
A Comparative Analysis of Biodegradation and Bioconversion of Lentinula edodes and Other Exotic Mushrooms
 and
and 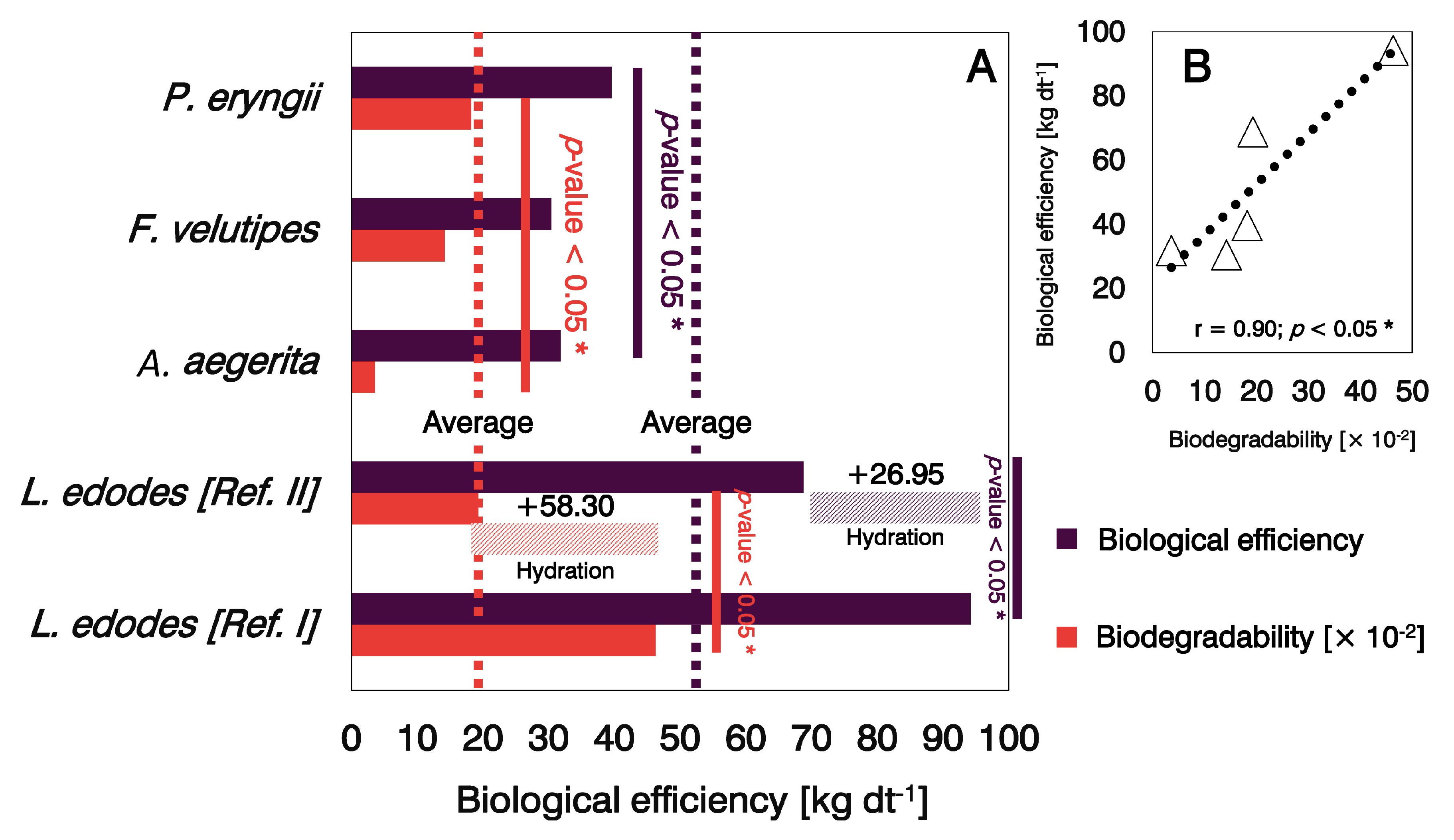{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Individual Growing Cycle Description
2.3. Chemical Analysis
2.4. Data Analyses
3. Results and Discussion
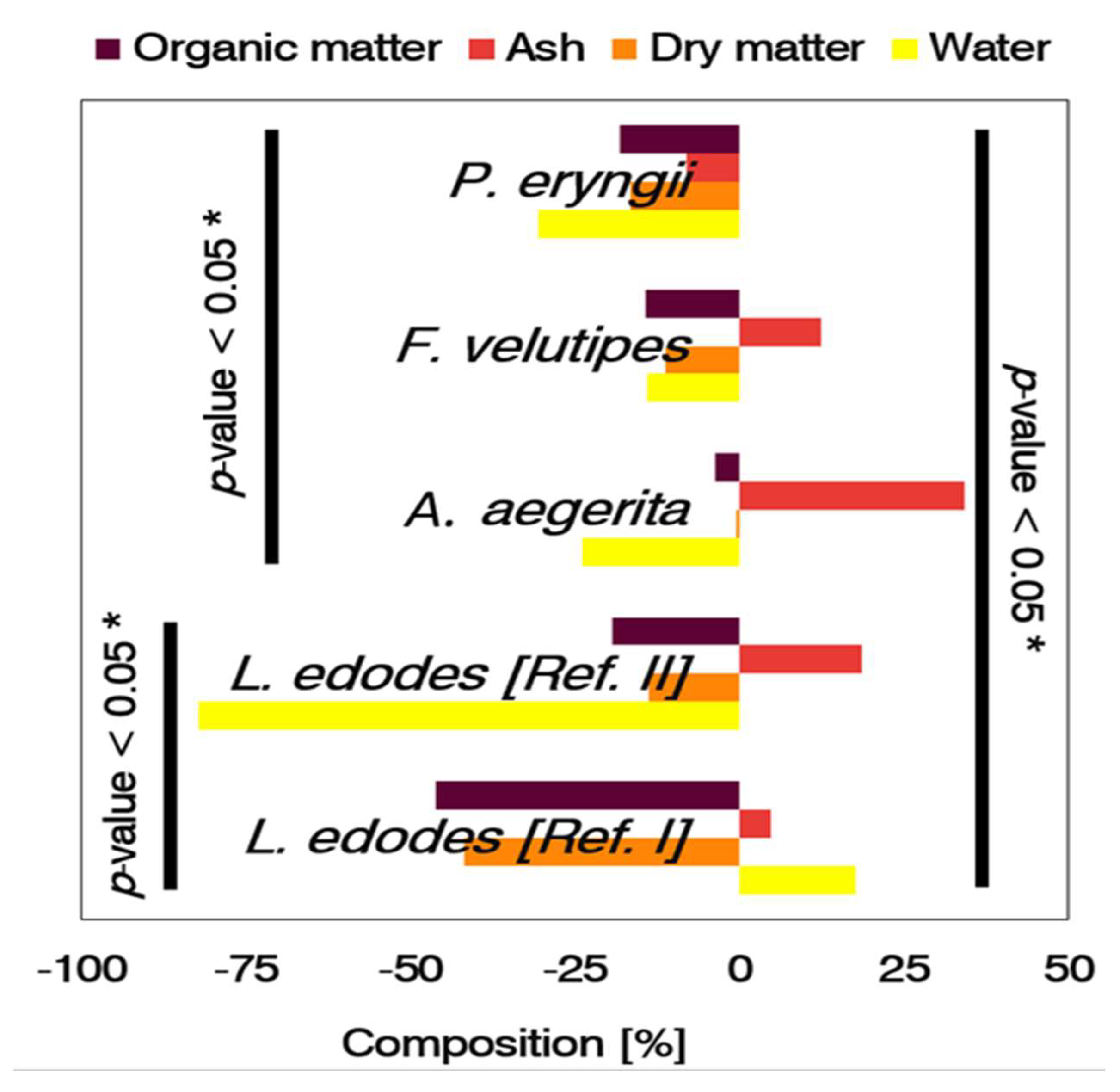
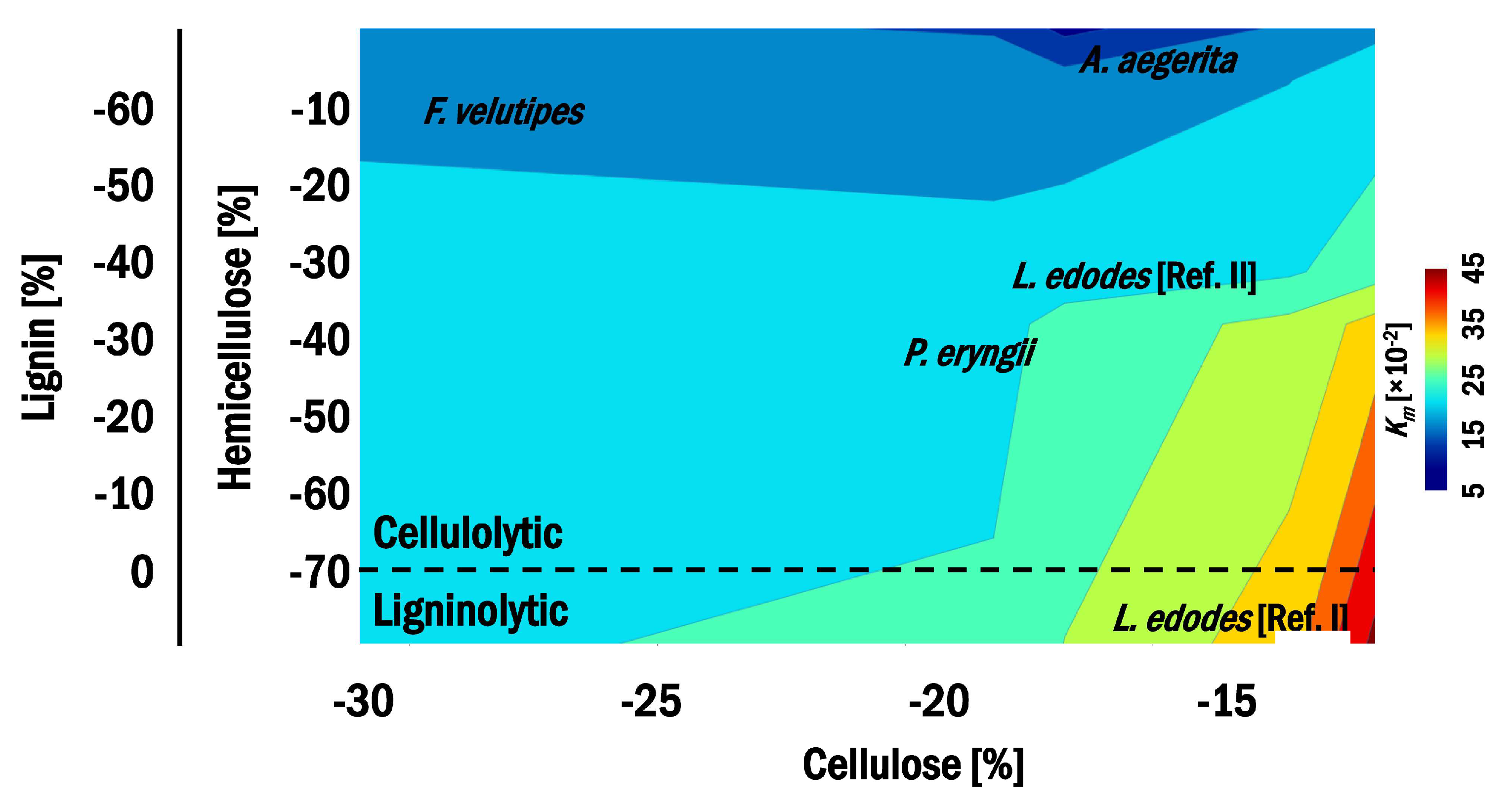
3.1. Biodegradability and Biological Efficiency
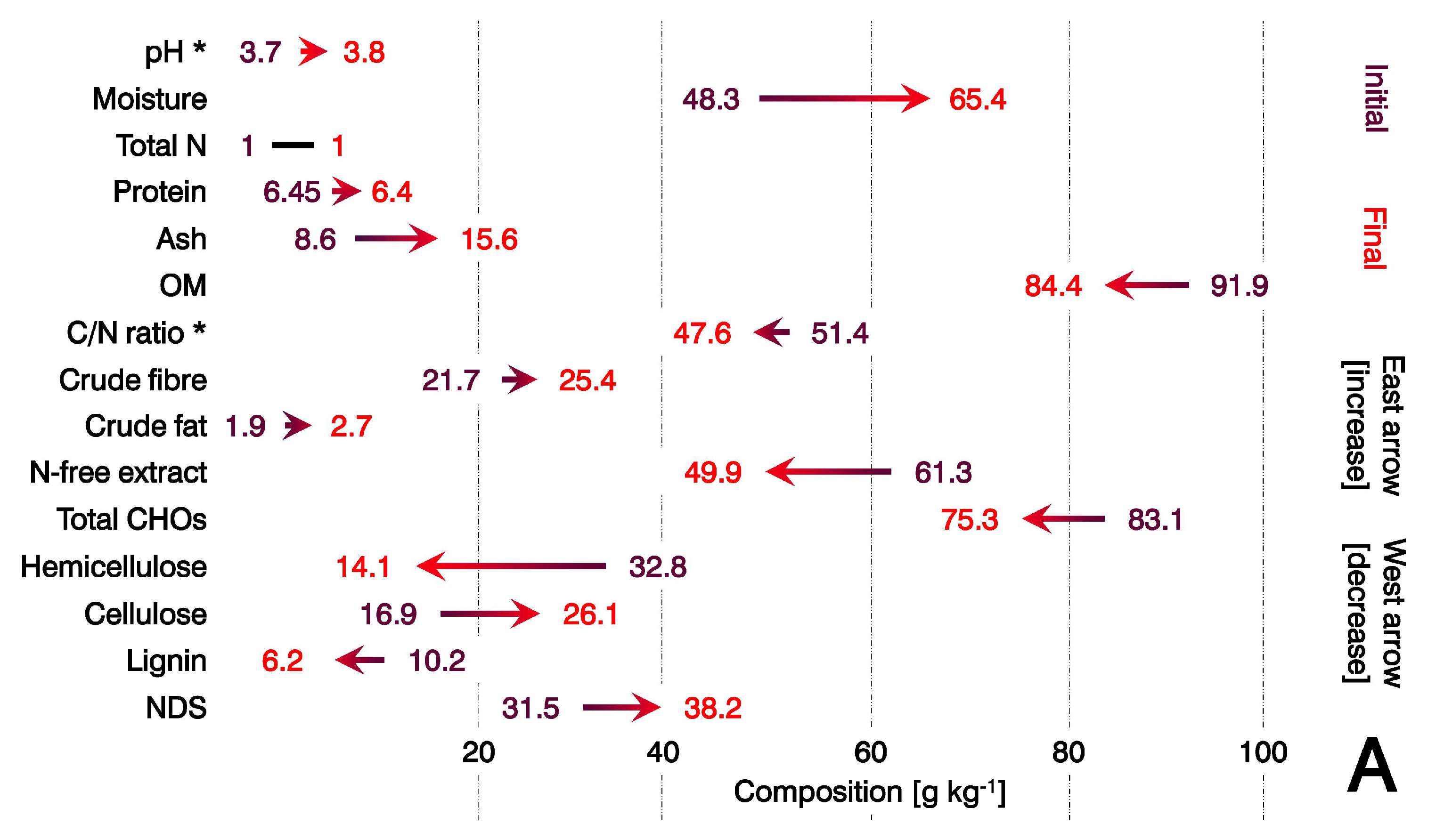
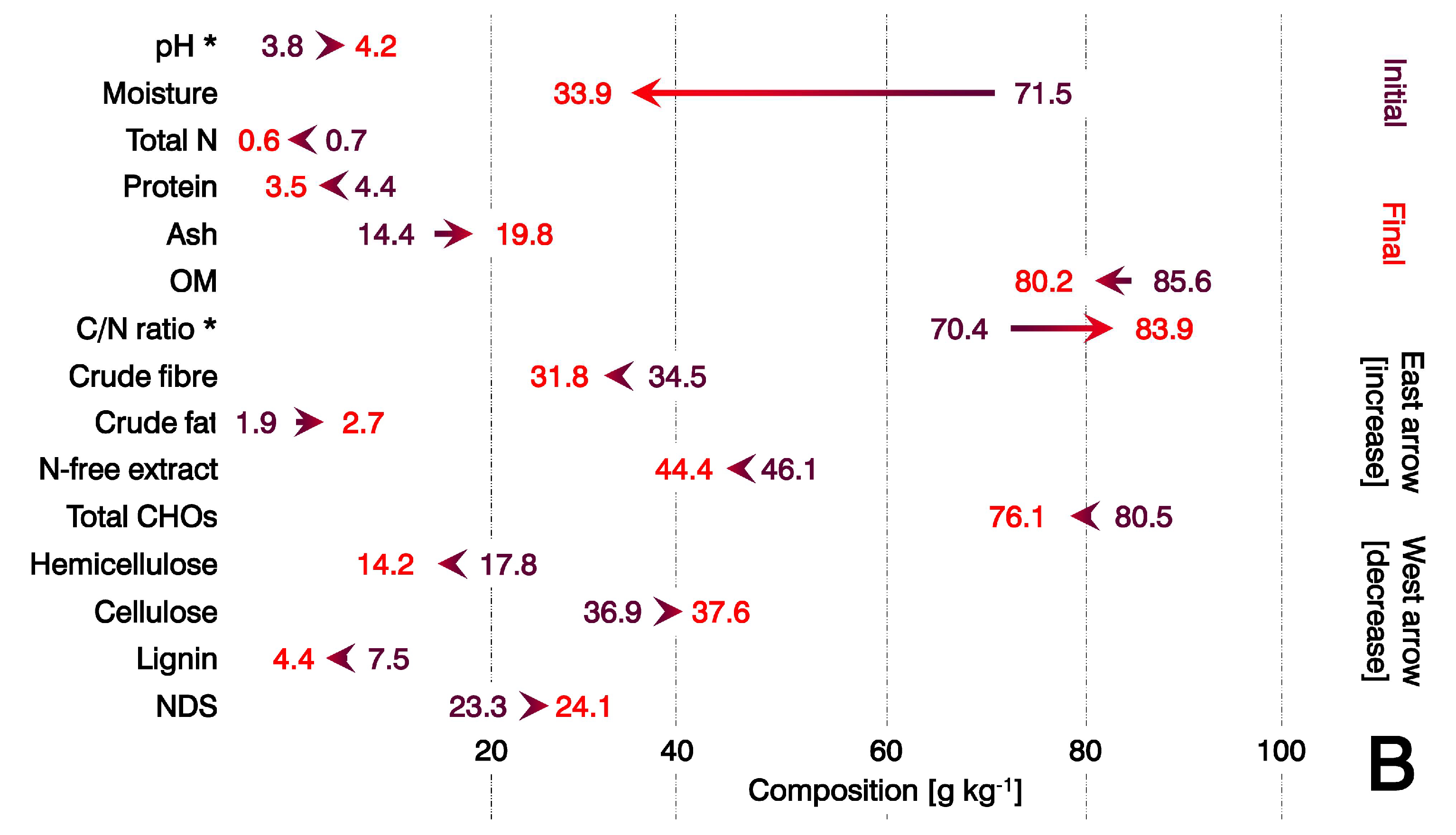
3.2. Sensitivity of the Substrate to Biotransformation
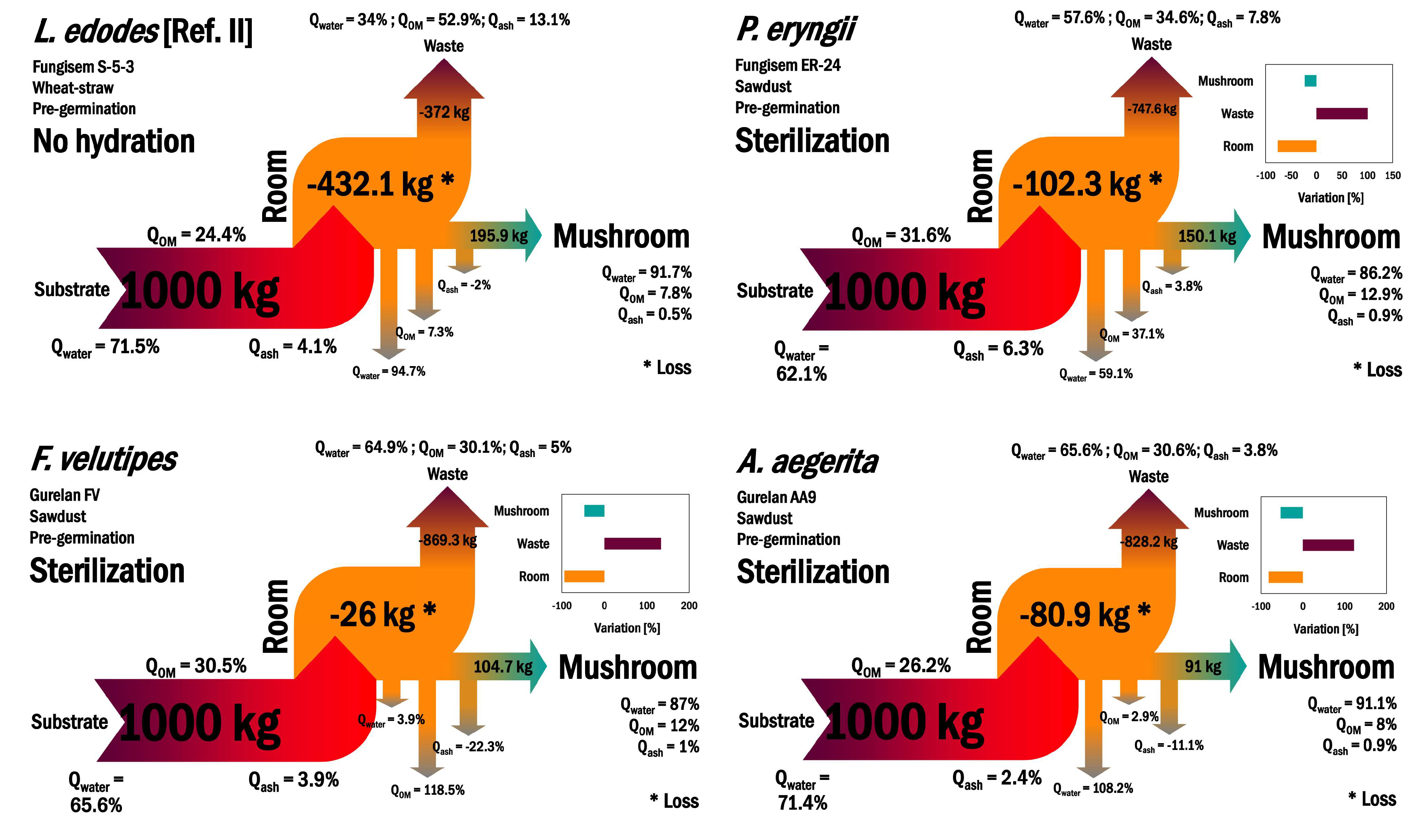
3.3. Technical Feasibility of Mushroom-Producing Systems on the Balance of Mass
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of Food and Agriculture 2020. In Overcoming Water Challenges in Agriculture; FAO: Rome, Italy, 2020. [Google Scholar]
- Bahar, N.; Lo, M.; Sanjaya, M.; Van Vianen, J.; Alexander, P.; Ickowitz, A.; Sunderland, T. Meeting the food security challenge for nine billion people in 2050: What impact on forests? Glob. Environ. Chang. 2020, 62, 102056. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Birowo, A.T.; Hansen, G.E. Agricultural and Rural Development: An Overview. In Agricultural and Rural Development in Indonesia, 1st ed.; Hansen, G.E., Ed.; Routledge: New York, NY, USA, 2019; Volume 1, pp. 1–27. [Google Scholar]
- Zhang, Y.; Kusch-Brandt, S.; Salter, A.M.; Heaven, S. Estimating the methane potential of energy crops: An overview on types of data sources and their limitations. Processes 2021, 9, 1565. [Google Scholar] [CrossRef]
