
Bacterial Communities in Lanna Fermented Soybeans from Three Different Ethnolinguistic Groups in Northern Thailand
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermented Soybean Sampling Sites
2.2. Fermented Soybean Samples
2.2.1. Analysis of Nutritional Compositions and Physicochemical Properties of Fermented Soybean Products
2.2.2. Fermented Soybean Sample Preservation
2.3. Bacterial DNA Extraction
2.4. Bacterial DNA Sequencing
2.5. Sequences Processing and Analysis
2.6. Alpha and Beta Diversity Analysis
2.7. Network Analysis of Bacteria in FSB Bacterial Communities
2.8. Functional Prediction Analysis
3. Results
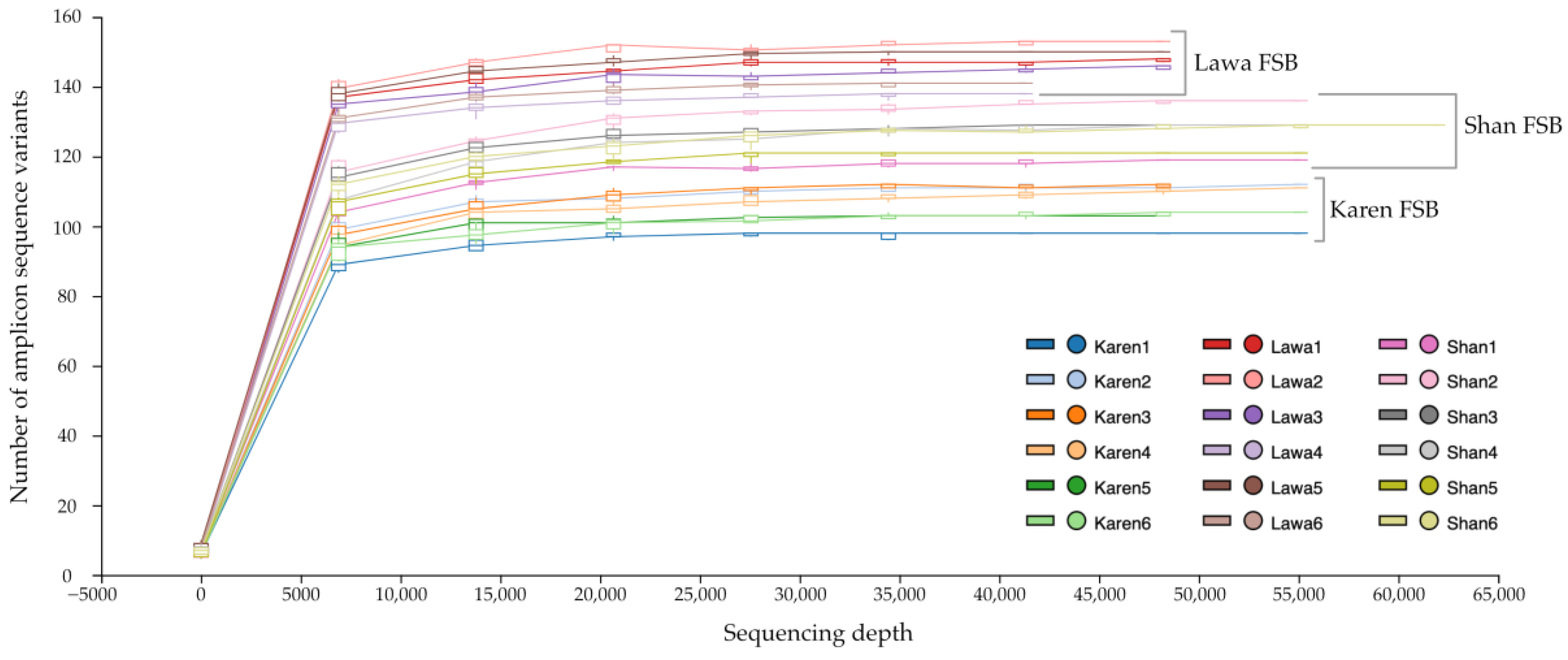
3.1. Number of Sequences Used in Analysis and Rarefaction Analysis
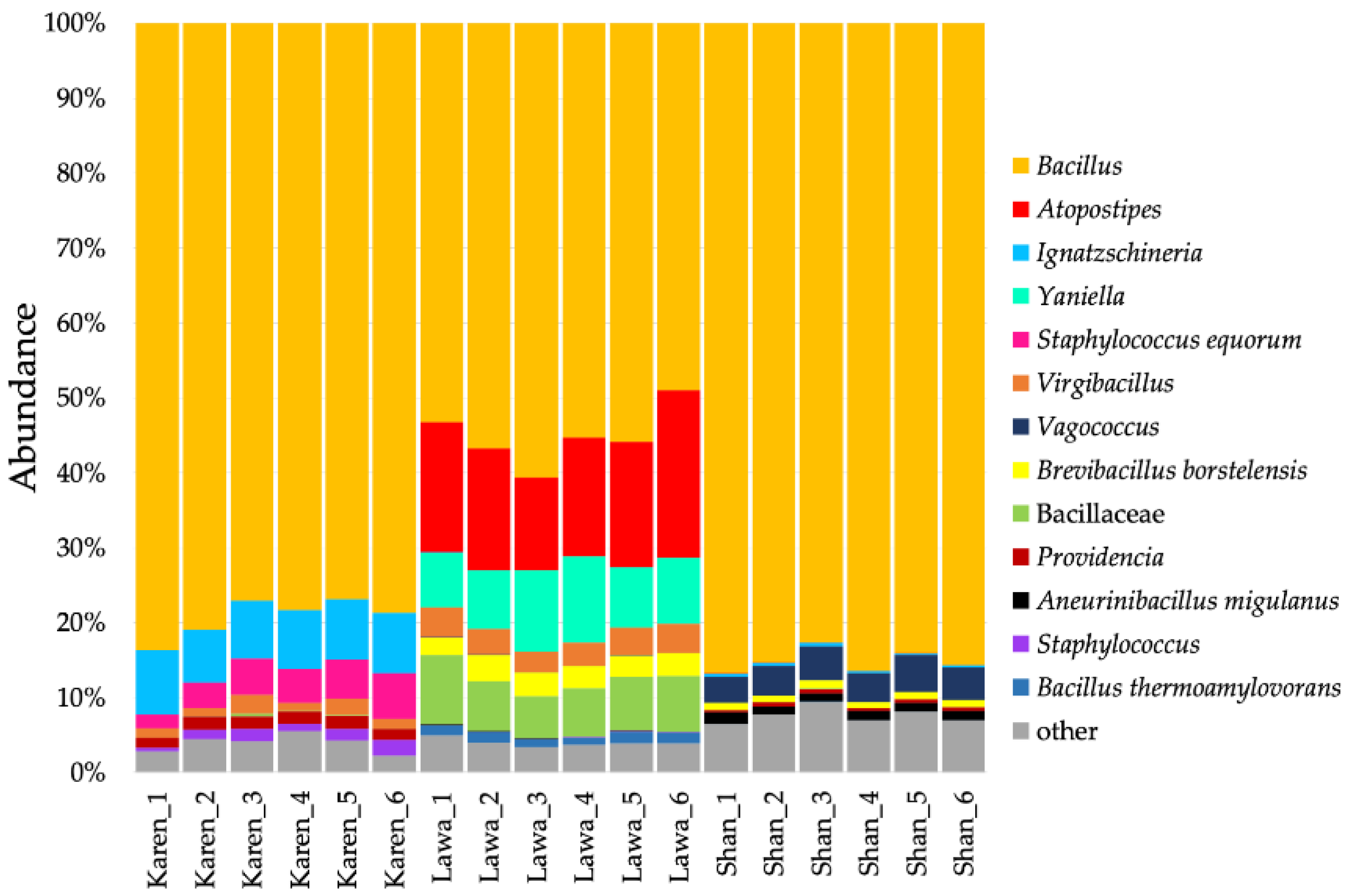
3.2. Bacterial Communities in Fermented Soybeans from Three Ethnic Groups
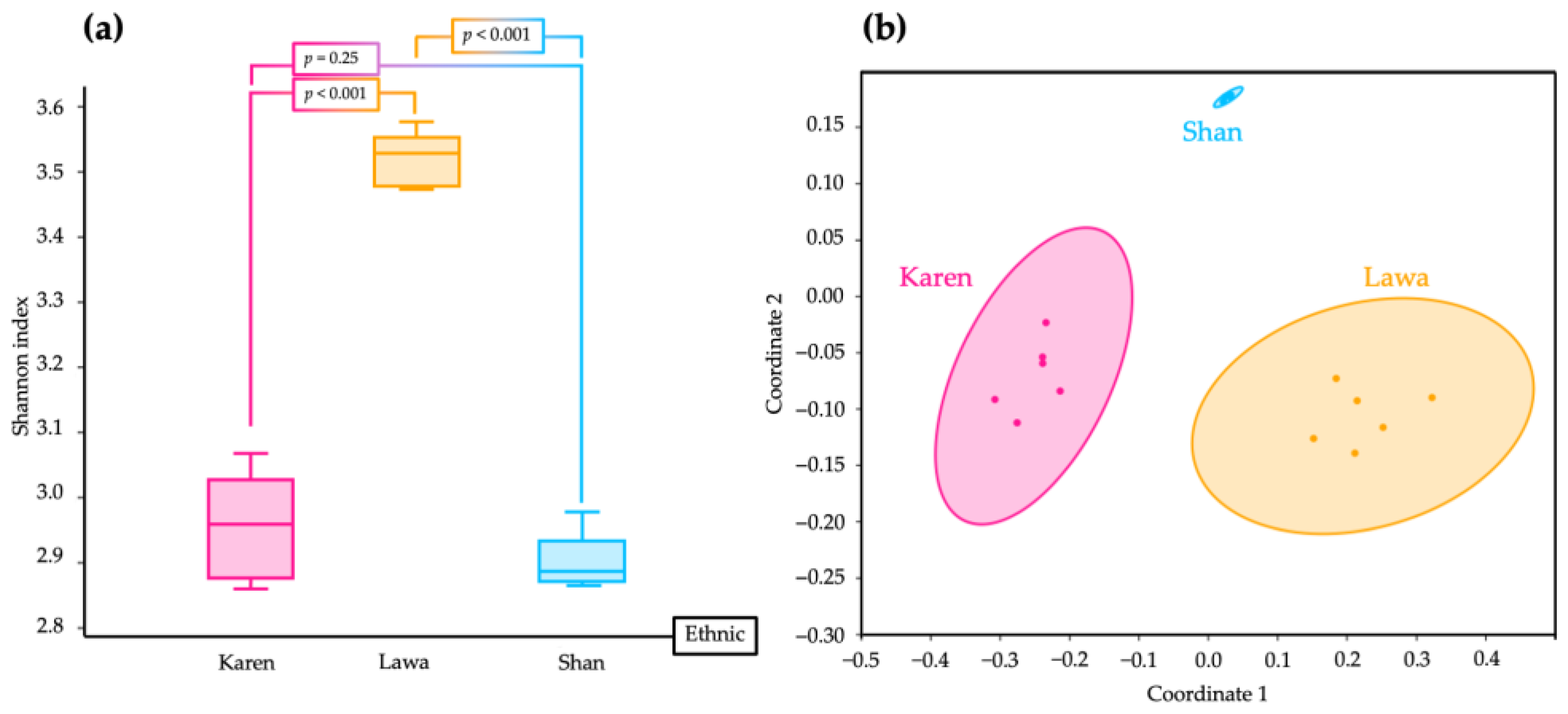
3.3. Alpha and Beta Diversity
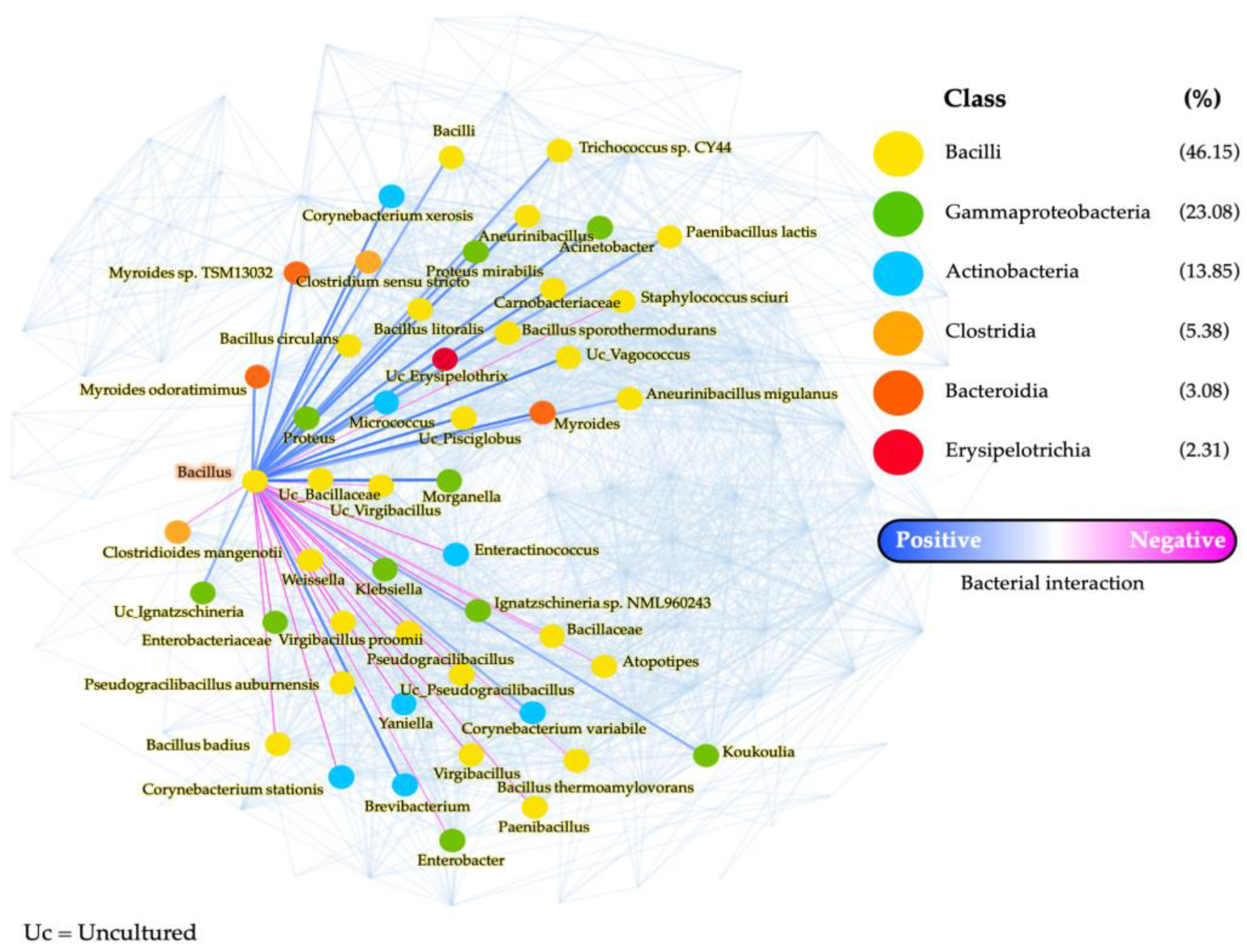
3.4. Network Analysis
3.5. Nutritional and Physicochemical Properties
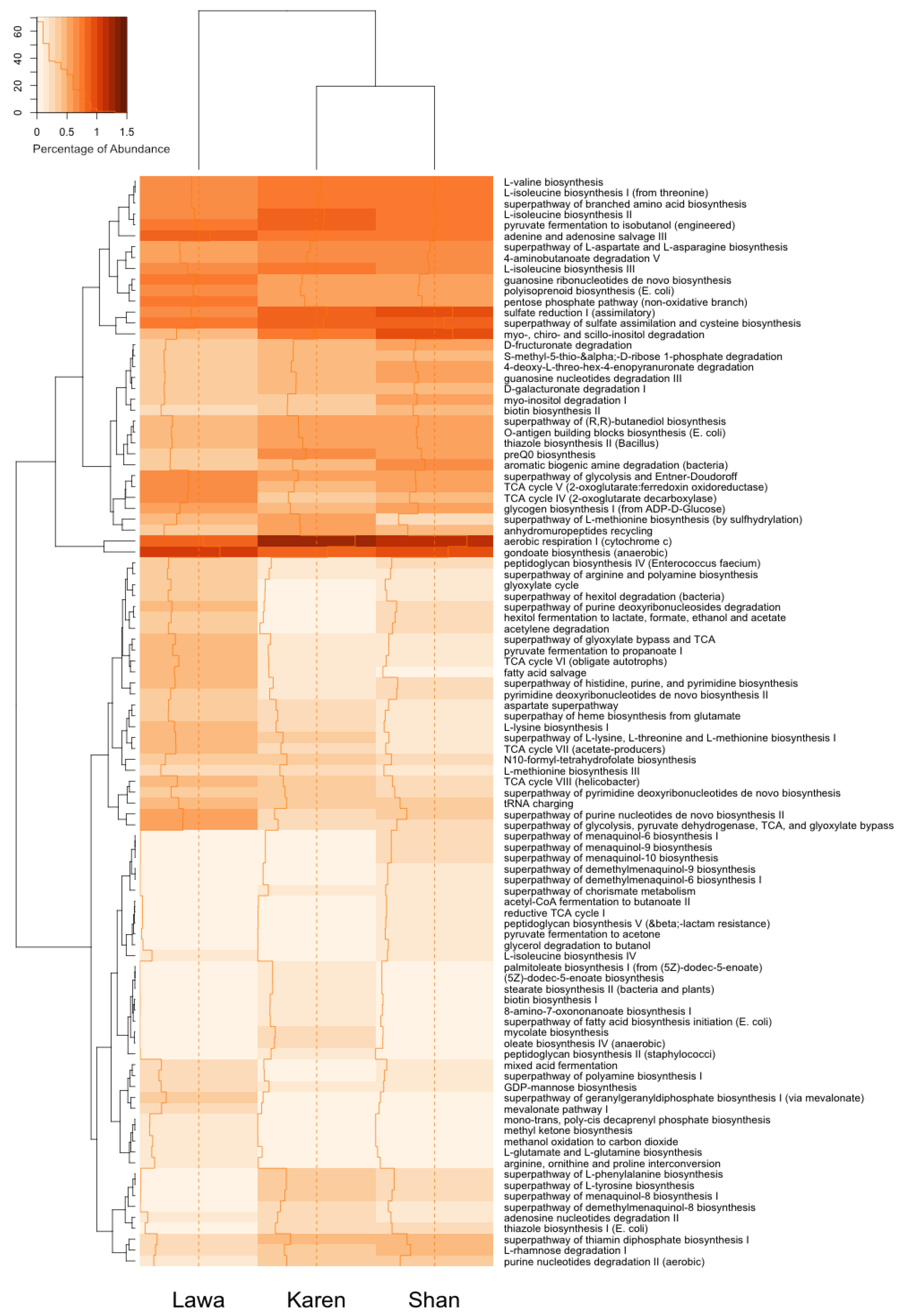
3.6. Functional Gene Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinkraus, K.H. Fermentations in World Food Processing. Compr. Rev. Food Sci. Food Saf. 2002, 1, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Chukeatirote, E. Thua Nao: Thai Fermented Soybean. J. Ethn. Foods 2015, 2, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Tamang, J.P. Himalayan Fermented Foods: Microbiology, Nutrition, and Ethnic Values; CRC Press: Boca Raton, FL, USA, 2009; ISBN 978-1-4200-9325-4. [Google Scholar]
- Kiuchi, K.; Watanabe, S. Industrialization of Japanese Natto. In Food Science and Technology-New York-Marcel Dekker; CRC Press: Boca Raton, FL, USA, 2004; pp. 193–238. ISBN 0-8247-4784-4. [Google Scholar]
