

Microbiota Phenotype Promotes Anastomotic Leakage in a Model of Rats with Ischemic Colon Resection
 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
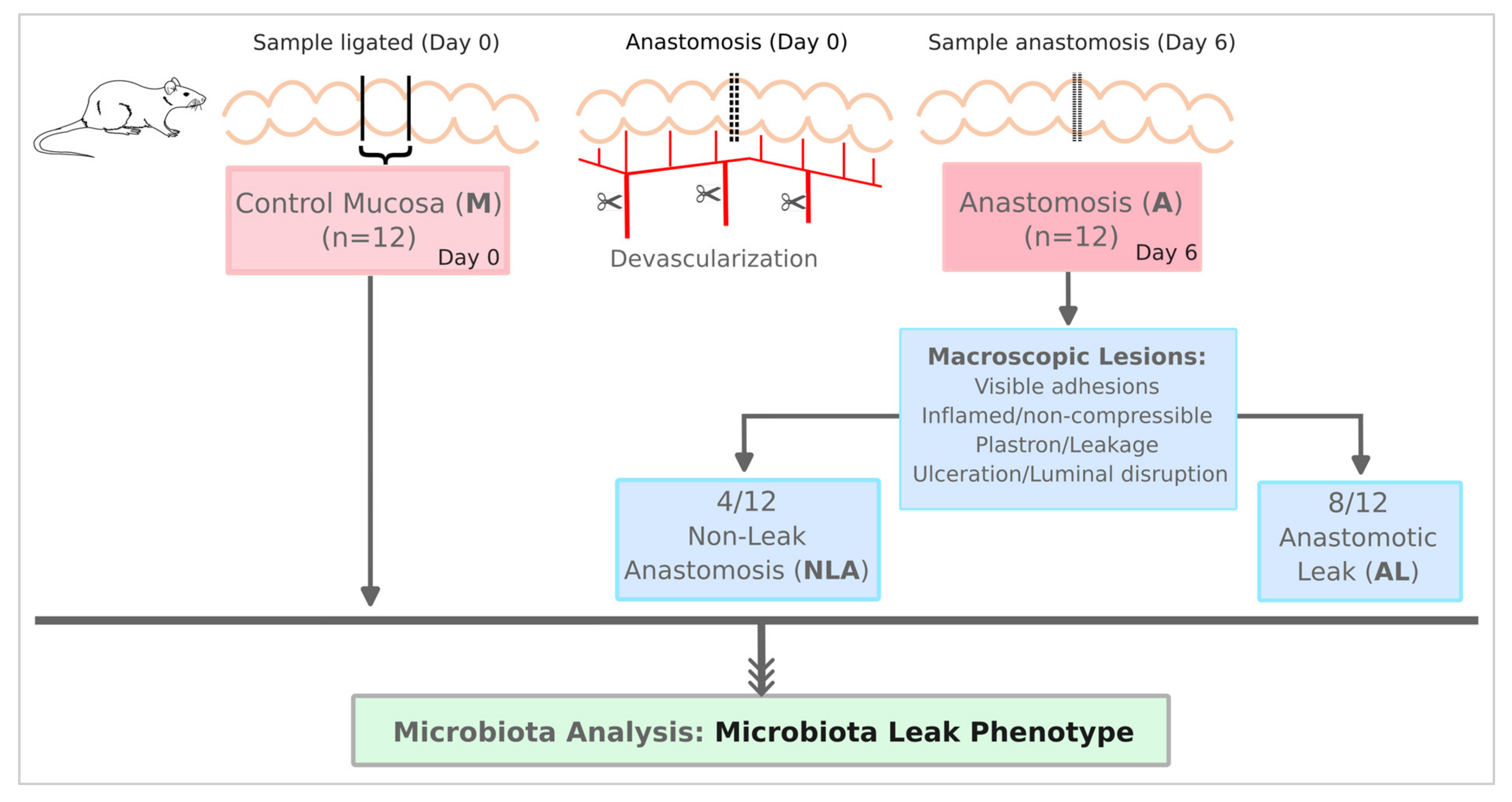
2.1. Animal Model
2.2. Sample Processing
2.3. Primers, 16S rRNA Amplification, and Sequencing Procedure
2.4. Processing and OTU (Operational Taxonomic Units) Grouping
2.5. Phylogenetic Affiliation and OPU (Operational Phylogenetic Unit) Clustering
2.6. Alpha Diversity, Beta Diversity, Comparative, and Statistical Analyses
3. Results
3.1. Samples, Sequences, and OPUs Distribution
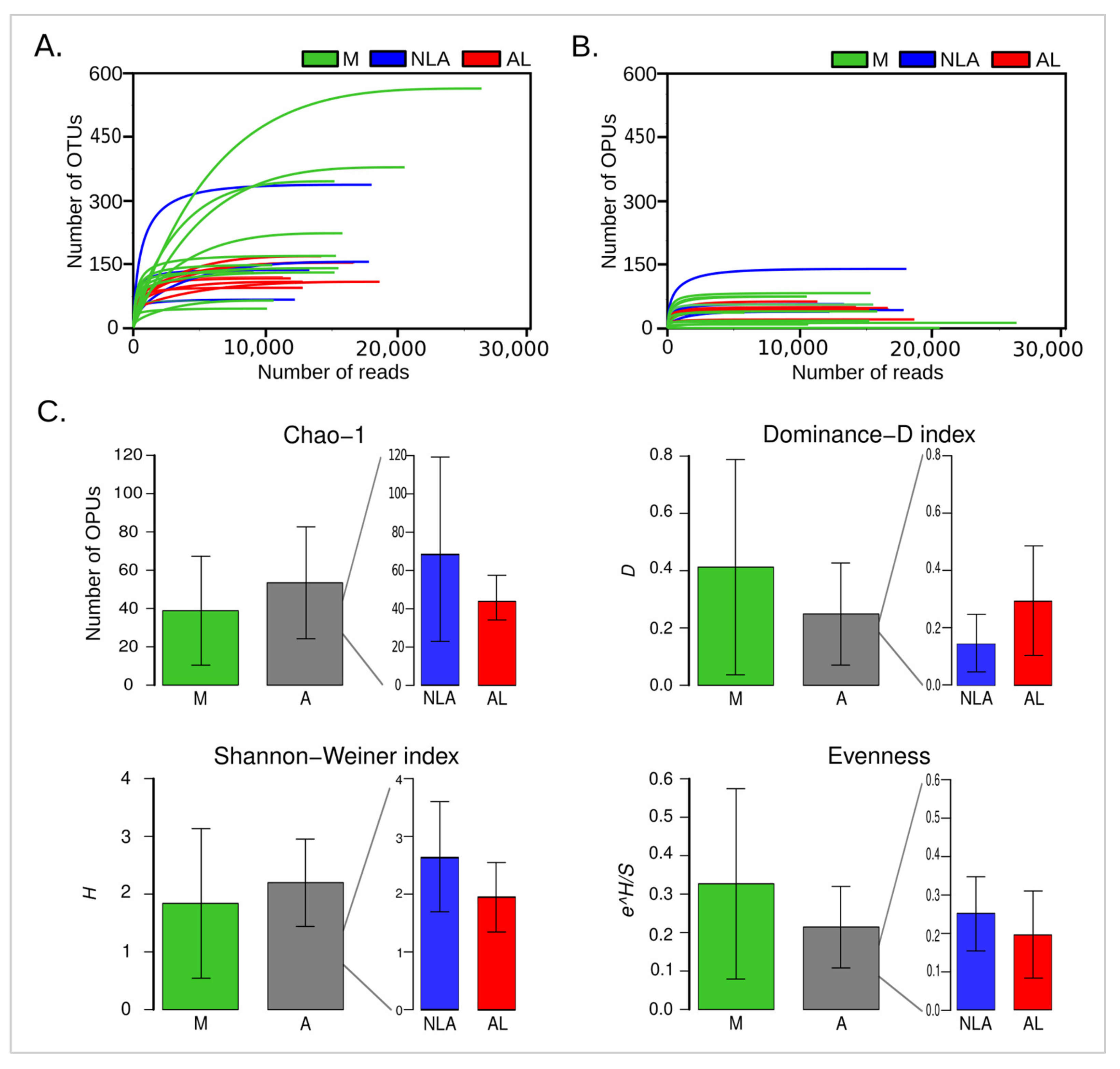
3.2. Alpha Diversity Involvement in Anastomotic Leak
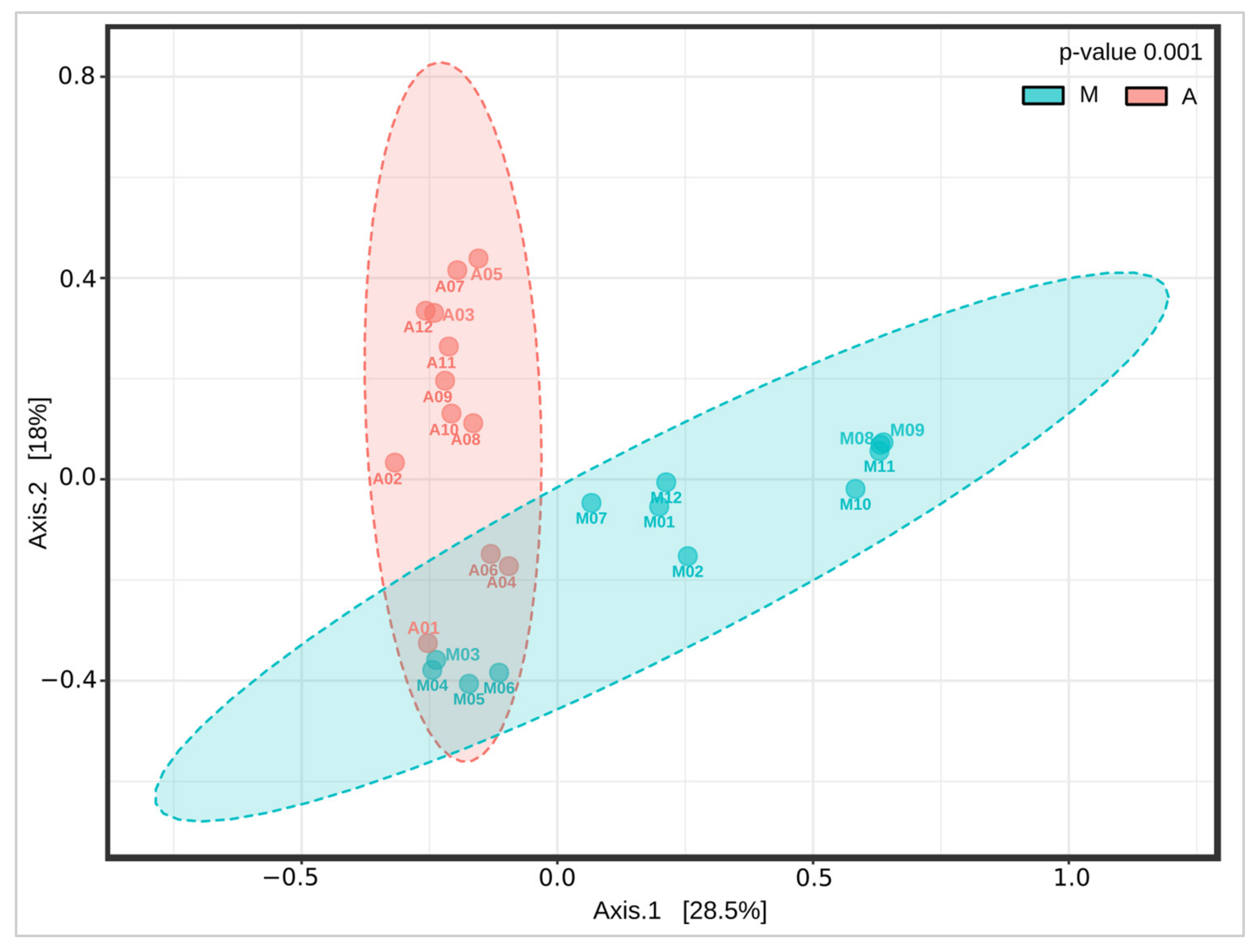
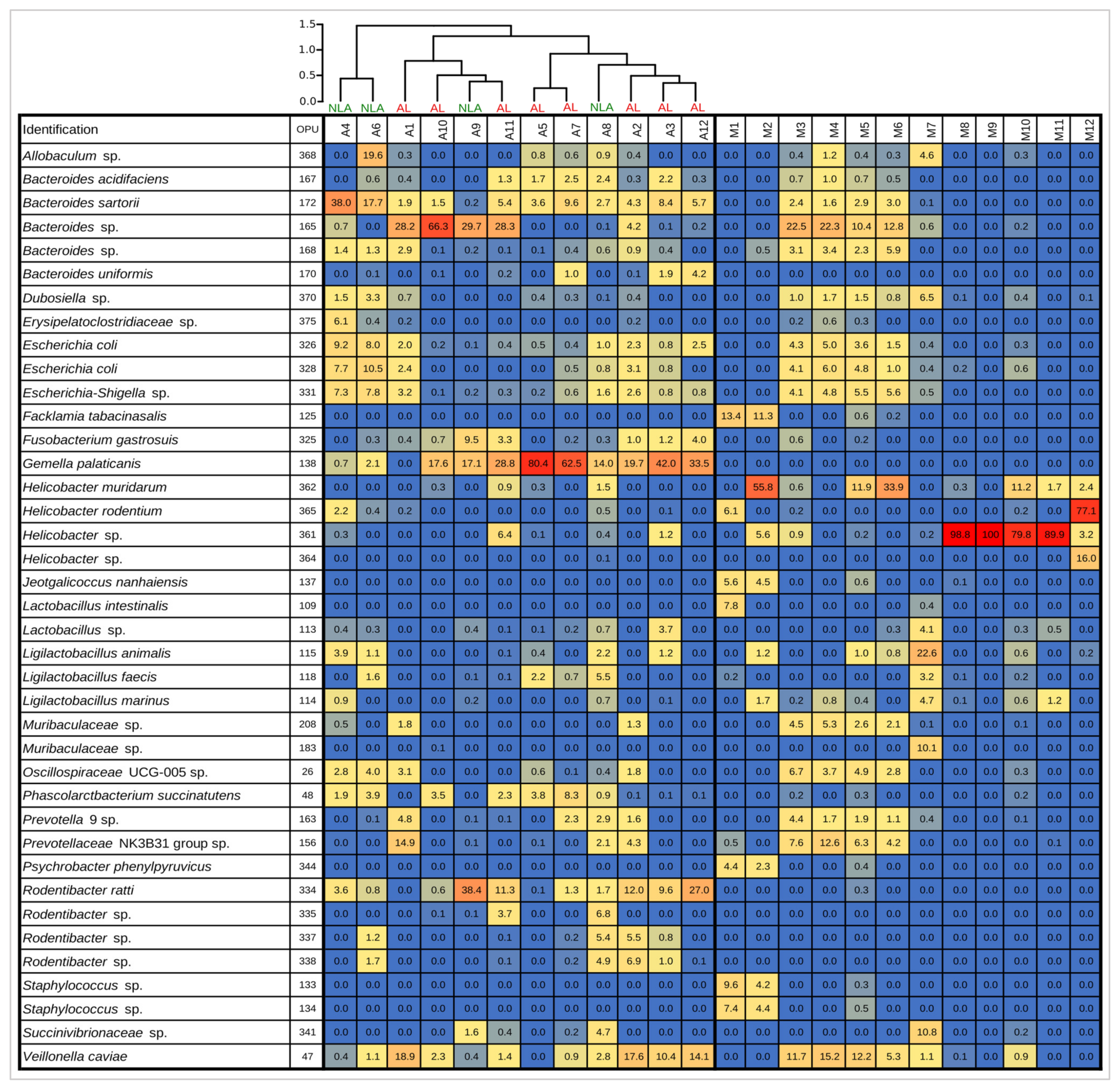
3.3. Changes in the Intestinal Microbiota Associated with Surgical Anastomosis
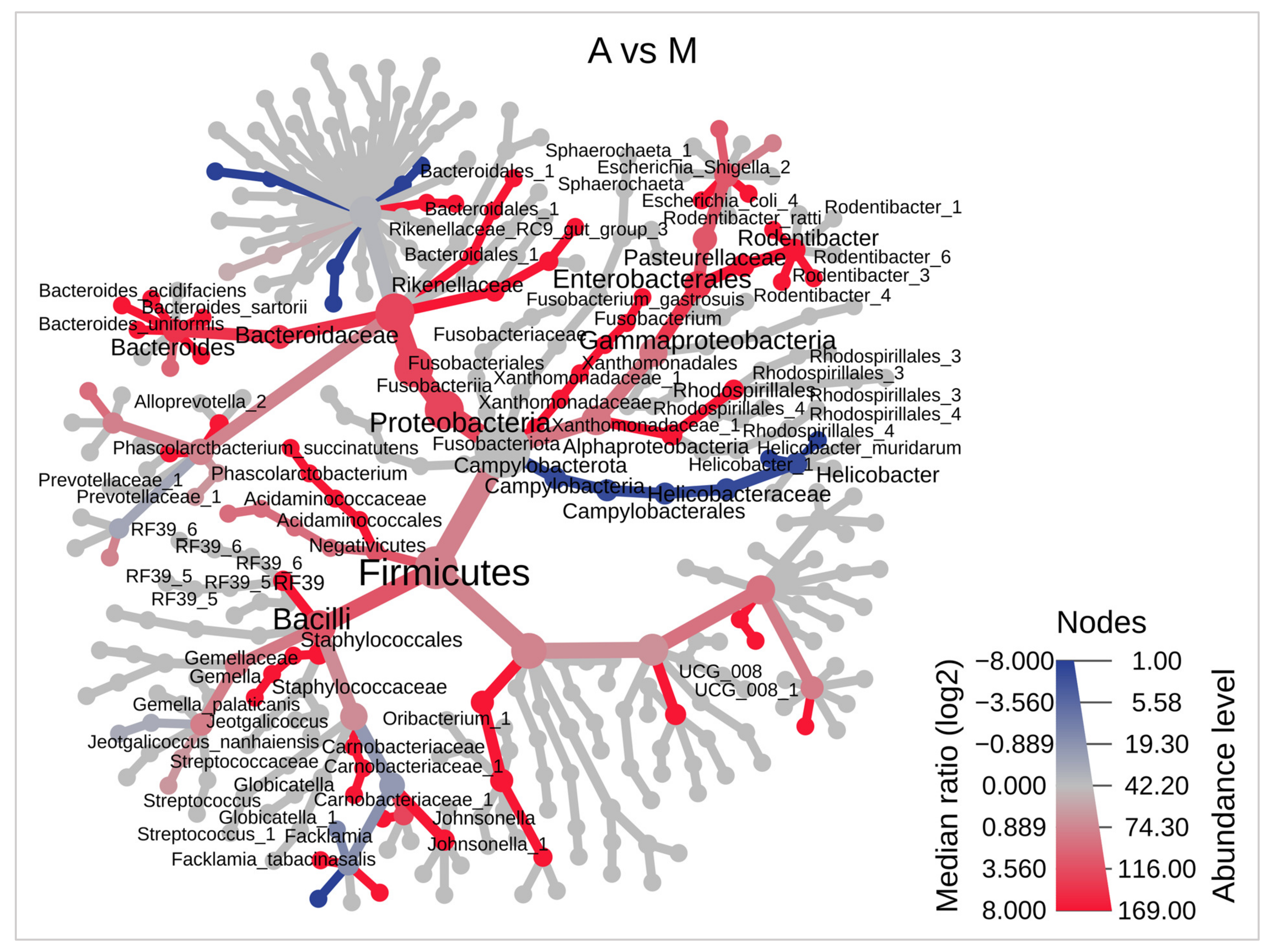
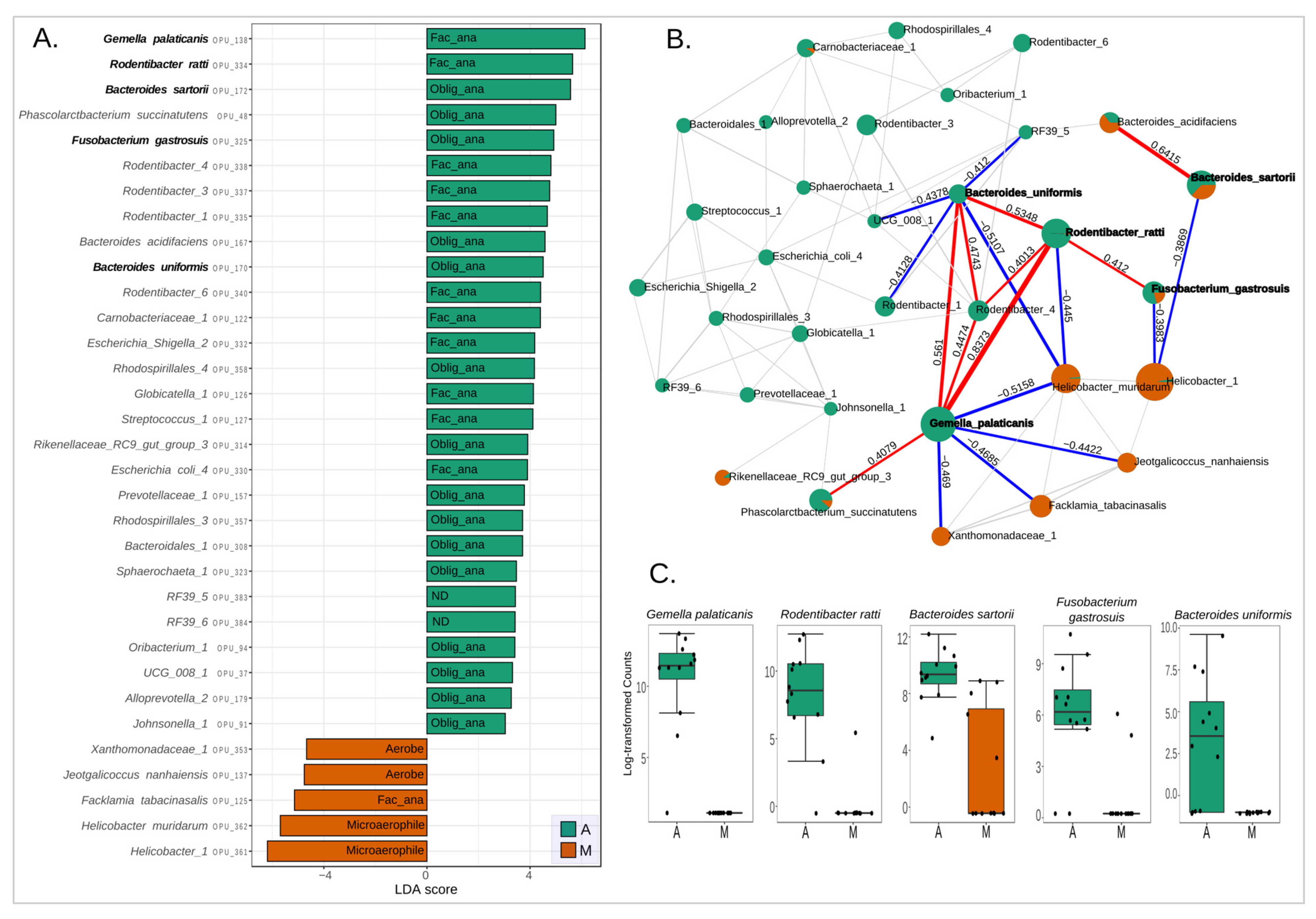
3.4. Bacterial Biomarkers of Anastomotic Intervention
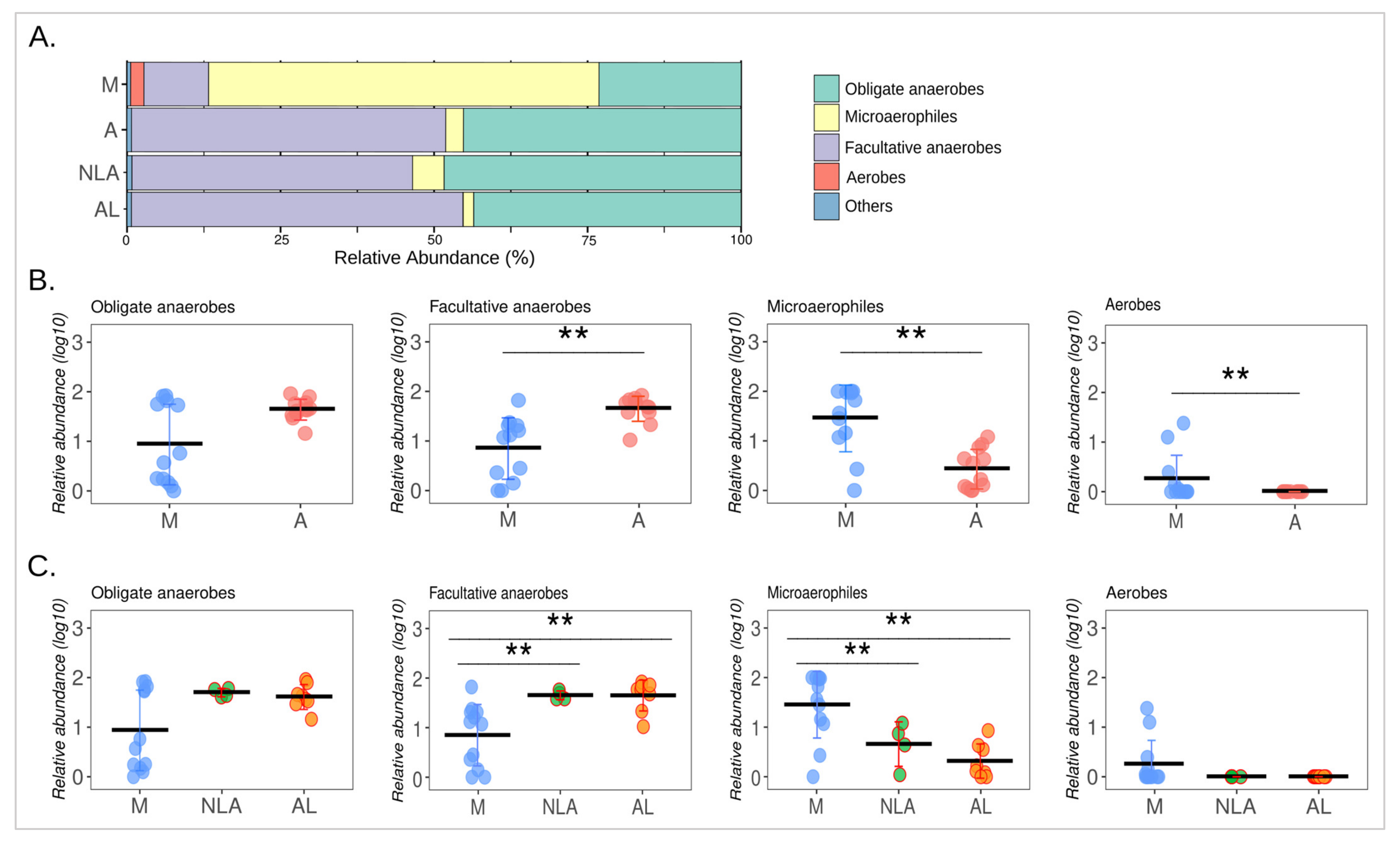
3.5. Changes in Microbial Respiration of the Intestinal Microbiota
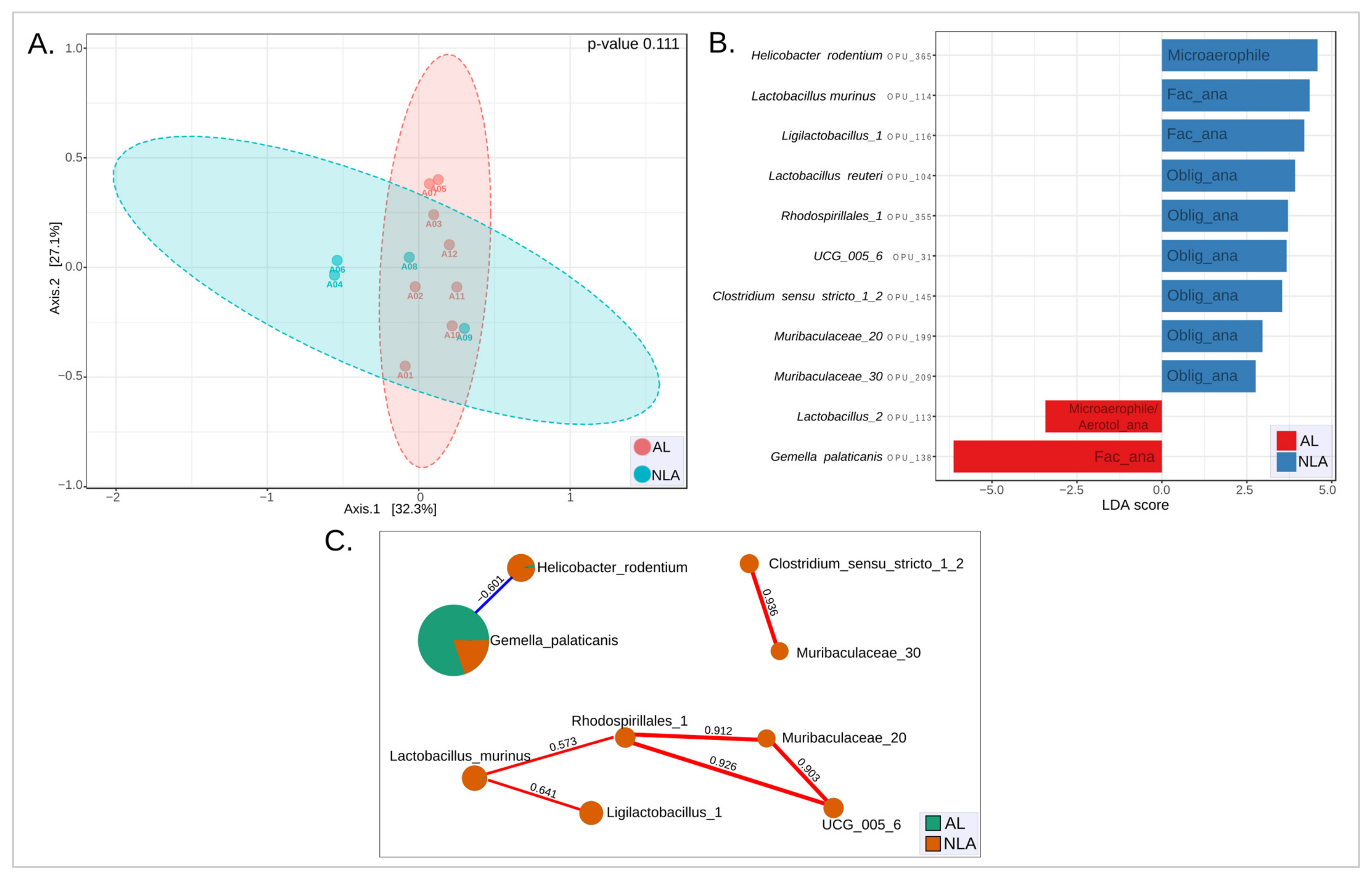
3.6. Microbiota Leak Phenotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paun, B.C.; Cassie, S.; MacLean, A.R.; Dixon, E.; Buie, W.D. Postoperative Complications Following Surgery for Rectal Cancer. Ann. Surg. 2010, 251, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Goligher, J.C.; Graham, N.G.; De Dombal, F.T. Anastomotic Dehiscence after Anterior Resection of Rectum and Sigmoid. Br. J. Surg. 2005, 57, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, J.W.; Dietz, D.W.; Stocchi, L. Anastomotic Leak after Restorative Proctosigmoidectomy for Cancer: What Are the Chances of a Permanent Ostomy? Int. J. Color. Dis. 2012, 27, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Buchs, N.C.; Gervaz, P.; Secic, M.; Bucher, P.; Mugnier-Konrad, B.; Morel, P. Incidence, Consequences, and Risk Factors for Anastomotic Dehiscence after Colorectal Surgery: A Prospective Monocentric Study. Int. J. Color. Dis. 2008, 23, 265–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McArdle, C.S.; McMillan, D.C.; Hole, D.J. Impact of Anastomotic Leakage on Long-Term Survival of Patients Undergoing Curative Resection for Colorectal Cancer. Br. J. Surg. 2005, 92, 1150–1154. [Google Scholar] [CrossRef]