- Fernandes, Â.; Antonio, A.L.; Oliveira, M.B.P.P.; Martins, A.; Ferreira, I.C.F.R. Effect of gamma and electron beam irradiation on the physico-chemical and nutritional properties of mushrooms: A review. Food Chem. 2012, 135, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.; Jang, K.-Y.; Oh, Y.-L.; Oh, M.; Im, J.-H.; Lakshmanan, H.; Sabaratnam, V. Cultivation and nutritional value of prominent Pleurotus spp.: An overview. Mycobiology 2021, 49, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.; Zied, D.C.; Pardo, J.E.; Preston, G.M.; Pardo-Giménez, A. Supplementation in mushroom crops and its impact on yield and quality. AMB Express 2018, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortei, N.K.; Odamtten, G.T.; Obodai, M.; Wiafe-Kwagyan, M.; Prempeh, J. Survey of mushroom consumption and the possible use of gamma irradiation for sterilization of compost for its cultivation in Southern Ghana. Agric. Food Secur. 2018, 7, 83. [Google Scholar] [CrossRef]
- Kaliyaperumal, M.; Kezo, K.; Gunaseelan, S. A Global Overview of Edible Mushrooms. In Biology of Macrofungi, Fungal Biology; Singh, B.P., Lallawmsanga, Passari, A.K., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, pp. 15–56. [Google Scholar]
- Kapahi, M. Recent Advances in Cultivation of Edible Mushrooms. In Biology of Macrofungi, Fungal Biology; Singh, B.P., Lallawmsanga, Passari, A.K., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, pp. 275–286. [Google Scholar]
