

The Microbiota-Dependent Worsening Effects of Melatonin on Gut Inflammation
 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Induction of Experimental Intestinal Inflammation
2.3. Control and Melatonin Treatments
2.4. Depletion of the Gut Microbiota
2.5. Evaluation of Clinical Disease Score
2.6. Euthanasia and Sample Collection
2.7. Intestinal Permeability Evaluation by FITC-Dextran
2.8. Total and Differential Leukocyte Counts
2.9. Indirect Quantification of Neutrophil and Macrophage Activities
2.10. Enzyme Immunoassay for Cytokine Measurement by ELISA
2.11. Immunophenotyping of Spleen Cells and Mesenteric Lymph Nodes
2.12. Analysis of the Gut Microbiota
2.13. Statistical Analysis
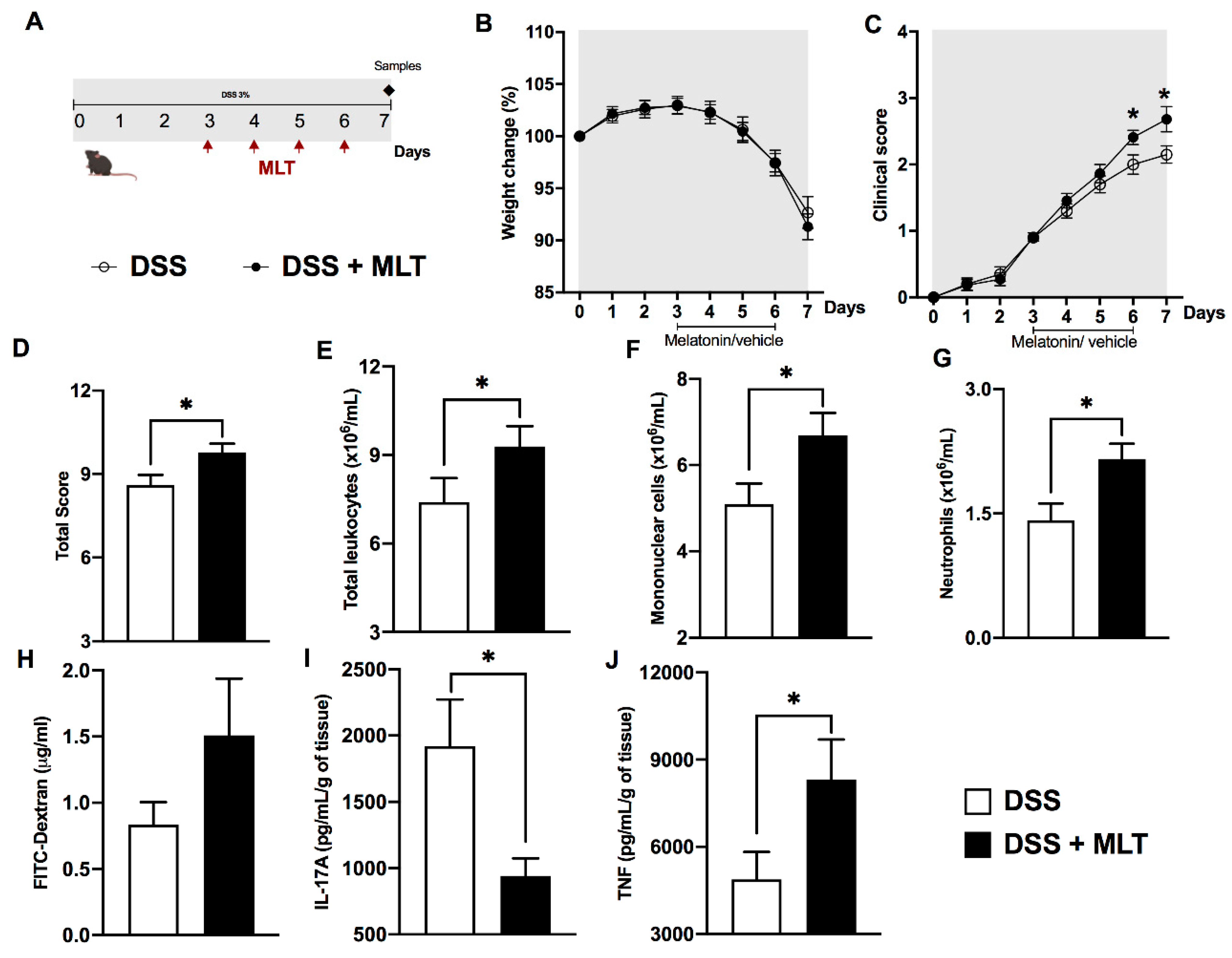
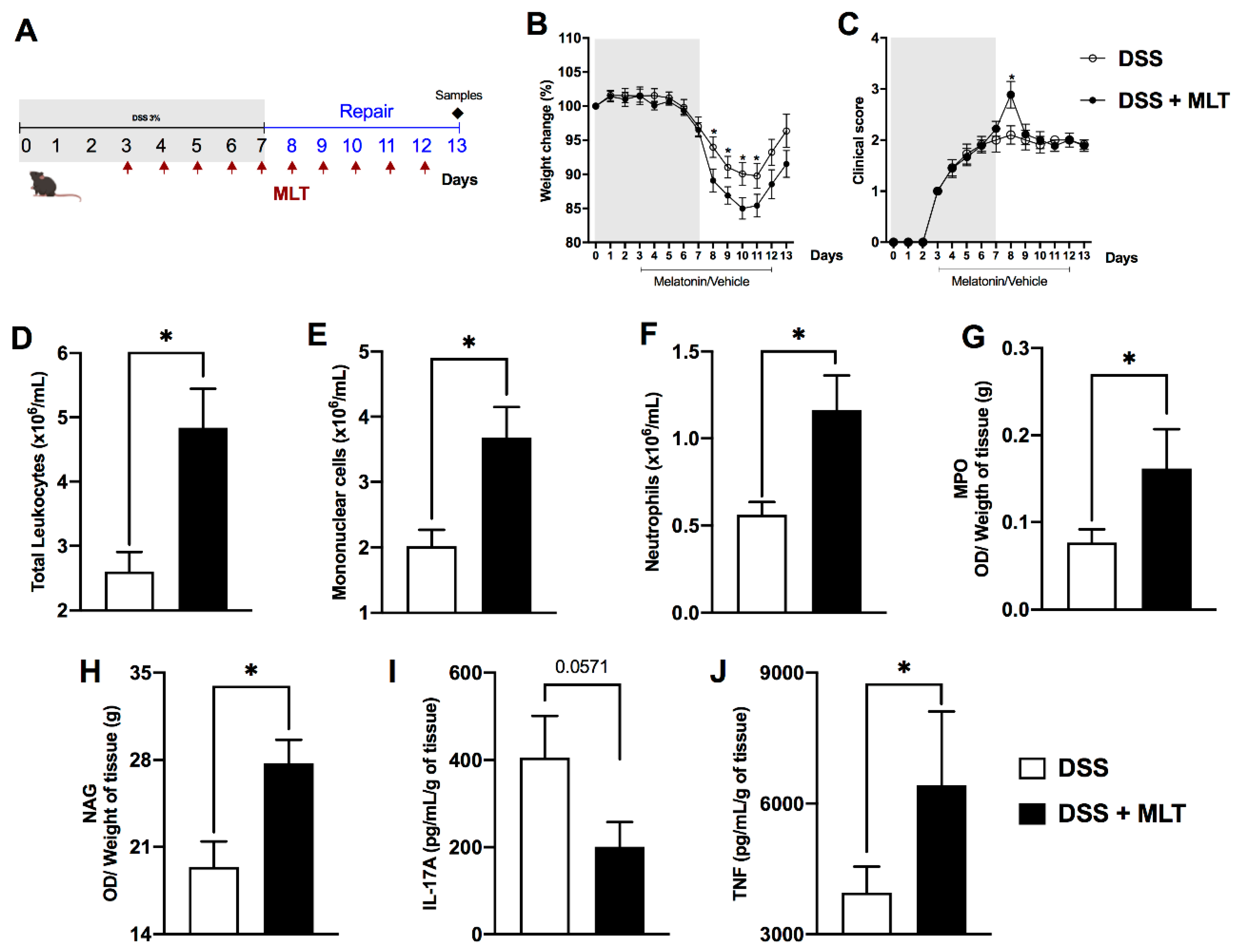
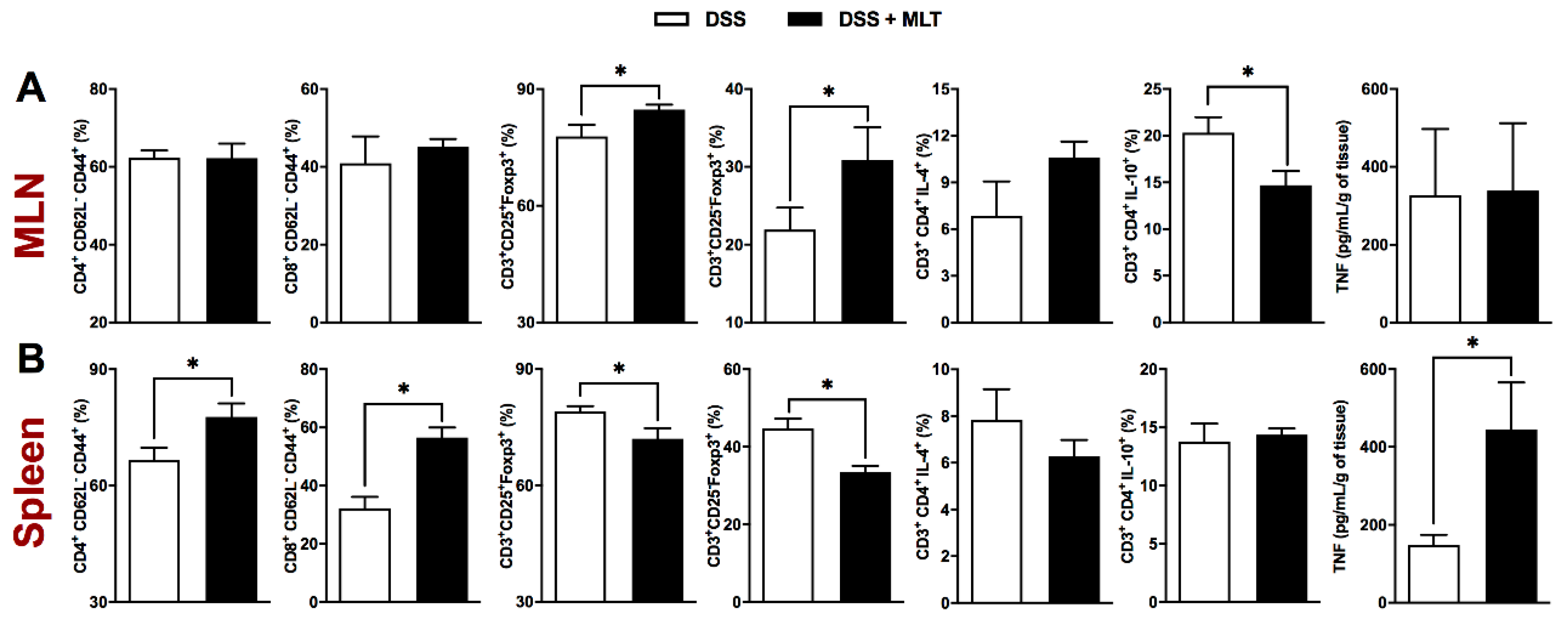
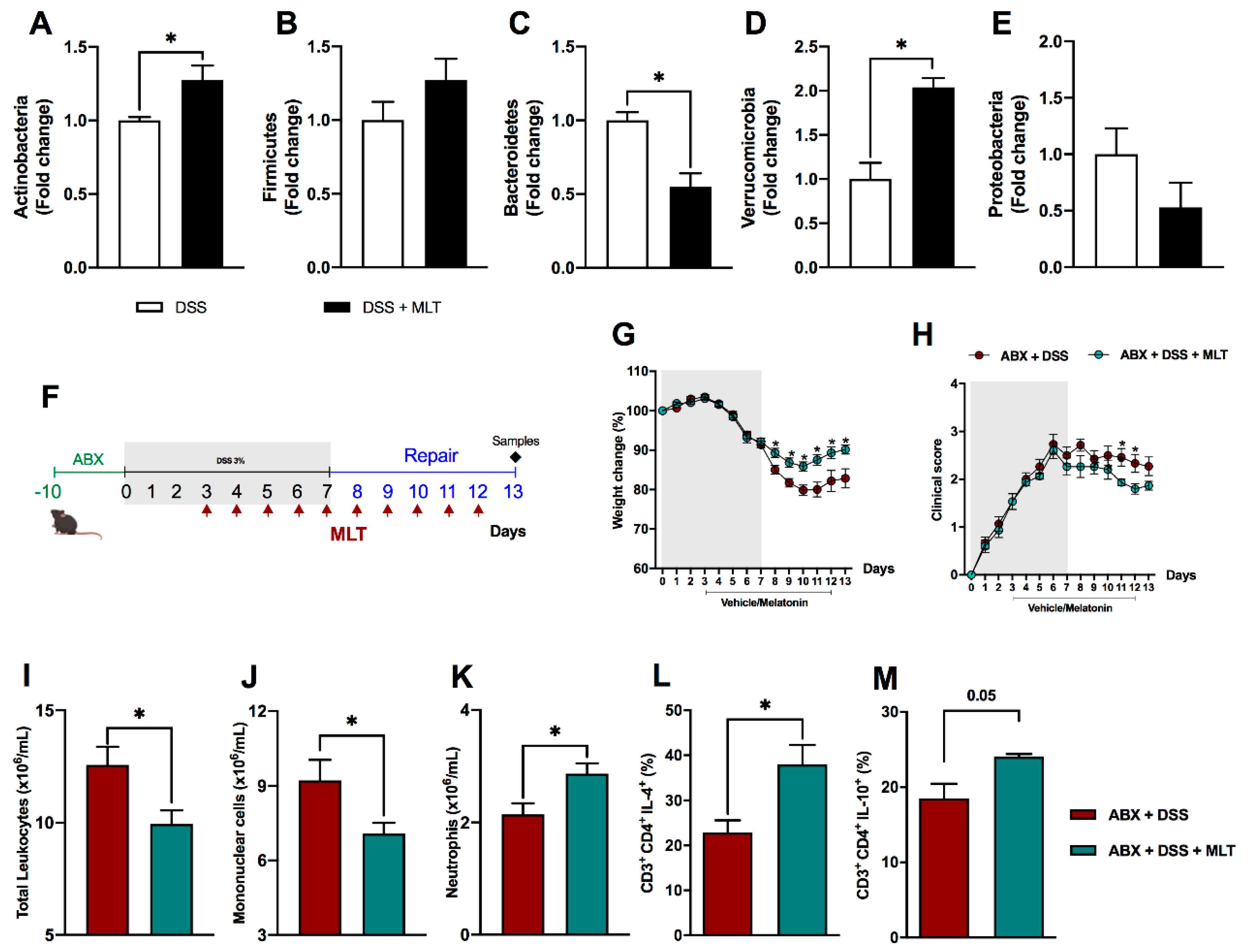
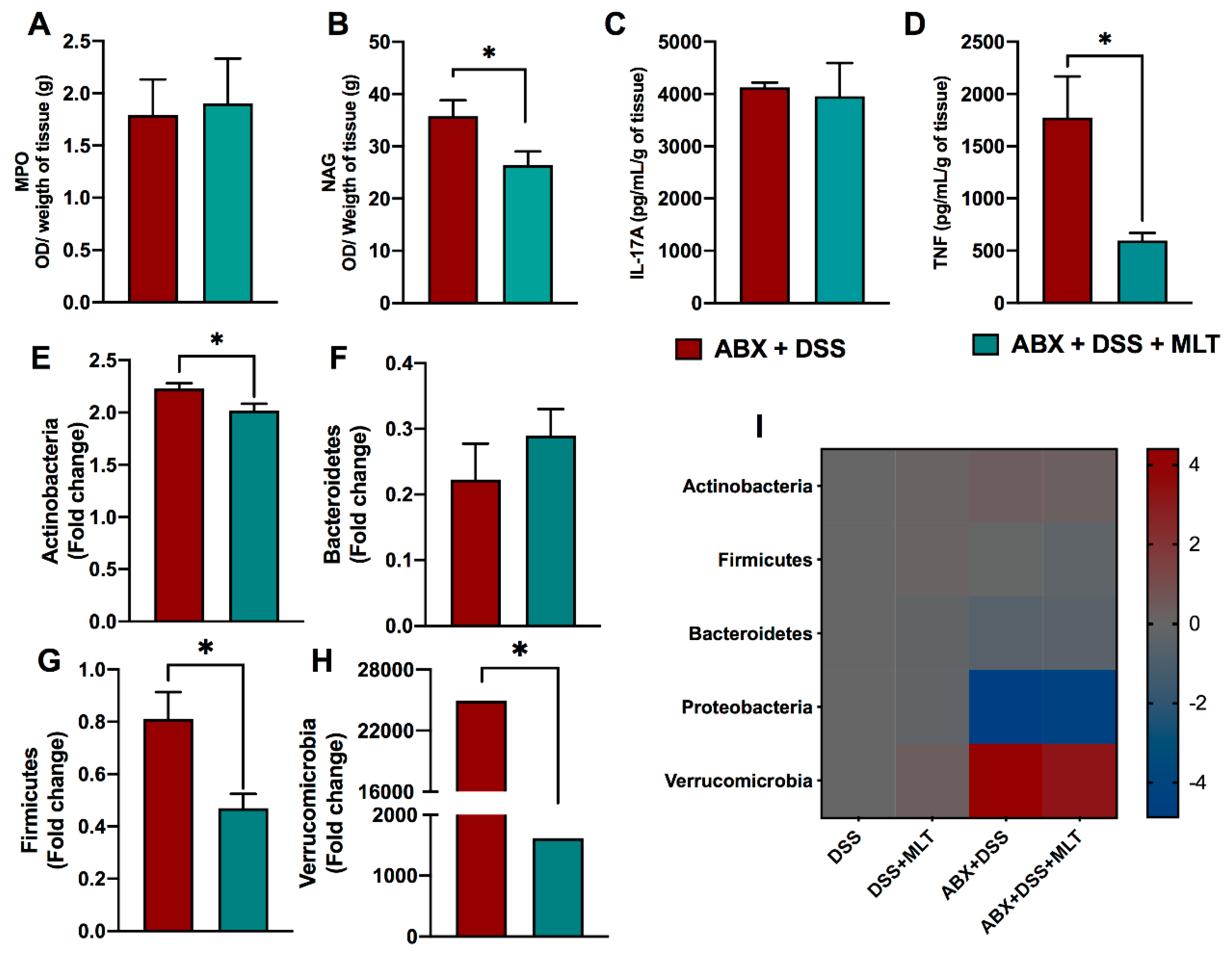
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Fazio, L.; Cavazza, E.; Spisni, E.; Strillacci, A.; Centanni, M.; Candela, M.; Praticò, C.; Campieri, M.; Ricci, C.; Valerii, M.C. Longitudinal analysis of inflammation and microbiota dynamics in a model of mild chronic dextran sulfate sodium-induced colitis in mice. World J. Gastroenterol. 2014, 20, 2051–2061. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef] [PubMed]