- Tamang, J.P. Native Microorganisms in the Fermentation of Kinema. Indian J. Microbiol. 2003, 43, 127–130. [Google Scholar]
- Tamang, J.P.; Thapa, N.; Bhalla, T.C. Savitri Ethnic Fermented Foods and Beverages of India. In Ethnic Fermented Foods and Alcoholic Beverages of Asia; Tamang, J.P., Ed.; Springer: New Delhi, India, 2016; pp. 17–72. ISBN 978-81-322-2800-4. [Google Scholar]
- Kitamura, Y.; Kusumoto, K.-I.; Oguma, T.; Nagai, T.; Furukawa, S.; Suzuki, C.; Satomi, M.; Magariyama, Y.; Takamine, K.; Tamaki, H. Ethnic Fermented Foods and Alcoholic Beverages of Japan. In Ethnic Fermented Foods and Alcoholic Beverages of Asia; Tamang, J.P., Ed.; Springer: New Delhi, India, 2016; pp. 193–236. ISBN 978-81-322-2800-4. [Google Scholar]
- Nam, Y.-D.; Yi, S.-H.; Lim, S.-I. Bacterial Diversity of Cheonggukjang, a Traditional Korean Fermented Food, Analyzed by Barcoded Pyrosequencing. Food Control 2012, 28, 135–142. [Google Scholar] [CrossRef]
- Pakwan, C.; Chitov, T.; Chantawannakul, P.; Manasam, M.; Bovonsombut, S.; Disayathanoowat, T. Bacterial Compositions of Indigenous Lanna (Northern Thai) Fermented Foods and Their Potential Functional Properties. PLoS ONE 2020, 15, e0242560. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Oncharoen, A.; Klanbut, K.; Chukeatirote, E.; Lumyong, S. Characterization of Proteases of Bacillus Subtilis Strain 38 Isolated from Traditionally Fermented Soybean in Northern Thailand. Sci. Asia 2002, 28, 241–245. [Google Scholar] [CrossRef]
- Chettri, R.; Bhutia, M.O.; Tamang, J.P. Poly-γ-Glutamic Acid (PGA)-Producing Bacillus Species Isolated from Kinema, Indian Fermented Soybean Food. Front. Microbiol. 2016, 7, 971. [Google Scholar] [CrossRef]