- Xiong, S.; Martín, C.; Eilertsen, L.; Wei, M.; Myronycheva, O.; Larsson, S.H.; Lestander, T.A.; Atterhem, L.; Jönsson, L.J. Energy-efficient substrate pasteurisation for combined production of shiitake mushroom (Lentinula edodes) and bioethanol. Bioresour. Technol. 2019, 274, 65–72. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current Overview of Mushroom Production in the World. In Edible and Medicinal Mushrooms; Zied, D.C., Pardo-Gimenez, A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 5–13. [Google Scholar]
- Jeffries, T.W. Biodegradation of lignin and hemicelluloses. In Biochemistry of Microbial Degradation; Ratledge, C., Ed.; Springer: Dordrecht, The Netherlands, 1994; pp. 233–277. [Google Scholar]
- Schimpf, U.; Schulz, R. Industrial by-products from white-rot fungi production. Part I: Generation of enzyme preparations and chemical, protein biochemical and molecular biological characterization. Process 2016, 51, 2034–2046. [Google Scholar] [CrossRef]
- Mata, G.; Savoie, J.-M. Shiitake Cultivation on Straw: An Alternative for Subtropical Regions. In Updates on Tropical Mushrooms: Basic and Applied Research; Sánchez, J.E., Mata, G., Royse, D.J., Eds.; El Colegio de la Frontera Sur: San Cristóbal de Las Casas, México, 2018; pp. 115–134. [Google Scholar]
- Ohga, S. Effect of water potential on fruit body formation of Lentinula edodes in sawdust-based substrate. J. Wood Sci. 1999, 45, 337–342. [Google Scholar] [CrossRef]
- Gaitán-Hernández, R.; Mata, G. Cultivation of the edible mushroom Lentinula edodes (shiitake) in pasteurized wheat straw: Alternative use of geothermal energy in Mexico. Eng. Life Sci. 2004, 4, 363–367. [Google Scholar]
- Sánchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liang, J.; Wang, Y.; Sun, F.; Tao, H.; Xu, Q.; Zhang, L.; Zhang, Z.; Ho, C.-T.; Wan, X. Tea waste: An effective and economic substrate for oyster mushroom cultivation. J. Sci. Food Agric. 2016, 96, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Kumla, J.; Suwannarach, N.; Sujarit, K.; Penkhrue, W.; Kakumyan, P.; Jatuwong, K.; Vadthanarat, S.; Lumyong, S. Cultivation of mushrooms and their lignocellulolytic enzyme production through the utilization of agro-industrial waste. Molecules 2020, 25, 2811. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Wang, X.; Zhen, L.; Gu, J.; Zhang, K.; Wang, Q.; Zhao, W. Effects of inoculating with lignocellulose-degrading consortium on cellulose-degrading genes and fungal community during co-composting of spent mushroom substrate with swine manure. Bioresour. Technol. 2019, 291, 121876. [Google Scholar] [CrossRef]
- Carrasco-Cabrera, C.P.; Bell, T.L.; Kertesz, M.A. Caffeine metabolism during cultivation of oyster mushroom (Pleurotus ostreatus) with spent coffee grounds. Appl. Microbiol. Biotechnol. 2019, 103, 5831–5841. [Google Scholar] [CrossRef]
- Elisashvili, V.; Penninckx, M.; Kachlishvili, E.; Asatiani, M.; Kvesitadze, G. Use of Pleurotus dryinus for lignocellulolytic enzymes production in submerged fermentation of mandarin peels and tree leaves. Enzyme Microb. Technol. 2006, 38, 998–1004. [Google Scholar] [CrossRef]
- O’Brien, B.J.; Milligan, E.; Carver, J.; Roy, E.D. Integrating anaerobic co-digestion of dairy manure and food waste with cultivation of edible mushrooms for nutrient recovery. Bioresour. Technol. 2019, 285, 121312. [Google Scholar] [CrossRef]
