
Screening and Identification of Goat-Milk-Derived Lactic Acid Bacteria with Bacteriocin-like Activity and Probiotic Potentials

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. LAB Screening
2.3. Determination of Antimicrobial Activity
2.4. Determination of Bacteriocin-like Substance Stability
2.5. Genomic DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and Strain Identification
2.6. Identification of Antibacterial Properties
2.7. Probiotics Properties Analysis
2.7.1. Hemolytic Test
2.7.2. Determination of Antibiotic Tolerance
2.7.3. Determination of Sugar Metabolism Capacity
2.7.4. Determination of Tolerance to Bile Salts
2.7.5. Determination of Tolerance to Simulate Gastrointestinal Fluids
2.7.6. Analysis of Bacterial Self-Aggregation and Co-Aggregation
2.7.7. Determination of β-Galactosidase Activity
2.8. Animal Experiment
2.9. Tissue Section Analysis
2.10. Data Analysis
3. Results and Discussion
3.1. Screening and Identification of LAB with Antimicrobial Activity
3.2. Determination of Antimicrobial Stability of Bacteriocin-Producing LAB Stains
3.3. Rep-PCR
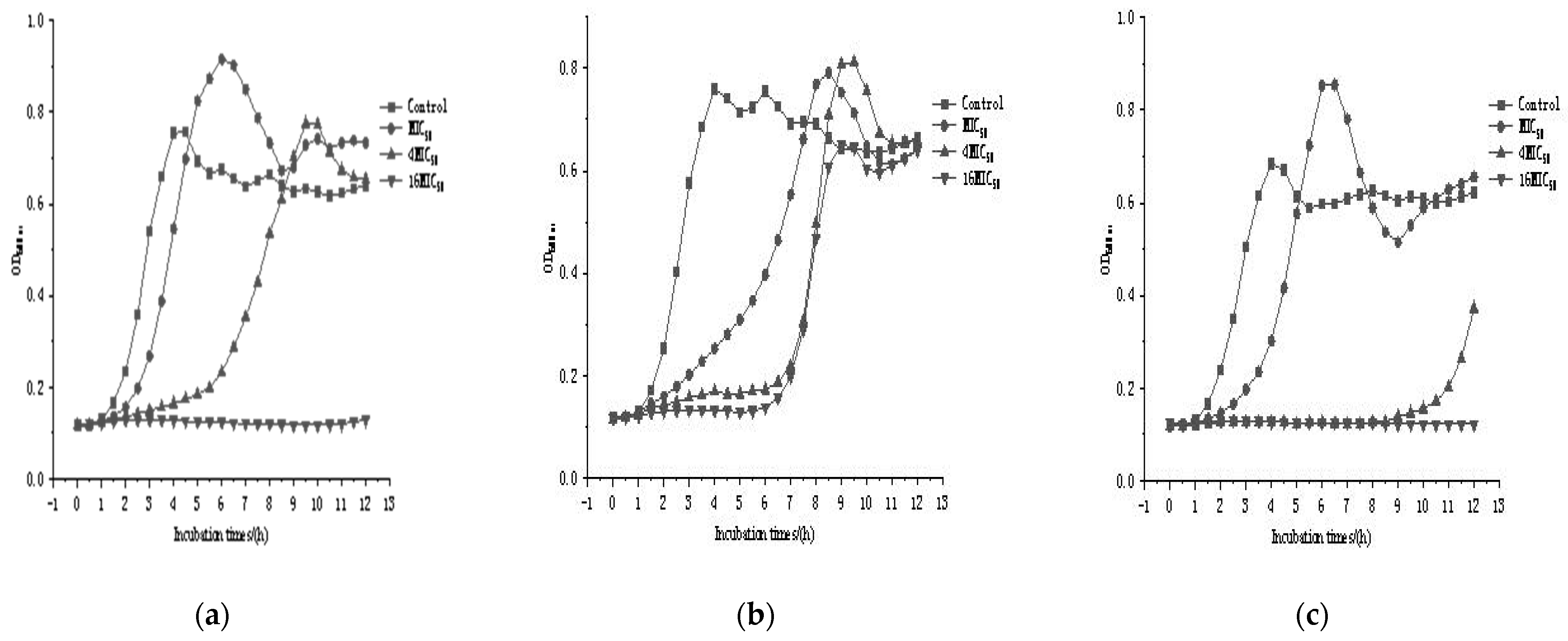
3.4. Fermentation Profiles of the Three Bacteriocin-Producing Strains
3.5. Bacteriostatic Properties of the Three Bacteriocin-Producing Strains
3.6. Characterization of the Probiotic Effects of the Three Bacteriocin-Producing Strains
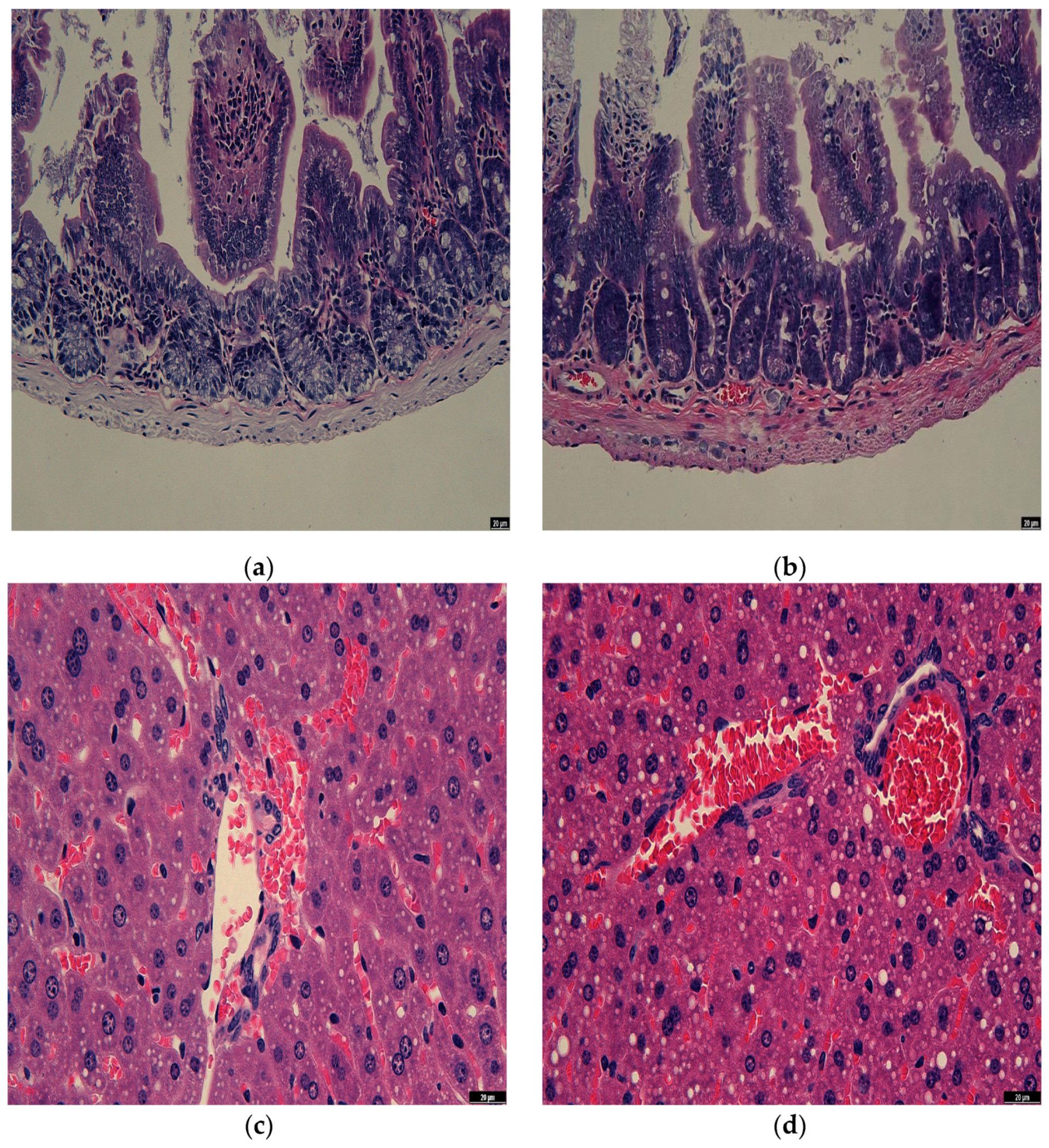
3.7. Observation of Rat Intestine Section
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Herich, R.; Levkut, M. Lactic acid bacteria, probiotics and immune system. Vet. Med. Czech 2002, 47, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Bhat, M.I.; Sharma, D.; Kapila, S.; Kapila, R. Adherence capability and safety assessment of an indigenous probiotic strain Lactobacillus rhamnosus MTCC-5897. Microb. Pathog. 2019, 130, 120–130. [Google Scholar] [CrossRef]
- Capucchio, M.T. Effects of Dietary Supplementation of Lactobacillus acidophilus on Blood Parameters and Gut Health of Rabbits. Animals 2022, 12, 3543. [Google Scholar] [CrossRef]
- Chávez-Martínez, A. Culture Age, Growth Medium, Ultrasound Amplitude, and Time of Exposure Influence the Kinetic Growth of Lactobacillus acidophilus. Fermentation 2023, 9, 63. [Google Scholar] [CrossRef]