- Wong, N.Y.; Eu, K.W. A Defunctioning Ileostomy Does Not Prevent Clinical Anastomotic Leak After a Low Anterior Resection: A Prospective, Comparative Study. Dis. Colon Rectum 2005, 48, 2076–2079. [Google Scholar] [CrossRef]
- Hallböök, O.; Sjödahl, R. Anastomotic Leakage and Functional Outcome after Anterior Resection of the Rectum. Br. J. Surg. 2005, 83, 60–62. [Google Scholar] [CrossRef]
- Nesbakken, A.; Nygaard, K.; Lunde, O.C. Outcome and Late Functional Results after Anastomotic Leakage Following Mesorectal Excision for Rectal Cancer. Br. J. Surg. 2002, 88, 400–404. [Google Scholar] [CrossRef]
- Mirnezami, A.; Mirnezami, R.; Chandrakumaran, K.; Sasapu, K.; Sagar, P.; Finan, P. Increased Local Recurrence and Reduced Survival from Colorectal Cancer Following Anastomotic Leak: Systematic Review and Meta-Analysis. Ann. Surg. 2011, 253, 890–899. [Google Scholar] [CrossRef]
- Shogan, B.D.; Carlisle, E.M.; Alverdy, J.C.; Umanskiy, K. Do We Really Know Why Colorectal Anastomoses Leak? J. Gastrointest. Surg. 2013, 17, 1698–1707. [Google Scholar] [CrossRef]
- Cohn, I.; Rives, J.D. Antibiotic Protection of Colon Anastomoses*. Ann. Surg. 1955, 141, 707–717. [Google Scholar] [CrossRef]
- Shogan, B.D.; Smith, D.P.; Christley, S.; Gilbert, J.A.; Zaborina, O.; Alverdy, J.C. Intestinal Anastomotic Injury Alters Spatially Defined Microbiome Composition and Function. Microbiome 2014, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Shogan, B.D.; Belogortseva, N.; Luong, P.M.; Zaborin, A.; Lax, S.; Bethel, C.; Ward, M.; Muldoon, J.P.; Singer, M.; An, G.; et al. Collagen Degradation and MMP9 Activation by Enterococcus faecalis Contribute to Intestinal Anastomotic Leak. Sci. Transl. Med. 2015, 7, 286ra68. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, R.A.; Wienholts, K.; Williamson, A.J.; Gaines, S.; Hyoju, S.; van Goor, H.; Zaborin, A.; Shogan, B.D.; Zaborina, O.; Alverdy, J.C. Enterococcus faecalis Exploits the Human Fibrinolytic System to Drive Excess Collagenolysis: Implications in Gut Healing and Identification of Druggable Targets. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 318, G1–G9. [Google Scholar] [CrossRef] [Green Version]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.