- Stoknes, K.; Beyer, D.M.; Norgaard, E. Anaerobically digested food waste in compost for Agaricus bisporus and Agaricus subrufescens and its effect on mushroom productivity. J. Sci. Food Agric. 2013, 93, 2188–2200. [Google Scholar] [CrossRef]
- Szumigaj-Tarnowska, J.; Szafranek, P.; Uliński, Z.; Ślusarski, C. Efficiency of gaseous ozone in disinfection of mushroom growing rooms. J. Hortic. Res. 2020, 28, 91–100. [Google Scholar] [CrossRef]
- Pardo-Giménez, A.; Pardo-González, J.E. Evaluation of casing materials made from spent mushroom substrate and coconut fibre pith for use in production of Agaricus bisporus (Lange) Imbach. Span. J. Agric. Res. 2008, 6, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Haug, R.T. The Practical Handbook of Compost Engineering; Lewis Publishers: Boca Raton, FL, USA, 1993. [Google Scholar]
- Zied, D.C.; Pardo, J.E.; Miasaki, C.; Tomaz, R.; Pardo-Giménez, A. Mycochemical Characterization of considering Their Morphological and Physiological Stage of Maturity on the Traceability Process. BioMed Res. Int. 2017, 2017, 2713742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, O. Methods of Chemical Analysis of Mushrooms. In Tropical Mushrooms: Biological Nature and Cultivation Methods; Chang, S.T., Quimio, T.H., Eds.; The Chinese University Press: Hong Kong, China, 1982; pp. 87–116. [Google Scholar]
- Sullivan, D.M. Proximate and Mineral Analysis. In Methods of Analysis for Nutrition Labeling; Sullivan, D.M., Carpenter, D.E., Eds.; AOAC International: Arlington, TX, USA, 1993; pp. 105–109. [Google Scholar]
