
Expression and Characterization of 3,6-Dihydroxy-picolinic Acid Decarboxylase PicC of Bordetella bronchiseptica RB50
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Culture Media
2.2. Plasmids and Bacterial Strains
2.3. Cloning the picCRB50 Gene
2.4. Expression and Purification PicCRB50
2.5. Enzyme Assay of Purified His-Tagged PicCRB50
2.6. Analytical Methods
3. Results
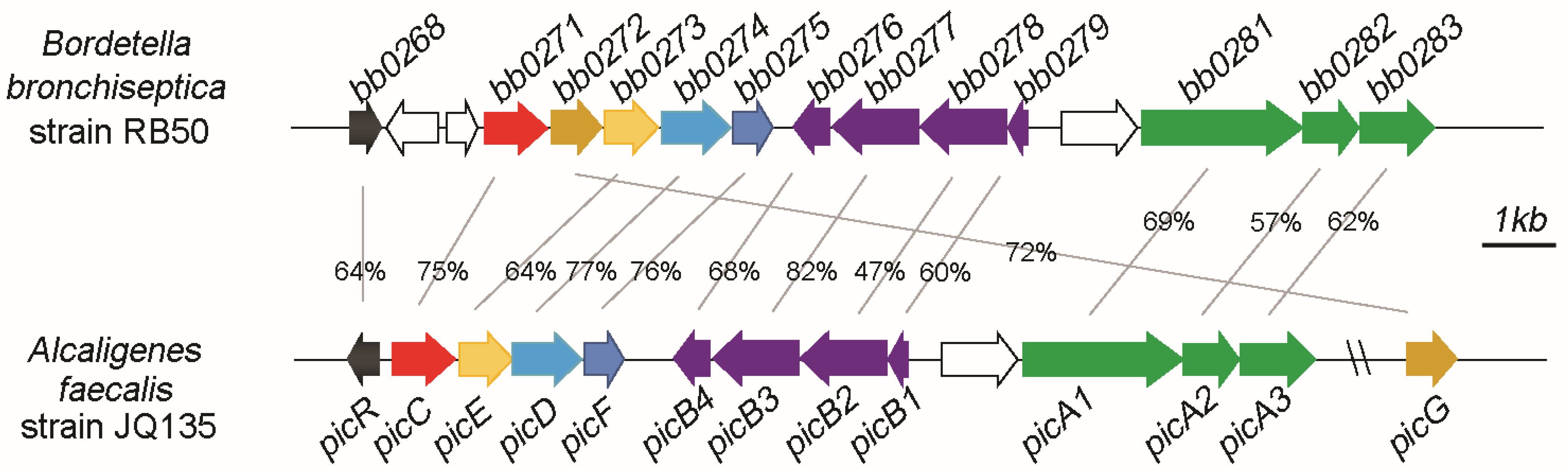
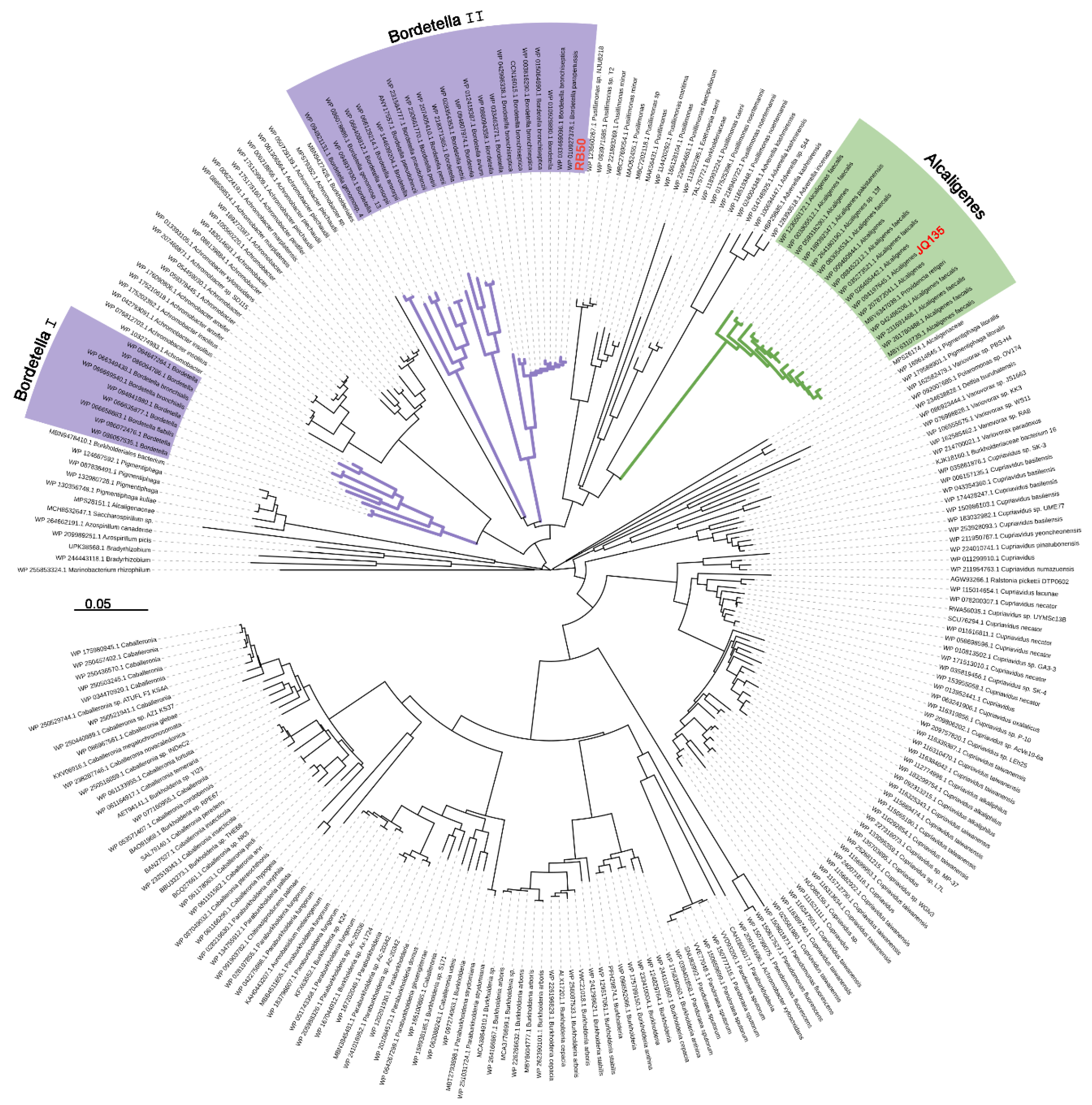
3.1. Pic Gene Cluster Present in Bordetella spp.
3.2. Organization of Pic Gene Cluster of B. bronchiseptica RB50
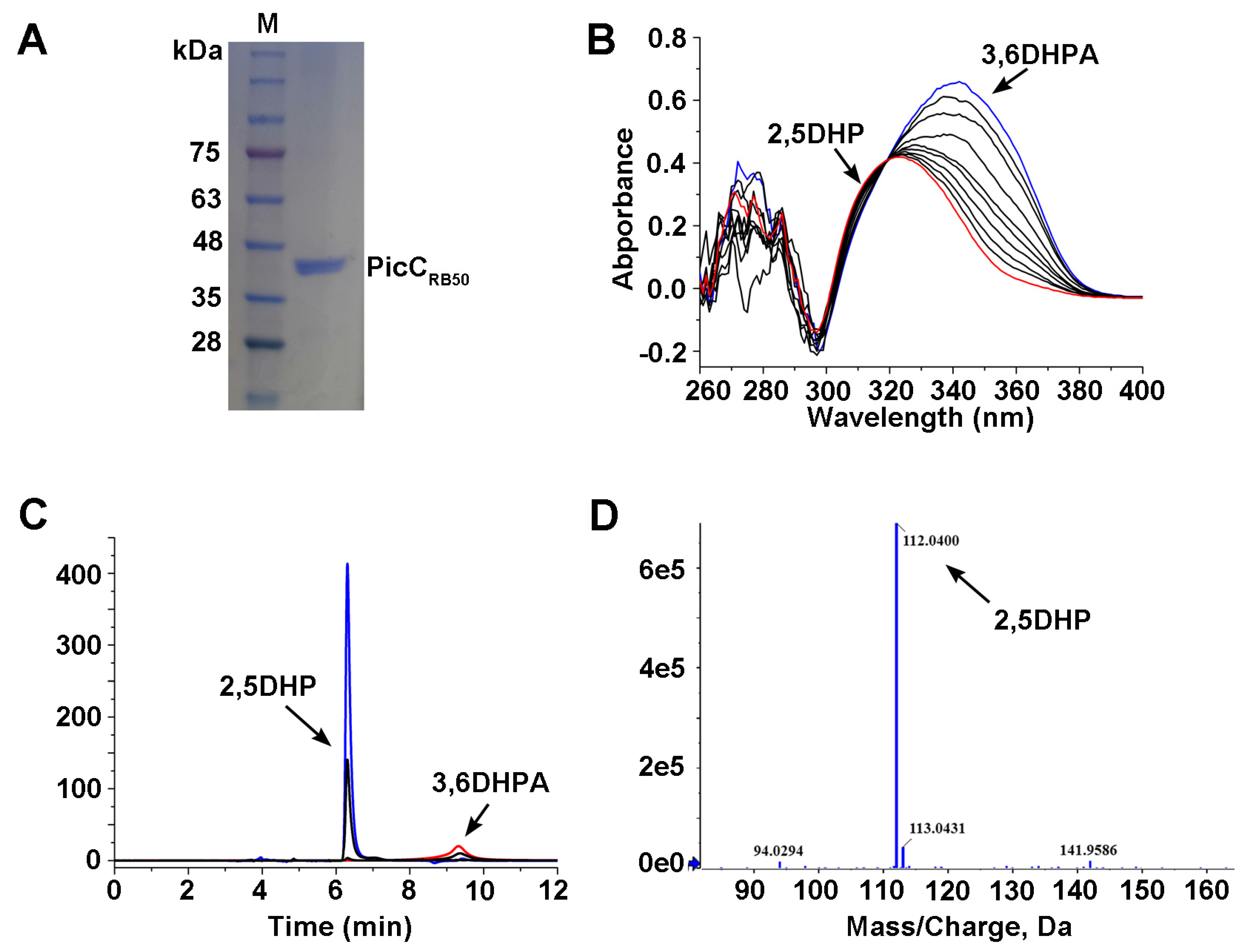
3.3. Cloning and Over-Expression of PicCRB50
3.4. The PicCRB50 Catalyzes 3,6DHPA into 2.5DHP
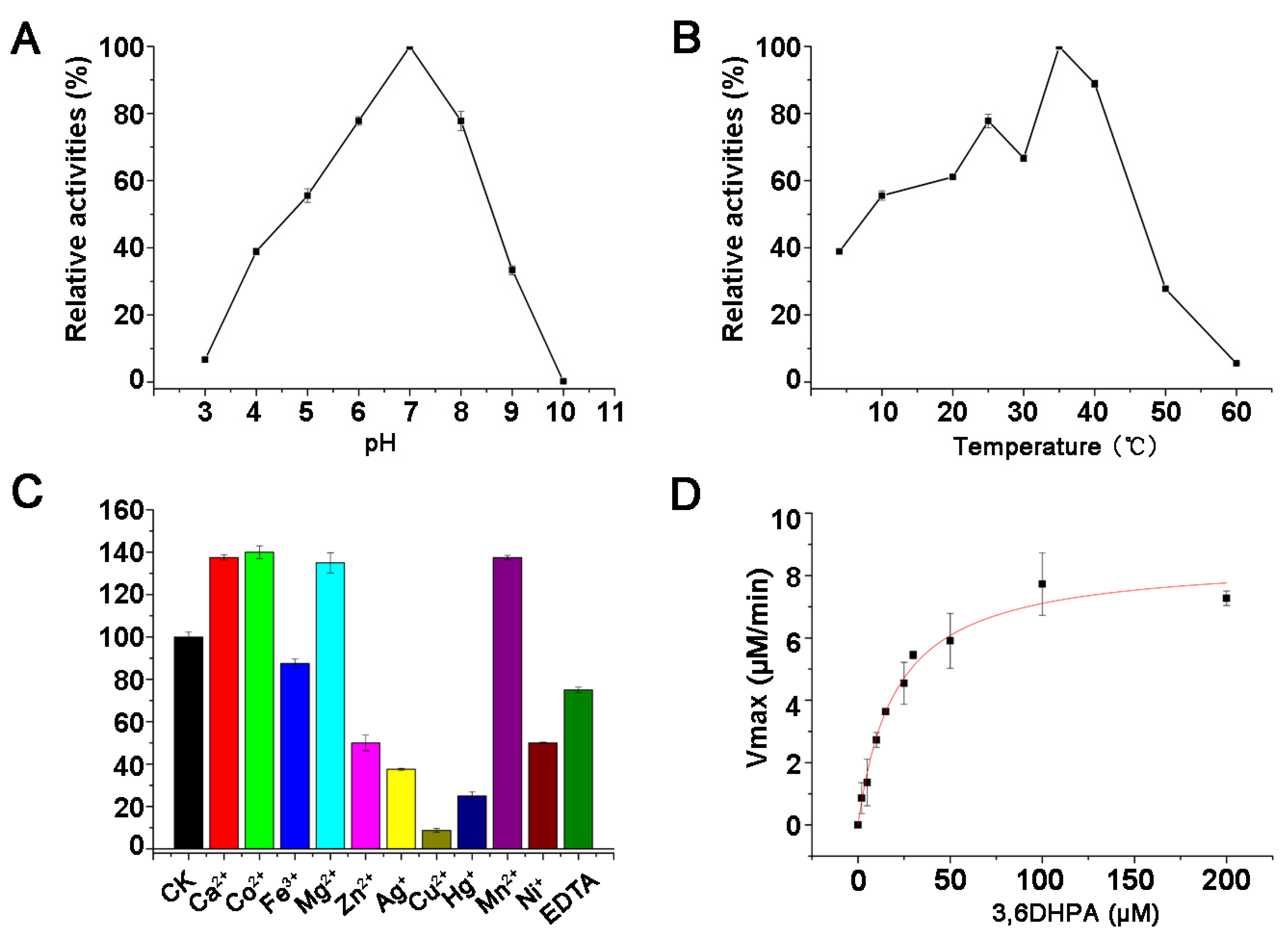
3.5. Characteristics of the PicCRB50
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaiser, J.-P.; Feng, Y.; Bollag, J.-M. Microbial metabolism of pyridine, quinoline, acridine, and their derivatives under aerobic and anaerobic conditions. Microbiol. Rev. 1996, 60, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Puripattanavong, J.; Songkram, C.; Lomlim, L.; Amnuaikit, T. Development of concentrated emulsion containing Nicotiana tabacum extract for use as pesticide. J. Appl. Pharmaceut. Sci. 2013, 3, 016–021. [Google Scholar]
- Časaitė, V.; Stanislauskienė, R.; Vaitekūnas, J.; Tauraitė, D.; Rutkienė, R.; Gasparavičiūtė, R.; Meškys, R. Microbial degradation of pyridine: A complete pathway in Arthrobacter sp. strain 68b deciphered. Appl. Environ. Microbiol. 2020, 86, e00902–e00920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Xiong, J.; Zhao, Z.; Chai, T. The enhancement of pyridine degradation by RhodococcusKDPy1 in coking wastewater. FEMS Microbiol. Lett. 2018, 366, fny271. [Google Scholar] [CrossRef]