- Adel, M.; Safari, R.; Lazado, C.; Caipang, C.; Dadar, M. Host-derived probiotics Enterococcus casseliflavus improves resistance against Streptococcus iniae infection in rainbow trout (Oncorhynchus mykiss) via immunomodulation. Fish Shellfish Immunol. 2016, 52, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Yeung, T.W.; Arroyo-Maya, I.J.; McClements, D.J.; Sela, D.A. Microencapsulation of probiotics in hydrogel particles: Enhancing Lactococcus lactis subsp. cremoris LM0230 viability using calcium alginate beads. Food Funct. 2016, 7, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.P.; Chanyi, R.M.; Schultz, M. Common Organisms and Probiotics: Streptococcus thermophilus (Streptococcus salivarius subsp. thermophilus). In The Microbiota in Gastrointestinal Pathophysiology; Academic Press: Cambridge, MA, USA, 2017; pp. 165–169. [Google Scholar] [CrossRef]
- Settanni, L.; Massitti, O.; Sinderen, D.V.; Corsetti, A. In situ activity of a bacteriocin-producing Lactococcus lactis strain. Influence on the interactions between lactic acid bacteria during sourdough fermentation. J. Appl. Microbiol. 2010, 99, 670–681. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, Q.; Lu, F.; Bie, X.; Zhao, H.; Lu, Z.; Lu, Y. A Novel Class IIb Bacteriocin-Plantaricin EmF Effectively Inhibits Listeria monocytogenes and Extends the Shelf Life of Beef in Combination with Chitosan. J. Agric. Food Chem. 2022, 70, 2187–2196. [Google Scholar] [CrossRef]
- Delves-Broughton, J.P.; Blackburn, P.; Evans, R.J.; Hugenholtz, J. Application of the bacteriocin, nisin. Antonie Leeuwenhoek 1996, 69, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, C.S.; Evans, C.A.; Baines, S.K.; Balthazar, C.F.; Cruz, A.G.; Esmerino, E.A.; Freitas, M.Q.; Pimentel, T.C.; Wittwer, A.E.; Naumovski, N.; et al. Probiotics in Goat Milk Products: Delivery Capacity and Ability to Improve Sensory Attributes. Compr. Rev. Food Sci. Food Saf. 2019, 18, 867–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yelnetty, A.; Purwadi; Ekawati, T.T. Indigenous Lactic Acid Bacteria Isolated from Spontaneously Fermented Goat Milk as Potential Probiotics. Pak. J. Biol. Sci. 2020, 23, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Multiple bacteriocin production in lactic acid bacteria. J. Biosci. Bioeng. 