- Dajanta, K.; Apichartsrangkoon, A.; Chukeatirote, E.; Frazier, R.A. Free-Amino Acid Profiles of Thua Nao, a Thai Fermented Soybean. Food Chem. 2011, 125, 342–347. [Google Scholar] [CrossRef]
- Dajanta, K.; Apichartsrangkoon, A.; Chukeatirote, E. Volatile Profiles of Thua Nao, a Thai Fermented Soy Product. Food Chem. 2011, 125, 464–470. [Google Scholar] [CrossRef]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.-S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen Elimination by Probiotic Bacillus via Signalling Interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Bairoch, A. The ENZYME Database in 2000. Nucleic Acids Res. 2000, 28, 304–305. [Google Scholar] [CrossRef] [Green Version]
- Allaire, J.J. RStudio: Integrated Development Environment for R; RStudio: Boston, MA, USA, 2012; Volume 770, pp. 165–171. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.-H.; Kim, Y.-M.; Park, W.-S.; Kim, J.-H. Ethnic Fermented Foods and Beverages of Korea. In Ethnic Fermented Foods and Alcoholic Beverages of Asia; Tamang, J.P., Ed.; Springer: New Delhi, India, 2016; pp. 263–308. ISBN 978-81-322-2800-4. [Google Scholar]
- Wang, J.; Fung, D.Y.C. Alkaline-Fermented Foods: A Review with Emphasis on Pidan Fermentation. Crit. Rev. Microbiol. 1996, 22, 101–138. [Google Scholar] [CrossRef]
- Parkouda, C.; Nielsen, D.S.; Azokpota, P.; Ivette Irène Ouoba, L.; Amoa-Awua, W.K.; Thorsen, L.; Hounhouigan, J.D.; Jensen, J.S.; Tano-Debrah, K.; Diawara, B.; et al. The Microbiology of Alkaline-Fermentation of Indigenous Seeds Used as Food Condiments in Africa and Asia. Crit. Rev. Microbiol. 2009, 35, 139–156. [Google Scholar] [CrossRef]