- Fornito, S.; Puliga, F.; Leonardi, P.; Di Foggia, M.; Zambonelli, A.; Francioso, O. Degradative ability of mushrooms cultivated on corn silage digestate. Molecules 2020, 25, 3020. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, R.; Yang, Q.; Li, X.; Xiang, Q.; Zhao, K.; Ma, M.; Yu, X.; Chen, Q.; Zeng, X.; et al. Cultivating Lentinula edodes on substrate containing composted sawdust affects the expression of carbohydrate and aromatic amino acid metabolism-related genes. mSystems 2022, 7, e00827-21. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, D.; Zhang, L.; Li, Q.; Song, C.; Shang, X.; Bao, D.; Tan, Q.; Chen, H.; Lv, B. Corncob as a substrate for the cultivation of Lentinula edodes. Waste Biomass Valoriz. 2022, 13, 929–939. [Google Scholar] [CrossRef]
- Ma, N.L.; Khoo, S.C.; Peng, W.; Ng, C.M.; Teh, C.H.; Park, Y.-K.; Lam, S.S. Green application and toxic risk of used diaper and food waste as growth substitute for sustainable cultivation of oyster mushroom (Pleurotus ostreatus). J. Clean. Prod. 2022, 268, 122272. [Google Scholar] [CrossRef]
- Pérez, J.; Muñoz-Dorado, J.; de la Rubia, T.; Martínez, J. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: An overview. Int. Microbiol. 2002, 5, 53–63. [Google Scholar] [CrossRef]
- Kuhad, R.C.; Singh, A.; Eriksson, K.-E.L. Microorganisms and Enzymes Involved in the Degradation of Plant Fiber Cell Walls. In Biotechnology in the Pulp and Paper Industry, Advances in Biochemical Engineering/Biotechnology; Eriksson, K.-E.L., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 45–125. [Google Scholar]