- Guadie, A.; Han, J.-L.; Liu, W.; Ding, Y.-C.; Minale, M.; Ajibade, F.O.; Zhai, S.; Wang, H.-C.; Cheng, H.; Ren, N.; et al. Evaluating the effect of fenton pretreated pyridine wastewater under different biological conditions: Microbial diversity and biotransformation pathways. J. Environ. Manag. 2021, 287, 112297. [Google Scholar] [CrossRef]
- Nishino, S.F.; Spain, J.C. Degradation of nitrobenzene by a Pseudomonas pseudoalcaligenes. Appl. Environ. Microbiol. 1993, 59, 2520–2525. [Google Scholar] [CrossRef] [Green Version]
- Bryleva, E.Y.; Brundin, L. Kynurenine pathway metabolites and suicidality. Neuropharmacology 2017, 112, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Chirino, B.; Strahsburger, E.; Agulló, L.; González, M.; Seeger, M. Genomic and Functional Analyses of the 2-Aminophenol Catabolic Pathway and Partial Conversion of Its Substrate into Picolinic Acid in Burkholderia xenovorans LB400. PLOS ONE 2013, 8, e75746. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Yasuda, M.; Yamasaki, K. Stability Constants of Picolinic and Quinaldic Acid Chelates of Bivalent Metals. J. Phys. Chem. 1957, 61, 229–231. [Google Scholar] [CrossRef]
- Donahue, C.J.; Archer, R.D. Transition metal eight-coordination. 8. Stereochemical integrity, geometrical isomers, and isomerization of mixed ligand tungsten (IV) chelates containing picolinic acid and 8-quinolinol derivatives. J. Am. Chem. Soc. 1977, 99, 6613–6623. [Google Scholar] [CrossRef]