- Woelk, C.H.; Snyder, A. Modulating gut microbiota to treat cancer. Science 2021, 371, 573–574. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, L.; Chen, S.; Guo, S.; Yue, T.; Hou, Q.; Feng, M.; Xu, H.; Liu, Y.; Wang, P.; et al. The administration of Escherichia coli Nissle 1917 ameliorates irinotecan-induced intestinal barrier dysfunction and gut microbial dysbiosis in mice. Life Sci. 2019, 231, 116529. [Google Scholar] [CrossRef]
- Panpetch, W.; Hiengrach, P.; Nilgate, S.; Tumwasorn, S.; Somboonna, N.; Wilantho, A.; Chatthanathon, P.; Prueksapanich, P.; Leelahavanichkul, A. Additional Candida albicans administration enhances the severity of dextran sulfate solution induced colitis mouse model through leaky gut-enhanced systemic inflammation and gut-dysbiosis but attenuated by Lactobacillus rhamnosus L34. Gut Microbes 2020, 11, 465–480. [Google Scholar] [CrossRef]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef]
- Lee, J.W.J.; Plichta, D.; Hogstrom, L.; Borren, N.Z.; Lau, H.; Gregory, S.M.; Tan, W.; Khalili, H.; Clish, C.; Vlamakis, H.; et al. Multi-omics reveal microbial determinants impacting responses to biologic therapies in inflammatory bowel disease. Cell Host Microbe 2021, 29, 1294–1304.e4. [Google Scholar] [CrossRef]
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Marié, I.J.; Brambilla, L.; Azzouz, D.; Chen, Z.; Baracho, G.V.; Arnett, A.; Li, H.S.; Liu, W.; Cimmino, L.; Chattopadhyay, P.; et al. Tonic interferon restricts pathogenic IL-17-driven inflammatory disease via balancing the microbiome. eLife 2021, 10, e68371. [Google Scholar] [CrossRef]