- Wang, D.; Liu, Y.; Ngo, H.H.; Zhang, C.; Yang, Q.; Peng, L.; He, D.; Zeng, G.; Li, X.; Ni, B.-J. Approach of Describing Dynamic Production of Volatile Fatty Acids from Sludge Alkaline Fermentation. Bioresour. Technol. 2017, 238, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M. Flavouring Composition Prepared by Fermentation with Bacillus spp. Int. J. Food Microbiol. 2002, 75, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Yokoyama, S. Trends in the Application of Bacillus in Fermented Foods. Curr. Opin. Biotechnol. 2019, 56, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Inatsu, Y.; Nakamura, N.; Yuriko, Y.; Fushimi, T.; Watanasiritum, L.; Kawamoto, S. Characterization of Bacillus Subtilis Strains in Thua Nao, a Traditional Fermented Soybean Food in Northern Thailand. Lett. Appl. Microbiol. 2006, 43, 237–242. [Google Scholar] [CrossRef]
- Muse, H.; Jenkins, R.L.; Oliver, M.B.; Kim, S.; Grantier, R.L.; Malhotra, B.K.; Parham, J.J.; Stover, K.R. A Case of Ignatzschineria indica Bacteremia Following Maggot Colonization. Case Rep. Infect. Dis. 2017, 2017, e3698124. [Google Scholar] [CrossRef] [Green Version]
- Deslandes, V.; Haney, C.; Bernard, K.; Desjardins, M. Ignatzschineria indica Bacteremia in a Patient with a Maggot-Infested Heel Ulcer: A Case Report and Literature Review. Access Microbiol. 2019, 2, acmi000078. [Google Scholar] [CrossRef]
- Reed, K.; Reynolds, S.B.; Smith, C. The First Case of Ignatzschineria Ureiclastica/Larvae in the United States Presenting as a Myiatic Wound Infection. Cureus 2021, 13, e14518. [Google Scholar] [CrossRef]
- Leroy, S.; Lebert, I.; Chacornac, J.-P.; Chavant, P.; Bernardi, T.; Talon, R. Genetic Diversity and Biofilm Formation of Staphylococcus equorum Isolated from Naturally Fermented Sausages and Their Manufacturing Environment. Int. J. Food Microbiol. 2009, 134, 46–51. [Google Scholar] [CrossRef]