2022, 134, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, A.R.U.; Jahid, I.K. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Chi, H.; Holo, H. Synergistic Antimicrobial Activity Between the Broad Spectrum Bacteriocin Garvicin KS and Nisin, Farnesol and Polymyxin B Against Gram-Positive and Gram-Negative Bacteria. Curr. Microbiol. 2018, 75, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Albufera, U.; Bhugaloo-Vial, P.; Jaufeerally-Fakim, Y. Molecular characterization of Salmonella isolates by REP-PCR and RAPD analysis. Infect. Genet. Evol. 2009, 9, 322–327. [Google Scholar] [CrossRef]
- Ruiz, M.; Silva, P.G.; Laciar, L.A. Comparison of microplate, agar drop and well diffusion plate methods for evaluating hemolytic activity of Listeria monocytogenes. Afr. J. Microbiol. Res. 2009, 3, 319–324. [Google Scholar] [CrossRef]
- Apexa, J.; Neekita, C. Determination of the Probiotics Properties of the Lactic Acid Bacteria Isolated from the Different Homemade Dairy Products. Adv. Life Sci. 2018, 7, 99–111. [Google Scholar]
- Sui, Y.; Liu, J.; Wang, Y.; Xiao, Y.; Gao, Y.; Zhu, D. In vitro probiotic characterization of Lactobacillus strains from fermented tangerine vinegar and their cholesterol degradation activity. Food Biosci. 2021, 39, 100843. [Google Scholar] [CrossRef]
- Sadeghipour, A.; Babaheidarian, P. Making Formalin-Fixed, Paraffin Embedded Blocks. Methods Mol. Biol. 2019, 1897, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthi, R.; Srinivash, M.; Mahalingam, P.U.; Malaikozhundan, B.; Suganya, P.; Gurushankar, K. Antimicrobial, anti-biofilm, antioxidant and cytotoxic effects of bacteriocin by Lactococcus lactis strain CH3 isolated from fermented dairy products-An in vitro and in silico approach. Int. J. Biol. Macromol. 2022, 220, 291–306. [Google Scholar] [CrossRef]
- Martín, I.; Rodríguez, A.; Delgado, J.; Córdoba, J.J. Strategies for Biocontrol of Listeria monocytogenes Using Lactic Acid Bacteria and Their Metabolites in Ready-to-Eat Meat- and Dairy-Ripened Products. Foods 2022, 11, 542. [Google Scholar] [CrossRef]
- Farizano, J.V.; Díaz, V.L.; Masias, E.; Baillo, A.A.; Torino, M.I.; Fadda, S.; Vanden, N.L.; Montenegro, M.A.; Saavedra, L.; Minahk, C. Biotechnological use of dairy by-products for the production and microencapsulation of the food preservative enterocin CRL35. FEMS Microbiol. Lett. 2022, 21, 369. [Google Scholar] [CrossRef]