- Kassouri, L.; Amiot, A.; Kirchgesner, J.; Tréton, X.; Allez, M.; Bouhnik, Y.; Beaugerie, L.; Carbonnel, F.; Meyer, A. The outcome of Crohn’s disease patients refractory to anti-TNF and either vedolizumab or ustekinumab. Dig. Liver Dis. 2020, 52, 1148–1155. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, T.; Cao, M.; Yuan, C.; Reiter, R.J.; Zhao, Z.; Zhao, Y.; Chen, L.; Fan, W.; Wang, X.; et al. Gut Microbiota Dysbiosis Induced by Decreasing Endogenous Melatonin Mediates the Pathogenesis of Alzheimer’s Disease and Obesity. Front. Immunol. 2022, 13, 900132. [Google Scholar] [CrossRef]
- Lin, R.; Wang, Z.; Cao, J.; Gao, T.; Dong, Y.; Chen, Y. Role of melatonin in murine “restraint stress”-induced dysfunction of colonic microbiota. J. Microbiol. 2021, 59, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Soták, M.; Mrnka, L.; Pácha, J. Heterogeneous expression of melatonin receptor MT1 mRNA in the rat intestine under control and fasting conditions. J. Pineal Res. 2006, 41, 183–188. [Google Scholar] [CrossRef]
- Wang, B.; Zhu, S.; Liu, Z.; Wei, H.; Zhang, L.; He, M.; Pei, F.; Zhang, J.; Sun, Q.; Duan, L. Increased Expression of Colonic Mucosal Melatonin in Patients with Irritable Bowel Syndrome Correlated with Gut Dysbiosis. Genomics Proteomics Bioinform. 2020, 18, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Q.; Fichna, J.; Bashashati, M.; Li, Y.-Y.; Storr, M. Distribution, function and physiological role of melatonin in the lower gut. World J. Gastroenterol. 2011, 17, 3888–3898. [Google Scholar] [CrossRef]