- Yousef, T.; El-Reash, G.A.; Al-Zahab, M.A.; Safaan, M. Physicochemical investigations, biological studies of the Cr (III), Mn (II), Fe (III), Co (II), Ni (II), Cu (II), Zn (II), Cd (II), Hg (II) and UO2 (VI) complexes of picolinic acid hydrazide derivative: A combined experimental and computational approach. J. Mol. Struct. 2019, 1197, 564–575. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, Y.; Yao, S.; Ren, H.; Qian, M.; Hong, Q.; Lu, Z.; He, J. Novel 3,6-Dihydroxypicolinic Acid Decarboxylase-Mediated Picolinic Acid Catabolism in Alcaligenes faecalis JQ135. J. Bacteriol. 2019, 201, e00665-18. [Google Scholar] [CrossRef]
- Qiu, J.; Zhao, L.; Xu, S.; Chen, Q.; Chen, L.; Liu, B.; Hong, Q.; Lu, Z.; He, J. Identification and Characterization of a Novel pic Gene Cluster Responsible for Picolinic Acid Degradation in Alcaligenes faecalis JQ135. J. Bacteriol. 2019, 201, e00077-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyes, M.P.; Chen, C.Y.; Major, E.O.; Saito, K. Different kynurenine pathway enzymes limit quinolinic acid formation by various human cell types. Biochem. J. 1997, 326, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Wang, Q.; Ning, Y.; Fan, Y.; Feng, S.; He, C.; Zhang, T.C.; Shen, Z. Isolation of a 2-picolinic acid-assimilating bacterium and its proposed degradation pathway. Bioresour. Technol. 2017, 245, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Zhou, J.; Wang, J.; Qu, B.; Lu, H.; Zhao, H. Aerobic degradation of 2-picolinic acid by a nitrobenzene-assimilating strain: Streptomyces sp. Z2. Bioresour. Technol. 2009, 100, 2082–2084. [Google Scholar] [CrossRef]