- Kim, J.; Ahn, J.; Ahn, C. Characterization of novel bacteriocin produced by bacteriocinogenic Carnobacterium maltaromaticum isolated from raw milk. Microb. Pathog. 2022, 173, 105872. [Google Scholar] [CrossRef] [PubMed]
- Ben, O.; Smaoui, S. Enterococci: Between Emerging Pathogens and Potential Probiotics. Biomed Res. Int. 2019, 23, 5938210. [Google Scholar] [CrossRef] [Green Version]
- Perin, L.M.; Belviso, S.; Bello, B.D.; Nero, L.A.; Cocolin, L. Technological Properties and Biogenic Amines Production by Bacteriocinogenic Lactococci and Enterococci Strains Isolated from Raw Goat’s Milk. J. Food Prot. 2017, 80, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Holo, H.; Nilssen, O.; Nes, I.F. Lactococcin A, a new bacteriocin from Lactococcus lactis subsp. cremoris: Isolation and characterization of the protein and its gene. J. Bacteriol. 1991, 173, 3879–3887. [Google Scholar] [CrossRef] [Green Version]
- Moll, G.; Ubbink-Kok, T.; Hildeng-Hauge, H.; Nissen-Meyer, J.; Nes, I.F.; Konings, W.N.; Driessen, A.J. Lactococcin G is a potassium ion-conducting, two-component bacteriocin. J. Bacteriol. 1996, 178, 600–605. [Google Scholar] [CrossRef] [Green Version]
- Chakchouk-Mtibaa, A.; Elleuch, L.; Smaoui, S.; Najah, S.; Sellem, I.; Abdelkafi, S.; Mellouli, L. An antilisterial bacteriocin BacFL31 produced by Enterococcus faecium FL31 with a novel structure containing hydroxyproline residues. Anaerobe 2014, 27, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chakchouk-Mtibaa, A.; Elleuch, L.; Smaoui, S.; Najah, S.; Sellem, I.; Mejdoub, H.; Abdelkafi, S.; Mellouli, L. Characterization of the bacteriocin BacJ1 and its effectiveness for the inactivation of Salmonella typhimurium during turkey escalope storage. Food Chem. 2014, 152, 566–572. [Google Scholar] [CrossRef]
- Simonetta, A.C.; Moragues de Velasco, L.G.; Frisón, L.N. Antibacterial activity of enterococci strains against Vibrio cholerae. Lett. Appl. Microbiol. 1997, 24, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Ben, O.; Cremonesi, P.; Morandi, S.; Smaoui, S.; Hani, K.; Ghrairi, T. Safety characterisation and inhibition of fungi and bacteria by a novel multiple enterocin-producing Enterococcus lactis 4CP3 strain. Microb. Pathog. 2018, 118, 32–38. [Google Scholar] [CrossRef]
