39 | Pseudomonas fluorescens | Chinese chive | ++ | ++ |
| N42 | Pseudomonas stutzeri | Chinese chive | ++ | +++ |
| N44 | Pseudomonas plecoglossicida | Chinese chive | ++ | ++ |
| O10 | Pseudomonas vranovensis | Onion | ++ | ++ |
| O26 | Pseudomonas umsongensis | Onion | +++ | +++ |
| O29 | Pseudomonas vranovensis | Onion | ++ | ++ |
| O35 | Burkholderia cepacia | Onion | +++ | +++ |
| O39 | Pseudomonas vranovensis | Onion | ++ | ++ |
| O41 | Pseudomonas fulval | Onion | ++ | ++ |
| A5 | Pseudomonas resinovorans | Garlic | +++ | +++ |
| A6 | Pseudomonas vranovensis | Garlic | ++ | ++ |
| A25 | Pseudomonas vranovensis | Garlic | +++ | +++ |
| A27 | Pseudomonas resinovorans | Garlic | +++ | ++ |
| A28 | Pseudomonas resinovorans | Garlic | +++ | +++ |
| A30 | Pseudomonas vranovensis | Garlic | ++ | +++ |
| A31 | Pseudomonas resinovorans | Garlic | ++ | ++ |
| A33 | Pseudomonas resinovorans | Garlic | +++ | +++ |
| A47 | Pseudomonas resinovorans | Garlic | +++ | +++ |
| C1 | Pseudomonas putida | Cucumber | ++ | ++ |
| C7 | Pseudomonas putida | Cucumber | + | ++ |
| C14 | Pseudomonas putida | Cucumber | +++ | ++ |
| C21 | Pseudomonas putida | Cucumber | +++ | +++ |
| C27 | Pseudomonas putida | Cucumber | +++ | +++ |
| C33 | Pseudomonas putida | Cucumber | ++ | ++ |
| C37 | Pseudomonas sp. | Cucumber | +++ | +++ |
| C40 | Pseudomonas putida | Cucumber | ++ | ++ |
| C46 | Pseudomonas putida | Cucumber | +++ | ++ |
| T1 | Pseudomonas brassicacearum | Tomato | ++ | + |
| T5 | Pseudomonas putida | Tomato | + | + |
| T11 | Pseudomonas putida | Tomato | ++ | + |
| T15 | Pseudomonas putida | Tomato | +++ | +++ |
| T21 | Pseudomonas fluorescens | Tomato | ++ | + |
| T26 | Pseudomonas brassicacearum | Tomato | +++ | ++ |
| T28 | Pseudomonas alcaligenes | Tomato | ++ | ++ |
| T31 | Pseudomonas putida | Tomato | ++ | ++ |
| T36 | Pseudomonas putida | Tomato | +++ | ++ |
| T41 | Pseudomonas putida | Tomato | ++ | ++ |
| B1 | Pseudomonas resinovorans | Bulk soil | +++ | +++ |
| B5 | Pseudomonas alcaligenes | Bulk soil | +++ | − |
| B6 | Pseudomonas pseudoalcaligenes | Bulk soil | + | + |
| B11 | Pseudomonas citronellolis | Bulk soil | +++ | +++ |
| B12 | Pseudomonas resinovorans | Bulk soil | +++ | ++ |
| B13 | Pseudomonas resinovorans | Bulk soil | +++ | ++ |
| B15 | Pseudomonas resinovorans | Bulk soil | ++ | ++ |
| B16 | Pseudomonas alcaligenes | Bulk soil | +++ | − |
| B19 | Pseudomonas resinovorans | Bulk soil | + | ++ |
| B22 | Pseudomonas resinovorans | Bulk soil | + | + |
| B28 | Pseudomonas resinovorans | Bulk soil | +++ | ++ |
| B29 | pseudomonas resinovorans | Bulk soil | +++ | ++ |
| B30 | Pseudomonas aeruginosa | Bulk soil | +++ | +++ |
| B32 | Pseudomonas resinovorans | Bulk soil | ++ | ++ |
| B36 | Pseudomonas resinovorans | Bulk soil | +++ | ++ |
| B39 | Pseudomonas resinovorans | Bulk soil | ++ | ++ |
| B42 | Pseudomonas alcaligenes | Bulk soil | ++ | + |
| B43 | Pseudomonas panipatensis | Bulk soil | ++ | ++ |