- Kvetnoy, I.M.; Ingel, I.E.; Kvetnaia, T.V.; Malinovskaya, N.K.; Rapoport, S.I.; Raikhlin, N.T.; Trofimov, A.V.; Yuzhakov, V.V. Gastrointestinal melatonin: Cellular identification and biological role. Neuroendocrinol. Lett 2002, 23, 121–132. [Google Scholar]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef]
- Paulose, J.K.; Cassone, V.M. The melatonin-sensitive circadian clock of the enteric bacterium Enterobacter aerogenes. Gut Microbes 2016, 7, 424–427. [Google Scholar] [CrossRef]
- Calvo, J.R.; Guerrero, J.M.; Osuna, C.; Molinero, P.; Carrillo-Vico, A. Melatonin triggers Crohn’s disease symptoms. J. Pineal Res. 2002, 32, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.P.; Jena, G.B. Melatonin reduces ulcerative colitis-associated local and systemic damage in mice: Investigation on possible mechanisms. Dig. Dis. Sci. 2013, 58, 3460–3474. [Google Scholar] [CrossRef]
- Song, T.-Y.; Lin, H.-C.; Chen, C.-L.; Wu, J.-H.; Liao, J.-W.; Hu, M.-L. Ergothioneine and melatonin attenuate oxidative stress and protect against learning and memory deficits in C57BL/6J mice treated with D-galactose. Free Radic. Res. 2014, 48, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Lin, L.; Li, H.; Wang, H.; Jiang, S.; Huang, P.; Lin, Q.; Chen, X.; Deng, Y. Melatonin Reduces Neuroinflammation and Improves Axonal Hypomyelination by Modulating M1/M2 Microglia Polarization via JAK2-STAT3-Telomerase Pathway in Postnatal Rats Exposed to Lipopolysaccharide. Mol. Neurobiol. 2021, 58, 6552–6576. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-M.; Zhao, C.-C.; Xie, Q.-M.; Xu, J.; Fei, G.-H. TLR2-Melatonin Feedback Loop Regulates the Activation of NLRP3 Inflammasome in Murine Allergic Airway Inflammation. Front. Immunol. 2020, 11, 172. [Google Scholar] [CrossRef]