- Haas, W.; Shepard, B.D.; Gilmore, M.S. Two-component regulator of Enterococcus faecalis cytolysin responds to quorum-sensing autoinduction. Nature 2002, 415, 84–87. [Google Scholar] [CrossRef]
- Arbulu, S.; Lohans, C.T.; van Belkum, M.J.; Cintas, L.M.; Herranz, C.; Vederas, J.C.; Hernández, P.E. Solution Structure of Enterocin HF, an Antilisterial Bacteriocin Produced by Enterococcus faecium M3K31. J. Agric. Food Chem. 2015, 63, 10689–10695. [Google Scholar] [CrossRef] [PubMed]
- Vuyst, L.D.; Leroy, F. Bacteriocins from lactic acid bacteria: Production, purification, and food applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, B.; Samet, A.; Leibner, J.; Sledzinska, A.; Kur, J. Evaluation of a PCR melting profile technique for bacterial strain differentiation. J. Clin. Microbiol. 2006, 44, 2327–2332. [Google Scholar] [CrossRef] [Green Version]
- Nissen-Meyer, J.; Rogne, P.; Oppegard, C.; Haugen, H.; Kristiansen, P. Structure-function relationships of the non-lanthionine-containing peptide (class II) bacteriocins produced by gram-positive bacteria. Curr. Pharm. Biotechnol. 2009, 10, 19–37. [Google Scholar] [CrossRef]
- Kjos, M.; Oppegrd, C.; Diep, D.B.; Nes, I.F.; Veening, J.W.; Nissen-Meyer, J.; Kristensen, T. Sensitivity to the two-peptide bacteriocin lactococcin G is dependent on UppP, an enzyme involved in cell-wall synthesis. Mol. Microbiol. 2014, 92, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Ekmekci, H.; Aslim, B.; Ozturk, S. Characterization of vaginal lactobacilli coaggregation ability with Escherichia coli. Microbiol. Immunol. 2009, 53, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Dogi, C.A.; Galdeano, C.M.; Perdigón, G. Gut immune stimulation by non pathogenic Gram(+) and Gram(-) bacteria. Comparison with a probiotic strain. Cytokine 2008, 41, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Chang, S.S.; Chang, H.Y.; Wu, C.H.; Pan, C.H.; Chang, C.C.; Chan, C.H.; Huang, H.Y. Probiotic supplementation attenuates age-related sarcopenia via the gut-muscle axis in SAMP8 mice. J. Cachexia Sarcopenia Muscle 2022, 13, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, R.; Gundamaraju, R.; Shinde, T.; Perera, A.P.; Basheer, W.; Southam, B.; Gondalia, S.V.; Karpe, A.V.; Beale, D.J.; Tristram, S.; et al. Lactobacillus acidophilus DDS-1 Modulates Intestinal-Specific Microbiota, Short-Chain Fatty Acid and Immunological Profiles in Aging Mice. Nutrients 2019, 11, 1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Samples | Heat Treatment | Enzyme Treatments | |