- Place, R.B.; Hiestand, D.; Gallmann, H.R.; Teuber, M. Staphylococcus equorum Subsp. Linens, Subsp. Nov., A Starter Culture Component for Surface Ripened Semi-Hard Cheeses. Syst. Appl. Microbiol. 2003, 26, 30–37. [Google Scholar] [CrossRef]
- Jeong, D.-W.; Han, S.; Lee, J.-H. Safety and Technological Characterization of Staphylococcus equorum Isolates from Jeotgal, a Korean High-Salt-Fermented Seafood, for Starter Development. Int. J. Food Microbiol. 2014, 188, 108–115. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Bae, J.-H.; Oh, B.-K.; Hong Lee, W.; Choi, J.-W. Enhancement of Proteolytic Enzyme Activity Excreted from Bacillus Stearothermophilus for a Thermophilic Aerobic Digestion Process. Bioresour. Technol. 2002, 82, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Boominadhan, U.; Rajakumar, R.; Sivakumaar, P.K.V.; Joe, M.M. Optimization of Protease Enzyme Production Using Bacillus sp. Isolated from Different Wastes. Bot. Res. Int. 2009, 2, 83–87. [Google Scholar]
- Xie, J.; Zhang, R.; Shang, C.; Guo, Y. Isolation and Characterization of a Bacteriocin Produced by an Isolated Bacillus Subtilis LFB112 That Exhibits Antimicrobial Activity against Domestic Animal Pathogens. Afr. J. Biotechnol. 2009, 8, 5611–5619. [Google Scholar]
- Zheng, G.; Slavik, M.F. Isolation, Partial Purification and Characterization of a Bacteriocin Produced by a Newly Isolated Bacillus Subtilis Strain. Lett. Appl. Microbiol. 1999, 28, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, T.R.; Lawson, P.A.; Cotta, M.A. Atopostipes. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 1–4. ISBN 978-1-118-96060-8. [Google Scholar]