-H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the Classification of Cultured and Uncultured Bacteria and Archaea Using 16S RRNA Gene Sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Kashyap, S.K.; Maherchandani, S.; Kumar, N. Chapter 18—Ribotyping: A Tool for Molecular Taxonomy. In Animal Biotechnology; Verma, A.S., Singh, A., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 327–344. ISBN 978-0-12-416002-6. [Google Scholar]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; et al. ARB: A Software Environment for Sequence Data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate High-Throughput Multiple Sequence Alignment of Ribosomal RNA Genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [Green Version]
- Vidal, R.; Ginard, D.; Khorrami, S.; Mora-Ruiz, M.; Munoz, R.; Hermoso, M.; Díaz, S.; Cifuentes, A.; Orfila, A.; Rosselló-Móra, R. Crohn Associated Microbial Communities Associated to Colonic Mucosal Biopsies in Patients of the Western Mediterranean. Syst. Appl. Microbiol. 2015, 38, 442–452. [Google Scholar] [CrossRef]
- Chamorro, N.; Montero, D.A.; Gallardo, P.; Farfán, M.; Contreras, M.; De la Fuente, M.; Dubois, K.; Hermoso, M.A.; Quera, R.; Pizarro-Guajardo, M.; et al. Landscapes and Bacterial Signatures of Mucosa-Associated Intestinal Microbiota in Chilean and Spanish Patients with Inflammatory Bowel Disease. Microb. Cell 2021, 8, 223–238. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for Comprehensive Statistical, Functional, and Meta-Analysis of Microbiome Data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A Web-Based Tool for the Analysis of Sets through Venn Diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Ferrie, S.; Webster, A.; Wu, B.; Tan, C.; Carey, S. Gastrointestinal Surgery and the Gut Microbiome: A Systematic Literature Review. Eur. J. Clin. Nutr. 2021, 75, 12–25. [Google Scholar] [CrossRef]
- Van Praagh, J.B.; de Goffau, M.C.; Bakker, I.S.; Harmsen, H.J.M.; Olinga, P.; Havenga, K. Intestinal Microbiota and Anastomotic Leakage of Stapled Colorectal Anastomoses: A Pilot Study. Surg. Endosc. 2016, 30, 2259–2265. [Google Scholar] [CrossRef]
- Van Praagh, J.B.; de Goffau, M.C.; Bakker, I.S.; van Goor, H.; Harmsen, H.J.M.; Olinga, P.; Havenga, K. Mucus Microbiome of Anastomotic Tissue During Surgery Has Predictive Value for Colorectal Anastomotic Leakage. Ann. Surg. 2019, 269, 911–916. [Google Scholar] [CrossRef]