- Sales-Campos, H.; de Souza, P.R.; Basso, P.J.; Ramos, A.D.; Nardini, V.; Chica, J.E.L.; Capurro, M.L.; Sá-Nunes, A.; de Barros Cardoso, C.R. Aedes aegypti salivary gland extract ameliorates experimental inflammatory bowel disease. Int. Immunopharmacol. 2015, 26, 13–22. [Google Scholar] [CrossRef]
- Basso, P.J.; Sales-Campos, H.; Nardini, V.; Duarte-Silva, M.; Alves, V.B.F.; Bonfá, G.; Rodrigues, C.C.; Ghirotto, B.; Chica, J.E.L.; Nomizo, A.; et al. Peroxisome Proliferator-Activated Receptor Alpha Mediates the Beneficial Effects of Atorvastatin in Experimental Colitis. Front. Immunol. 2021, 12, 618365. [Google Scholar] [CrossRef]
- Sales-Campos, H.; de Souza, P.R.; Basso, P.J.; Nardini, V.; Silva, A.; Banquieri, F.; Alves, V.B.F.; Chica, J.E.L.; Nomizo, A.; Cardoso, C.R.B. Amelioration of experimental colitis after short-term therapy with glucocorticoid and its relationship to the induction of different regulatory markers. Immunology 2017, 150, 115–126. [Google Scholar] [CrossRef]
- Leite, J.A.; Pessenda, G.; Guerra-Gomes, I.C.; de Santana, A.K.M.; André Pereira, C.; Ribeiro Campos Costa, F.; Ramos, S.G.; Simões Zamboni, D.; Caetano Faria, A.M.; Candido de Almeida, D.; et al. The DNA Sensor AIM2 Protects against Streptozotocin-Induced Type 1 Diabetes by Regulating Intestinal Homeostasis via the IL-18 Pathway. Cells 2020, 9, 959. [Google Scholar] [CrossRef]