| Strain | Genus Species | Host Plant | Chemotactic Activity | |||||
|---|---|---|---|---|---|---|---|---|
| Malic Acid mM | Citric Acid mM | |||||||
| 1 | 10 | 50 | 1 | 10 | 50 | |||
| N6 | Pseudomonas brassicacearum | Chinese chive | +++ | +++ | ++ | +++ | ++ | ++ |
| N16 | Pseudomonas brassicacearum | Chinese chive | + | ++ | ++ | + | ++ | ++ |
| N32 | Pseudomonas brassicacearum | Chinese chive | +++ | ++ | ++ | +++ | +++ | ++ |
| N42 | Pseudomonas stutzeri | Chinese chive | +++ | +++ | +++ | +++ | +++ | ++ |
| O26 | Pseudomonas umsongensis | Onion | +++ | +++ | ++ | +++ | +++ | ++ |
| O35 | Bacillus cepacia | Onion | +++ | +++ | ++ | +++ | +++ | ++ |
| A5 | Pseudomonas resinovorans | Garlic | +++ | +++ | +++ | +++ | +++ | ++ |
| A28 | Pseudomonas resinovorans | Garlic | +++ | +++ | ++ | +++ | +++ | ++ |
| A30 | Pseudomonas vranovensis | Garlic | +++ | +++ | ++ | +++ | +++ | ++ |
| A33 | Pseudomonas resinovorans | Garlic | ++ | ++ | ++ | +++ | ++ | + |
| A47 | Pseudomonas resinovorans | Garlic | +++ | +++ | ++ | +++ | +++ | ++ |
| C21 | Pseudomonas putida | Cucumber | +++ | +++ | ++ | +++ | ++ | ++ |
| C27 | Pseudomonas putida | Cucumber | +++ | +++ | ++ | +++ | +++ | ++ |
| C37 | Pseudomonas sp. | Cucumber | ++ | ++ | ++ | ++ | ++ | ++ |
| T15 | Pseudomonas putida | Tomato | +++ | +++ | ++ | +++ | +++ | ++ |
| B11 | Pseudomonas citronellolis | Bulk soil | ++ | ++ | ++ | ++ | ++ | ++ |
| B30 | Pseudomonas aeruginosa | Bulk soil | +++ | +++ | +++ | +++ | +++ | ++ |
| B5 | Pseudomonas alcaligenes | Bulk soil | +++ | +++ | ++ | − | − | − |
| B16 | Pseudomonas alcaligenes | Bulk soil | +++ | +++ | ++ | − | − | − |
| Treatments | Peroxidase | Polyphenol Oxidase | ||
|---|---|---|---|---|
| 3DPI | 6DPI | 3DPI | 6DPI | |
| O26 | 50.9b | 38.9a | 25.3b | 22.4b |
| A5 | 54.8a | 39.5a | 29.7a | 25.2a |
| A30 | 51.2b | 35.6b | 24.9b | 21.9b |
| A28 | 46.3c | 31.5c | 20.8c | 17.4c |
| A33 | 56.1a | 41.1a | 30.2a | 25.4a |
| A47 | 55.3a | 40.5a | 29.9a | 24.8a |
| N6 | 51.5b | 35.9b | 25.3b | 21.8b |
| N32 | 51.3b | 36.1b | 25.4b | 22.2b |
| N42 | 55.1a | 39.8a | 30.5a | 25.6a |
| T15 | 55.9a | 40.7a | 30.1a | 25.3a |
| C21 | 45.9c | 30.4c | 19.9c | 15.7d |
| B30 | 46.1c | 31.2c | 20.6c | 15.4d |
| B5 | 40.4d | 26.3d | 19.8c | 14.9d |
| B16 | 41.9d | 25.9d | 20.3c | 15.3d |
| Control | 19.1e | 18.6e | 8.9d | 8.9e |
| Treatments | Peroxidase | Polyphenol Oxidase | ||
|---|---|---|---|---|
| 3DPI | 6DPI | 3DPI | 6DPI | |
| O26 + BABA | 60.6b* | 44.6b | 35.3c | 32.7c |
| A5 + BABA | 66.7a | 45.3b | 39.9b | 32.5c |
| A30 + BABA | 59.1b | 39.8c | 34.8c | 29.4d |
| A33 + BABA | 65.7a | 45.6b | 35.1c | 30.2d |
| A47 + BABA | 65.9a | 45.1b | 41.2a | 36.3b |
| N6 + BABA | 60.4b | 41.1c | 34.3c | 30.8d |
| N32 + BABA | 67.3a | 49.2a | 41.5a | 37.9b |
| N42 + BABA | 66.1a | 40.9c | 34.3c | 30.5d |
| T15 + BABA | 66.0a | 49.4a | 45.4a | 41.1a |
| BABA | 54.2c | 33.2d | 31.5e | 27.9e |
| Control | 21.3d | 17.2e | 9.5f | 9.1f |