- Vomhof-DeKrey, E.E.; Stover, A.; Basson, M.D. Microbiome Diversity Declines While Distinct Expansions of Th17, INKT, and Dendritic Cell Subpopulations Emerge after Anastomosis Surgery. Gut Pathog. 2021, 13, 51. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, B.; Sun, Y.; Liu, Q.; Cao, Q.; Li, T.; Li, J. Risk Factors and Preventive Measures for Anastomotic Leak in Colorectal Cancer. Technol. Cancer Res. Treat. 2022, 21, 1–14. [Google Scholar] [CrossRef]
- Buffie, C.G.; Pamer, E.G. Microbiota-Mediated Colonization Resistance against Intestinal Pathogens. Nat. Rev. Immunol. 2013, 13, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yu, Y.; Wang, Y.; Wang, J.; Guan, R.; Sun, Y.; Shi, F.; Gao, J.; Fu, X. Role of SCFAs in Gut Microbiome and Glycolysis for Colorectal Cancer Therapy. J. Cell. Physiol. 2019, 234, 17023–17049. [Google Scholar] [CrossRef]
- The Human Microbiome Project Consortium. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Imlay, J.A. When Anaerobes Encounter Oxygen: Mechanisms of Oxygen Toxicity, Tolerance and Defence. Nat. Rev. Microbiol. 2021, 19, 774–785. [Google Scholar] [CrossRef]
- Agnes, A.; Puccioni, C.; D’Ugo, D.; Gasbarrini, A.; Biondi, A.; Persiani, R. The Gut Microbiota and Colorectal Surgery Outcomes: Facts or Hype? A Narrative Review. BMC Surg. 2021, 21, 83. [Google Scholar] [CrossRef]
- Ohigashi, S.; Sudo, K.; Kobayashi, D.; Takahashi, T.; Nomoto, K.; Onodera, H. Significant Changes in the Intestinal Environment After Surgery in Patients with Colorectal Cancer. J. Gastrointest. Surg. 2013, 17, 1657–1664. [Google Scholar] [CrossRef]
- Chow, P.K.H.; Ng, R.T.H.; Ogden, B.E. (Eds.) Animal Models in Biomedical Research: A Primer for the Investigator; World Scientific: Hackensack, NJ, USA, 2008; ISBN 978-981-270-663-8. [Google Scholar]
- Komen, N.; van der Wal, H.-C.; Ditzel, M.; Kleinrensink, G.-J.; Jeekel, H.; Lange, J.F. Colorectal Anastomotic Leakage: A New Experimental Model. J. Surg. Res. 2009, 155, 7–12. [Google Scholar] [CrossRef]
- Van der Ham, A.C.; Kort, W.J.; Weijma, I.M.; van den Ingh, H.F.G.M.; Jeekel, H. Healing of Ischemic Colonic Anastomosis: Fibrin Sealant Does Not Improve Wound Healing. Dis. Colon Rectum 1992, 35, 884–891. [Google Scholar] [CrossRef]