- Cotta, M.A.; Whitehead, T.R.; Collins, M.D.; Lawson, P.A. Atopostipes Suicloacale Gen. Nov., Sp. Nov., Isolated from an Underground Swine Manure Storage Pit. Anaerobe 2004, 10, 191–195. [Google Scholar] [CrossRef]
- Chen, Y.-G.; Chen, J.; Chen, Q.-H.; Tang, S.-K.; Zhang, Y.-Q.; He, J.-W.; Li, W.-J.; Liu, Y.-Q. Yaniella Soli Sp. Nov., a New Actinobacterium Isolated from Non-Saline Forest Soil in China. Antonie Van Leeuwenhoek 2010, 98, 395–401. [Google Scholar] [CrossRef]
- Schornsteiner, E.; Mann, E.; Bereuter, O.; Wagner, M.; Schmitz-Esser, S. Cultivation-Independent Analysis of Microbial Communities on Austrian Raw Milk Hard Cheese Rinds. Int. J. Food Microbiol. 2014, 180, 88–97. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Button, J.E.; Santarelli, M.; Dutton, R.J. Cheese Rind Communities Provide Tractable Systems for In Situ and In Vitro Studies of Microbial Diversity. Cell 2014, 158, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Tamang, J.P.; Holzapfel, W.H.; Shin, D.H.; Felis, G.E. Editorial: Microbiology of Ethnic Fermented Foods and Alcoholic Beverages of the World. Front. Microbiol. 2017, 8, 1377. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.A.; Devi, K.R.; Ahmed, G.; Jeyaram, K. Microbial and Endogenous Origin of Fibrinolytic Activity in Traditional Fermented Foods of Northeast India. Food Res. Int. 2014, 55, 356–362. [Google Scholar] [CrossRef]
- Schuster, C.; Schmitt, A. Efficacy of a bacterial preparation of Aneurinibacillus migulanus against downy mildew of cucumber (Pseudoperonospora cubensis). Eur. J. Plant Pathol. 2018, 151, 439–450. [Google Scholar] [CrossRef]