- Ma, N.; Zhang, J.; Reiter, R.J.; Ma, X. Melatonin mediates mucosal immune cells, microbial metabolism, and rhythm crosstalk: A therapeutic target to reduce intestinal inflammation. Med. Res. Rev. 2020, 40, 606–632. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, Z.; Sun, H.; Song, J.; Chen, X.; Huang, J.; Lin, X.; Zhou, R. Effect of melatonin on T/B cell activation and immune regulation in pinealectomy mice. Life Sci. 2020, 242, 117191. [Google Scholar] [CrossRef] [PubMed]
- Pentney, P.T.; Bubenik, G.A. Melatonin reduces the severity of dextran-induced colitis in mice. J. Pineal Res. 1995, 19, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Jiang, Q.; Chen, S.; Fang, J.; Ren, W.; Yin, J.; Yao, K.; Yin, Y. Melatonin alters amino acid metabolism and inflammatory responses in colitis mice. Amino Acids 2017, 49, 2065–2071. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Mazzon, E.; Serraino, I.; Lepore, V.; Terranova, M.L.; Ciccolo, A.; Caputi, A.P. Melatonin reduces dinitrobenzene sulfonic acid-induced colitis. J. Pineal Res. 2001, 30, 1–12. [Google Scholar] [CrossRef]
- Mazzon, E.; Esposito, E.; Crisafulli, C.; Riccardi, L.; Muià, C.; Di Bella, P.; Meli, R.; Cuzzocrea, S. Melatonin modulates signal transduction pathways and apoptosis in experimental colitis. J. Pineal Res. 2006, 41, 363–373. [Google Scholar] [CrossRef]