- Hem, A.; Eide, D.M.; Engh, E.; Smith, A. (Eds.) Laboratory Animal Science; Norwegian Veterinary School: Oslo, Norway, 2001; ISBN 82-7725-117-3. [Google Scholar]
- Van der Ham, A.C.; Kort, W.J.; Weijma, I.M.; Jeekel, H. Transient Protection of Incomplete Colonic Anastomoses with Fibrin Sealant: An Experimental Study in the Rat. J. Surg. Res. 1993, 55, 256–260. [Google Scholar] [CrossRef]
- Henne-Bruns, D.; Kreischer, H.P.; Schmiegelow, P.; Kremer, B. Reinforcement of Colon Anastomoses with Polyglycolic Acid Mesh: An Experimental Study. Eur. Surg. Res. 1990, 22, 224–230. [Google Scholar] [CrossRef]
- Shakhsheer, B.A.; Lec, B.; Zaborin, A.; Guyton, K.; Defnet, A.M.; Bagrodia, N.; Kandel, J.J.; Zaborina, O.; Hernandez, S.L.; Alverdy, J. Lack of Evidence for Tissue Hypoxia as a Contributing Factor in Anastomotic Leak Following Colon Anastomosis and Segmental Devascularization in Rats. Int. J. Color. Dis. 2017, 32, 539–547. [Google Scholar] [CrossRef]
- Gibiino, G.; Binda, C.; Cristofaro, L.; Sbrancia, M.; Coluccio, C.; Petraroli, C.; Jung, C.F.M.; Cucchetti, A.; Cavaliere, D.; Ercolani, G.; et al. Dysbiosis and Gastrointestinal Surgery: Current Insights and Future Research. Biomedicines 2022, 10, 2532. [Google Scholar] [CrossRef]
- Edomskis, P.; Goudberg, M.R.; Sparreboom, C.L.; Menon, A.G.; Wolthuis, A.M.; D’Hoore, A.; Lange, J.F. Matrix Metalloproteinase-9 in Relation to Patients with Complications after Colorectal Surgery: A Systematic Review. Int. J. Color. Dis. 2021, 36, 1–10. [Google Scholar] [CrossRef]
- Lam, A.; Keskey, R.; Alverdy, J. The Invisible Enemy: Gut Microbiota and Their Role in Anastomotic Leak. Semin. Colon Rectal Surg. 2022, 33, 100880. [Google Scholar] [CrossRef]
- Hyoju, S.K.; Adriaansens, C.; Wienholts, K.; Sharma, A.; Keskey, R.; Arnold, W.; van Dalen, D.; Gottel, N.; Hyman, N.; Zaborin, A.; et al. Low-Fat/High-Fibre Diet Prehabilitation Improves Anastomotic Healing via the Microbiome: An Experimental Model. Br. J. Surg. 2020, 107, 743–755. [Google Scholar] [CrossRef]