- Alenezi, F.N.; Rekik, I.; Bełka, M.; Ibrahim, A.F.; Luptakova, L.; Jaspars, M.; Woodward, S.; Belbahri, L. Strain-Level diversity of secondary metabolism in the biocontrol species Aneurinibacillus migulanus. Microbiol. Res. 2016, 182, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Chandel, S.; Allan, E.J.; Woodward, S. Biological control of Fusarium oxysporum f. sp. lycopersici on tomato by Brevibacillus brevis. J. Phytopathol. 2010, 158, 470–478. [Google Scholar] [CrossRef]
- Nehra, V.; Saharan, B.S.; Choudhary, M. Evaluation of Brevibacillus brevis as a potential plant growth promoting rhizobacteria for cotton (Gossypium hirsutum) Crop. Springerplus 2016, 5, 948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdhi, A.; Kamoun, F.; Messina, C.; Santulli, A.; Bakhrouf, A. Probiotic properties of Brevibacillus brevis and its influence on sea bass (Dicentrarchus labrax) larval rearing. Afr. J. Microbiol. Res. 2012, 6, 6487–6495. [Google Scholar] [CrossRef]
- Choi, M.Y.; Shin, K.-C.; Ho, T.-H.; Park, H.; Nguyen, D.Q.; Park, Y.S.; Kim, D.W.; Oh, D.-K.; Kang, L.-W. Fructuronate-Tagaturonate Epimerase UxaE from Cohnella Laeviribosi Has a Versatile TIM-Barrel Scaffold Suitable for a Sugar Metabolizing Biocatalyst. Int. J. Biol. Macromol. 2020, 163, 1369–1374. [Google Scholar] [CrossRef]
- Rodionova, I.A.; Scott, D.A.; Grishin, N.V.; Osterman, A.L.; Rodionov, D.A. Tagaturonate—Fructuronate Epimerase UxaE, a Novel Enzyme in the Hexuronate Catabolic Network in Thermotoga Maritima. Environ. Microbiol. 2012, 14, 2920–2934. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, e2491. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Composition | Fermented Soybean | ||
|---|---|---|---|
| Karen | Lawa | Shan | |
| Energy (Kcal/100 g) | 310.66 | 291.35 | 315.43 |