- Maldonado, M.D.; Calvo, J.R. Melatonin usage in ulcerative colitis: A case report. J. Pineal Res. 2008, 45, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Marquez, E.; Sánchez-Fidalgo, S.; Calvo, J.R.; la de Lastra, C.A.; Motilva, V. Acutely administered melatonin is beneficial while chronic melatonin treatment aggravates the evolution of TNBS-induced colitis. J. Pineal Res. 2006, 40, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-X.; Yuan, X.; Cui, Y.-Y.; Liu, J.; Shen, J.; Jin, B.-Y.; Feng, B.-C.; Zhai, Y.-J.; Zheng, M.-Q.; Kou, G.-J.; et al. Melatonin Mitigates Oxazolone-Induced Colitis in Microbiota-Dependent Manner. Front. Immunol. 2021, 12, 783806. [Google Scholar] [CrossRef]
- Ma, F.; Hao, H.; Gao, X.; Cai, Y.; Zhou, J.; Liang, P.; Lv, J.; He, Q.; Shi, C.; Hu, D.; et al. Melatonin ameliorates necrotizing enterocolitis by preventing Th17/Treg imbalance through activation of the AMPK/SIRT1 pathway. Theranostics 2020, 10, 7730–7746. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, Z.; Hu, Y.; Li, Z.; Xie, Y.; Huang, H.; Chen, Q.; Chen, G.; Zhu, W.; Chen, Y.; et al. Melatonin, an endogenous hormone, modulates Th17 cells via the reactive-oxygen species/TXNIP/HIF-1α axis to alleviate autoimmune uveitis. J. Neuroinflammation 2022, 19, 124. [Google Scholar] [CrossRef]
- Marafini, I.; Sedda, S.; Dinallo, V.; Monteleone, G. Inflammatory cytokines: From discoveries to therapies in IBD. Expert Opin. Biol. Ther. 2019, 19, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.-R.; Bain, C.C.; Fenton, T.M.; Kelly, A.; Brown, S.L.; Ivens, A.C.; Travis, M.A.; Cook, P.C.; MacDonald, A.S. Dynamics of Colon Monocyte and Macrophage Activation During Colitis. Front. Immunol. 2018, 9, 2764. [Google Scholar] [CrossRef]