- Li, H.; Ni, Y.; Su, M.; Qiu, Y.; Zhou, M.; Qiu, M.; Zhao, A.; Zhao, L.; Jia, W. Pharmacometabonomic Phenotyping Reveals Different Responses to Xenobiotic Intervention in Rats. J. Proteome Res. 2007, 6, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Ivulic, D.; Rossello-Mora, R.; Viver, T.; Montero, D.A.; Vidal, S.; Aspee, F.; Hidalgo, H.; Vidal, R. Litter Management Strategies and Their Impact on the Environmental and Respiratory Microbiome Might Influence Health in Poultry. Microorganisms 2022, 10, 878. [Google Scholar] [CrossRef] [PubMed]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low Diversity Gut Microbiota Dysbiosis: Drivers, Functional Implications and Recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef]
- Collins, M.D.; Rodriguez Jovita, M.; Foster, G.; Sjödén, B.; Falsen, E. Characterization of a Gemella-like Organism from the Oral Cavity of a Dog: Description of Gemella palaticanis Sp. Nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 1523–1526. [Google Scholar] [CrossRef] [Green Version]
- Selçuk, N.; Esma, C.; Ahmet Rıza, Ş.; Selma, A. Evaluation of Cases with Gemella Infection: Cross-Sectional Study. J. Infect. Dis. Epidemiol. 2018, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Youssef, D.; Youssef, I.; Marroush, T.S.; Sharma, M. Gemella Endocarditis: A Case Report and a Review of the Literature. Avicenna J. Med. 2019, 09, 164–168. [Google Scholar] [CrossRef]
- Collins, M.D. The Genus Gemella. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 511–518. ISBN 978-0-387-25494-4. [Google Scholar]
- Damé-Teixeira, N.; Parolo, C.C.F.; Maltz, M.; Rup, A.G.; Devine, D.A.; Do, T. Gene Expression of Bacterial Collagenolytic Proteases in Root Caries. J. Oral Microbiol. 2018, 10, 1424475. [Google Scholar] [CrossRef] [Green Version]
- Ho, E.X.P.; Cheung, C.M.G.; Sim, S.; Chu, C.W.; Wilm, A.; Lin, C.B.; Mathur, R.; Wong, D.; Chan, C.M.; Bhagarva, M.; et al. Human Pharyngeal Microbiota in Age-Related Macular Degeneration. PLoS ONE 2018, 13, e0201768. [Google Scholar] [CrossRef] [Green Version]
- Ternes, D.; Tsenkova, M.; Pozdeev, V.I.; Meyers, M.; Koncina, E.; Atatri, S.; Schmitz, M.; Karta, J.; Schmoetten, M.; Heinken, A.; et al. The Gut Microbial Metabolite Formate Exacerbates Colorectal Cancer Progression. Nat. Metab. 2022, 4, 458–475. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Fung, A.M.Y.; Chiu, S.K.; Yung, R.W.H.; Yuen, K.Y. Gemella bacteraemia Characterised by 16S Ribosomal RNA Gene Sequencing. J. Clin. Pathol. 2003, 56, 690–693. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, B.; Wei, Y. New Understanding of Gut Microbiota and Colorectal Anastomosis Leak: A Collaborative Review of the Current Concepts. Front. Cell. Infect. Microbiol. 2022, 12, 1022603. [Google Scholar] [CrossRef]