- Leonowicz, A.; Matuszewska, A.; Luterek, J.; Ziegenhagen, D.; Wojtaś-Wasilewska, M.; Cho, N.-S.; Hofrichter, M.; Rogalski, J. Biodegradation of lignin by white rot fungi. Fungal Genet. Biol. 1999, 27, 175–185. [Google Scholar] [CrossRef]
- Rabinovich, M.L.; Melnick, M.S.; Bolobova, A.V. The structure and mechanism of action of cellulolytic enzymes. Biochemistry 2002, 67, 850–871. [Google Scholar]
- Cullen, D.; Kersten, P.J. Enzymology and Molecular Biology of Lignin Degradation. In Biochemistry and Molecular Biology, The Mycota; Brambl, R., Marzluf, G.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 249–273. [Google Scholar]
- Jensen, K.A.; Bao, W.; Kawai, S.; Srebotnik, E.; Hammel, K.E. Manganese-dependent cleavage of nonphenolic lignin structures by Ceriporiopsis subvermispora in the absence of lignin peroxidase. Appl. Environ. Microbiol. 1996, 62, 3679–3686. [Google Scholar] [CrossRef] [Green Version]
- Hammel, K.E. Fungal Degradation of Lignin. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Wallingford, UK, 1997; pp. 33–45. [Google Scholar]
- Goyal, A.; Ghosh, B.; Eveleigh, D. Characteristics of fungal cellulases. Bioresour. Technol. 1991, 36, 37–50. [Google Scholar] [CrossRef]
- Esterbauer, H.; Steiner, W.; Labudova, I.; Hermann, A.; Hayn, M. Production of Trichoderma cellulase in laboratory and pilot scale. Bioresour. Technol. 1991, 36, 51–65. [Google Scholar] [CrossRef]