- Tate, R.L.; Ensign, J.C. Picolinic acid hydroxylase of Arthrobacter picolinophilus. Can. J. Microbiol. 1974, 20, 695–702. [Google Scholar] [CrossRef]
- Xu, S.; Wang, X.; Zhang, F.; Jiang, Y.; Zhang, Y.; Cheng, M.; Yan, X.; Hong, Q.; He, J.; Qiu, J. PicR as a MarR Family Transcriptional Repressor Multiply Controls the Transcription of Picolinic Acid Degradation Gene Cluster pic in Alcaligenes faecalis JQ135. Appl. Environ. Microbiol. 2022, 88, e00172-22. [Google Scholar] [CrossRef]
- Parkhill, J.; Sebaihia, M.; Preston, A.; Murphy, L.D.; Thomson, N.; Harris, D.E.; Holden, M.T.; Churcher, C.M.; Bentley, S.D.; Mungall, K.L. Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat. Genet. 2003, 35, 32. [Google Scholar] [CrossRef]
- Linz, B.; Ma, L.; Rivera, I.; Harvill, E.T. Genotypic and phenotypic adaptation of pathogens: Lesson from the genus Bordetella. Current. Opinion Infect. Diseas. 2019, 32, 223. [Google Scholar] [CrossRef]
- Bridel, S.; Bouchez, V.; Brancotte, B.; Hauck, S.; Armatys, N.; Landier, A.; Mühle, E.; Guillot, S.; Toubiana, J.; Maiden, M.C.J.; et al. A comprehensive resource for Bordetella genomic epidemiology and biodiversity studies. Nat. Commun. 2022, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nash, Z.M.; Cotter, P.A. Regulated, sequential processing by multiple proteases is required for proper maturation and release of Bordetella filamentous hemagglutinin. Mol. Microbiol. 2019, 112, 820–836. [Google Scholar] [CrossRef]