| Energy from Fat (Kcal/100 g) | 162.67 | 193.65 | 142.56 |
| Fat (g/100 g) | 7.55 | 8.67 | 6.98 |
| Protein (g/100 g) | 16.33 | 19.23 | 18.86 |
| Carbohydrate (g/100 g) | 7.34 | 9.68 | 8.34 |
| Fiber (g/100 g) | 10.57 | 12.69 | 11.23 |
| Sugar (g/100 g) | 0.26 | 0.52 | 0.32 |
| Sodium (Na) (mg/100 g) | 27.20 | 65.67 | 10.77 |
| Iron (Fe) (mg/100 g) | 0.11 | 0.12 | 0.14 |
| Calcium (Ca) (mg/100 g) | 3.36 | 4.6 | 3.47 |
| Potassium (K) (mg/100 g) | 22.40 | 54.37 | 33.45 |
| Ash (g/100 g) | 2.37 | 4.88 | 2.87 |
| Moisture (g/100 g) | 66.40 | 57.54 | 30.65 |
| pH | 6.43 | 6.70 | 6.29 |
| Water activity (aw) | 0.85 | 0.83 | 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yongsawas, R.; In-on, A.; Inta, A.; Kampuansai, J.; Pandith, H.; Suwannarach, N.; Lumyong, S.; Chitov, T.; Disayathanoowat, T. Bacterial Communities in Lanna Fermented Soybeans from Three Different Ethnolinguistic Groups in Northern Thailand. Microorganisms 2023, 11, 649. https://doi.org/10.3390/microorganisms11030649
Yongsawas R, In-on A, Inta A, Kampuansai J, Pandith H, Suwannarach N, Lumyong S, Chitov T, Disayathanoowat T. Bacterial Communities in Lanna Fermented Soybeans from Three Different Ethnolinguistic Groups in Northern Thailand. Microorganisms. 2023; 11(3):649. https://doi.org/10.3390/microorganisms11030649
Chicago/Turabian StyleYongsawas, Rujipas, Ammarin In-on, Angkana Inta, Jatupol Kampuansai, Hataichanok Pandith, Nakarin Suwannarach, Saisamorn Lumyong, Thararat Chitov, and Terd Disayathanoowat. 2023. "Bacterial Communities in Lanna Fermented Soybeans from Three Different Ethnolinguistic Groups in Northern Thailand" Microorganisms 11, no. 3: 649. https://doi.org/10.3390/microorganisms11030649