- Speziale, P.; Pietrocola, G. The Multivalent Role of Fibronectin-Binding Proteins A and B (FnBPA and FnBPB) of Staphylococcus aureus in Host Infections. Front. Microbiol. 2020, 11, 2054. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, M.; Wang, M.; Zang, J.; Zhang, X.; Hang, T. Structural Insights into the Intermolecular Interaction of the Adhesin SdrC in the Pathogenicity of Staphylococcus aureus. Acta Crystallogr. F Struct. Biol. Commun. 2021, 77, 47–53. [Google Scholar] [CrossRef]
- Joseph, S.; Aduse-Opoku, J.; Hashim, A.; Hanski, E.; Streich, R.; Knowles, S.C.L.; Pedersen, A.B.; Wade, W.G.; Curtis, M.A. A 16S RRNA Gene and Draft Genome Database for the Murine Oral Bacterial Community. mSystems 2021, 6, e01222-20. [Google Scholar] [CrossRef]
- Komiyama, E.Y.; Lepesqueur, L.S.S.; Yassuda, C.G.; Samaranayake, L.P.; Parahitiyawa, N.B.; Balducci, I.; Koga-Ito, C.Y. Enterococcus Species in the Oral Cavity: Prevalence, Virulence Factors and Antimicrobial Susceptibility. PLoS ONE 2016, 11, e0163001. [Google Scholar] [CrossRef] [Green Version]
- Najafi, K.; Ganbarov, K.; Gholizadeh, P.; Tanomand, A.; Rezaee, M.A.; Mahmood, S.S.; Asgharzadeh, M.; Kafil, H.S. Oral Cavity Infection by Enterococcus faecalis: Virulence Factors and Pathogenesis. Rev. Med. Microbiol. 2020, 31, 51–60. [Google Scholar] [CrossRef]
- Takayasu, L.; Suda, W.; Takanashi, K.; Iioka, E.; Kurokawa, R.; Shindo, C.; Hattori, Y.; Yamashita, N.; Nishijima, S.; Oshima, K.; et al. Circadian Oscillations of Microbial and Functional Composition in the Human Salivary Microbiome. DNA Res. 2017, 24, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, S.; Zhang, K. Gemella bergeriae Endocarditis Diagnosed by Sequencing of RRNA Genes in Heart Valve Tissue. J. Clin. Microbiol. 2004, 42, 4897–4900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García López, E.; Martín-Galiano, A.J. The Versatility of Opportunistic Infections Caused by Gemella Isolates Is Supported by the Carriage of Virulence Factors From Multiple Origins. Front. Microbiol. 