- Bone, M.A.; Wilk, A.J.; Perault, A.I.; Marlatt, S.A.; Scheller, E.V.; Anthouard, R.; Chen, Q.; Stibitz, S.; Cotter, P.A.; Julio, S.M. Bordetella PlrSR regulatory system controls BvgAS activity and virulence in the lower respiratory tract. Proc. Natl. Acad. Sci. USA 2017, 114, E1519–E1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brickman, T.J.; Armstrong, S.K. The Bordetella bronchiseptica nic locus encodes a nicotinic acid degradation pathway and the 6-hydroxynicotinate-responsive regulator BpsR. Mol. Microbiol. 2018, 108, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jiang, W.; Wang, X.; Cheng, T.; Zhang, D.; Wang, H.; Qiu, J.; Cao, L.; Wang, X.; Hong, Q. An amidase gene ipaH is responsible for the initial degradation step of iprodione in strain Paenarthrobacter sp. YJN-5. Appl. Environ. Microbiol. 2018, 84, 01150-18. [Google Scholar] [CrossRef] [Green Version]
- Gross, R.; Guzman, C.A.; Sebaihia, M.; Santos, V.A.P.M.D.; Pieper, D.H.; Koebnik, R.; Lechner, M.; Bartels, D.; Buhrmester, J.; Choudhuri, J.V.; et al. The missing link: Bordetella petrii is endowed with both the metabolic versatility of environmental bacteria and virulence traits of pathogenic Bordetellae. BMC Genom. 2008, 9, 449. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Yao, Y.; Wang, L.; Yu, H.; Ren, Y.; Wu, G.; Xu, P. Genomic analysis of Pseudomonas putida: Genes in a genome island are crucial for nicotine degradation. Sci. Rep. 2012, 2, 377. [Google Scholar] [CrossRef] [Green Version]