- Malherbe, S.; Cloete, T.E. Lignocellulose biodegradation: Fundamentals and applications. Rev. Environ. Sci. Biotechnol. 2002, 1, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Gong, W.; Yan, L.; Zhu, Z.; Hu, Z.; Peng, Y. Biodegradation of ramie stalk by Flammulina velutipes: Mushroom production and substrate utilization. AMB Express 2017, 7, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazvinian, A.; Farrokhsiar, P.; Vieira, F.; Pecchia, J.; Gursoy, B. Mycelium-Based Bio-Composites for Architecture: Assessing the Effects of Cultivation Factors on Compressive Strength Background on Designing with Mycelium-Based Bio-Composites What is a Mycelium-based Bio-Composite? Mater. Stud. Innov. 2019, 2, 505–514. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zied, D.C.; da Silva Freitas, M.A.; de Almeida Moreira, B.R.; da Silva Alves, L.; Pardo-Giménez, A. A Comparative Analysis of Biodegradation and Bioconversion of Lentinula edodes and Other Exotic Mushrooms. Microorganisms 2023, 11, 897. https://doi.org/10.3390/microorganisms11040897
Zied DC, da Silva Freitas MA, de Almeida Moreira BR, da Silva Alves L, Pardo-Giménez A. A Comparative Analysis of Biodegradation and Bioconversion of Lentinula edodes and Other Exotic Mushrooms. Microorganisms. 2023; 11(4):897. https://doi.org/10.3390/microorganisms11040897
Chicago/Turabian StyleZied, Diego Cunha, Marcos Antônio da Silva Freitas, Bruno Rafael de Almeida Moreira, Lucas da Silva Alves, and Arturo Pardo-Giménez. 2023. "A Comparative Analysis of Biodegradation and Bioconversion of Lentinula edodes and Other Exotic Mushrooms" Microorganisms 11, no. 4: 897. https://doi.org/10.3390/microorganisms11040897