| Treatments | Morphological and Physiological Characters | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | Leaf Area (cm) | No. of Branches | Fresh Weight (g) | Dry Weight (g) | Chlorophyll Contents | No. of Flowers | No. of Fruits | Weight of Fruits/Plant | |
| O26 | 75.0e* | 225.4d | 20.0c | 417.3d | 65.5c | 44.7b | 105.2b | 34.6a | 4.6b |
| A5 | 92.1b | 277.8b | 24.2b | 449.0b | 76.3a | 48.3a | 116.9a | 42.8a | 5.8a |
| A30 | 91.3b | 273.5c | 23.9b | 447.9b | 76.2a | 47.6a | 113.9 | 38.5b | 5.7a |
| A28 | 79.4c | 226.4d | 20.4c | 427.5c | 68.6b | 45.1b | 105.8b | 35.1c | 4.7b |
| A33 | 80.2c | 226.7d | 21.1c | 428.2c | 69.2b | 45.3b | 104.1b | 35.7c | 4.9b |
| A47 | 92.3b | 278.1b | 24.1b | 449.2b | 77.1a | 48.3a | 117. 3a | 42.9a | 5.9a |
| N6 | 91.9b | 276.9b | 23.4b | 448.7b | 76.9a | 48.1a | 117.8a | 42.6a | 5.9a |
| N32 | 62.7f | 192.4e | 16.5d | 337.9e | 52.8b | 39.1c | 99.3c | 23.2b | 4.0c |
| N42 | 96.5a | 281.7a | 26.2a | 452.1a | 78.3a | 48.7a | 117.4a | 43.0a | 6.0a |
| T15 | 97.6a | 281.9a | 25.9a | 451.9a | 78.1a | 49.1a | 118.1a | 43.5a | 6.2a |
| C21 | 92.3b | 277.3a | 25.2a | 451.7a | 76.9a | 47.5a | 117.9a | 42.6a | 5.9a |
| B30 | 78.4d | 226.7d | 20.6c | 428.3c | 68.5b | 45.2b | 104.8b | 34.8c | 4.6b |
| B5 | 63.7f | 193.1e | 17.2d | 339.0e | 53.3d | 38.6c | 97.9c | 29.7d | 4.1c |
| B16 | 62.9f | 194.2e | 17.0d | 338.4e | 52.9d | 39.1c | 98.2c | 29.9d | 4.2c |
| Control | 49.5g | 119.2f | 13.5e | 244.0f | 38.1e | 35.5d | 78.4d | 20.6e | 3.3d |
| Treatments | Data Analysis | ||
|---|---|---|---|
| Acidity | Total Soluble Solids (TSS)% | Vitamin C mg/100 g | |
| O26 | 0.53b* | 4.76b | 13.14b |
| A5 | 0.58a | 5.22a | 13.54ab |
| A30 | 0.57a | 5.19a | 13.49ab |
| A28 | 0.54b | 4.69b | 13.19b |
| A33 | 0.52b | 4.75b | 13.09b |
| A47 | 0.58a | 4.78b | 13.63a |
| N6 | 0.58a | 4.73b | 13.72a |
| N32 | 0.49c | 4.81b | 12.63c |
| N42 | 0.59a | 5.19a | 13.69a |
| T15 | 0.58a | 5.21a | 13.73a |
| C21 | 0.57a | 5.18a | 13.62a |
| B30 | 0.53b | 4.79b | 13.17b |
| B5 | 0.48c | 4.75b | 12.59c |
| B16 | 0.46c | 4.73b | 12.68c |
| Control | 0.47c | 4.69b | 12.02d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsharkawy, M.M.; Khedr, A.A.; Mehiar, F.; El-Kady, E.M.; Alwutayd, K.M.; Behiry, S.I. Rhizobacterial Colonization and Management of Bacterial Speck Pathogen in Tomato by Pseudomonas spp. Microorganisms 2023, 11, 1103. https://doi.org/10.3390/microorganisms11051103
Elsharkawy MM, Khedr AA, Mehiar F, El-Kady EM, Alwutayd KM, Behiry SI. Rhizobacterial Colonization and Management of Bacterial Speck Pathogen in Tomato by Pseudomonas spp. Microorganisms. 2023; 11(5):1103. https://doi.org/10.3390/microorganisms11051103
Chicago/Turabian StyleElsharkawy, Mohsen M., Amr A. Khedr, Farid Mehiar, Elsayed M. El-Kady, Khairiah Mubarak Alwutayd, and Said I. Behiry. 2023. "Rhizobacterial Colonization and Management of Bacterial Speck Pathogen in Tomato by Pseudomonas spp." Microorganisms 11, no. 5: 1103. https://doi.org/10.3390/microorganisms11051103