- Jhamandas, K.; Boegman, R.; Beninger, R.; Bialik, M. Quinolinate-induced cortical cholinergic damage: Modulation by tryptophan metabolites. Brain Res. 1990, 529, 185–191. [Google Scholar] [CrossRef]
- Nakata, H.M.; Halvorson, H.O. Biochemical changes occurring during growth and sporulation of bacillus cereus. J. Bacteriol. 1960, 80, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Lowe, S.E.; Jain, M.K.; Zeikus, J.G. Biology, ecology, and biotechnological applications of anaerobic bacteria adapted to environmental stresses in temperature, pH, salinity, or substrates. Microbiol. Rev. 1993, 57, 451–509. [Google Scholar] [CrossRef] [PubMed]
- Irving, S.E.; Choudhury, N.R.; Corrigan, R.M. The stringent response and physiological roles of (pp) pGpp in bacteria. Nat. Rev. Microbiol. 2021, 19, 256–271. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, C.; Zhao, L.; Tong, L.; Wang, L.; Ke, Z.; Yang, Y.; He, J. Expression and Characterization of 3,6-Dihydroxy-picolinic Acid Decarboxylase PicC of Bordetella bronchiseptica RB50. Microorganisms 2023, 11, 854. https://doi.org/10.3390/microorganisms11040854
Yuan C, Zhao L, Tong L, Wang L, Ke Z, Yang Y, He J. Expression and Characterization of 3,6-Dihydroxy-picolinic Acid Decarboxylase PicC of Bordetella bronchiseptica RB50. Microorganisms. 2023; 11(4):854. https://doi.org/10.3390/microorganisms11040854
Chicago/Turabian StyleYuan, Cansheng, Lingling Zhao, Lu Tong, Lin Wang, Zhuang Ke, Ying Yang, and Jian He. 2023. "Expression and Characterization of 3,6-Dihydroxy-picolinic Acid Decarboxylase PicC of Bordetella bronchiseptica RB50" Microorganisms 11, no. 4: 854. https://doi.org/10.3390/microorganisms11040854