2020, 11, 524. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.V.; Nallapareddy, S.R.; Sillanpää, J.; Murray, B.E. Importance of the Collagen Adhesin Ace in Pathogenesis and Protection against Enterococcus faecalis Experimental Endocarditis. PLoS Pathog. 2010, 6, e1000716. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Thomas, V.C.; Narayanan, S.; Olson, S.; Fleming, S.D.; Hancock, L.E. Gelatinase Contributes to the Pathogenesis of Endocarditis Caused by Enterococcus faecalis. Infect. Immun. 2010, 78, 4936–4943. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, C.V.; Taddei, A.; Amedei, A. The Controversial Role of Enterococcus faecalis in Colorectal Cancer. Ther. Adv. Gastroenterol. 2018, 11, 175628481878360. [Google Scholar] [CrossRef] [Green Version]
- Dubin, K.; Pamer, E.G. Enterococci and Their Interactions with the Intestinal Microbiome. Microbiol. Spectr. 2017, 5, BAD-0014-2016. [Google Scholar] [CrossRef] [Green Version]
- Loftus, M.; Hassouneh, S.A.-D.; Yooseph, S. Bacterial Community Structure Alterations within the Colorectal Cancer Gut Microbiome. BMC Microbiol. 2021, 21, 98. [Google Scholar] [CrossRef]
- Villmones, H.C.; Haug, E.S.; Ulvestad, E.; Grude, N.; Stenstad, T.; Halland, A.; Kommedal, Ø. Species Level Description of the Human Ileal Bacterial Microbiota. Sci. Rep. 2018, 8, 4736. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamorano, D.; Ivulic, D.; Viver, T.; Morales, F.; López-Kostner, F.; Vidal, R.M. Microbiota Phenotype Promotes Anastomotic Leakage in a Model of Rats with Ischemic Colon Resection. Microorganisms 2023, 11, 680. https://doi.org/10.3390/microorganisms11030680
Zamorano D, Ivulic D, Viver T, Morales F, López-Kostner F, Vidal RM. Microbiota Phenotype Promotes Anastomotic Leakage in a Model of Rats with Ischemic Colon Resection. Microorganisms. 2023; 11(3):680. https://doi.org/10.3390/microorganisms11030680
Chicago/Turabian StyleZamorano, Diego, Dinka Ivulic, Tomeu Viver, Felipe Morales, Francisco López-Kostner, and Roberto M. Vidal. 2023. "Microbiota Phenotype Promotes Anastomotic Leakage in a Model of Rats with Ischemic Colon Resection" Microorganisms 11, no. 3: 680. https://doi.org/10.3390/microorganisms11030680