|---|---|---|---|
| 100 °C, 10 min | Trypsin | Proteinase K | |
| DH9003 | + | - | - |
| DH9011 | + | - | - |
| DH9012 | + | - | - |
| Sugar Components | DH9003 | DH9011 | DH9012 |
|---|---|---|---|
| L-Arabinose | + | - | + |
| D-xylose | - | + | - |
| Sorbitol | - | - | + |
| α-methyl-d-mannoside | + | - | - |
| Amygdalin | - | ± | - |
| Melibiose | + | - | - |
| D-Trehalose anhydrous | + | + | - |
| Properties | DH9003 | DH9011 | DH9012 |
|---|---|---|---|
| Bile salt tolerance | + | + | + |
| Artificial gastric juice (pH 2.5) | + | + | + |
| Artificial gastric juice (pH 3.0) | + | + | + |
| Artificial gastric juice (pH 3.5) | + | + | + |
| Artificial intestinal fluids | + | + | + |
| Self-aggregation (%) | 52.3 | 46.9 | 30.5 |
| Co-aggregation with Listeria monocytogenes (%) | 52.6 | 15.6 | 68.5 |
| Co-aggregation with Escherichia coli (%) | 63.2 | N | 57.6 |
| Hemolysis | γ-hemolysis | γ-hemolysis | γ-hemolysis |
| β-Galactosidase activity (U/104 cell) | 0.678 | 0.611 | 0.596 |
| Ampicillin (50 mg/mL) | S | S | S |
| Streptomycin sulfate (100 mg/mL) | S | S | S |
| Chloramphenicol (2.5 mg/mL) | R | R | R |
| Nalidixic acid (10 mg/100 mL) | R | R | R |
| Furantoin (10 mg/100 mL) | R | R | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Ni, L.; Tian, W.; Chi, H. Screening and Identification of Goat-Milk-Derived Lactic Acid Bacteria with Bacteriocin-like Activity and Probiotic Potentials. Microorganisms 2023, 11, 849. https://doi.org/10.3390/microorganisms11040849
Yang Z, Ni L, Tian W, Chi H. Screening and Identification of Goat-Milk-Derived Lactic Acid Bacteria with Bacteriocin-like Activity and Probiotic Potentials. Microorganisms. 2023; 11(4):849. https://doi.org/10.3390/microorganisms11040849
Chicago/Turabian StyleYang, Zhenzhu, Ling Ni, Wei Tian, and Hai Chi. 2023. "Screening and Identification of Goat-Milk-Derived Lactic Acid Bacteria with Bacteriocin-like Activity and Probiotic Potentials" Microorganisms 11, no. 4: 849. https://doi.org/10.3390/microorganisms11040849