- Kühl, A.A.; Kakirman, H.; Janotta, M.; Dreher, S.; Cremer, P.; Pawlowski, N.N.; Loddenkemper, C.; Heimesaat, M.M.; Grollich, K.; Zeitz, M.; et al. Aggravation of different types of experimental colitis by depletion or adhesion blockade of neutrophils. Gastroenterology 2007, 133, 1882–1892. [Google Scholar] [CrossRef]
- Zhong, G.; Zhang, J.; Guo, Y.; Wang, Y.; Wu, M.; Ren, J.; Li, Y.; Zhang, X.; Zhou, B.; Zhao, W.; et al. IF1 inactivation attenuates experimental colitis through downregulation of neutrophil infiltration in colon mucosa. Int. Immunopharmacol. 2021, 99, 107980. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Yu, L.; Fang, L.; Yang, W.; Yu, T.; Miao, Y.; Chen, M.; Wu, K.; Chen, F.; Cong, Y.; et al. CD177+ neutrophils as functionally activated neutrophils negatively regulate IBD. Gut 2018, 67, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Bishu, S.; El Zaatari, M.; Hayashi, A.; Hou, G.; Bowers, N.; Kinnucan, J.; Manoogian, B.; Muza-Moons, M.; Zhang, M.; Grasberger, H.; et al. CD4+ Tissue-resident Memory T Cells Expand and Are a Major Source of Mucosal Tumour Necrosis Factor α in Active Crohn’s Disease. J. Crohn’s Colitis 2019, 13, 905–915. [Google Scholar] [CrossRef]
- Gebhardt, T.; Whitney, P.G.; Zaid, A.; Mackay, L.K.; Brooks, A.G.; Heath, W.R.; Carbone, F.R.; Mueller, S.N. Different patterns of peripheral migration by memory CD4+ and CD8+ T cells. Nature 2011, 477, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Tomita, T.; Kanai, T.; Nemoto, Y.; Fujii, T.; Nozaki, K.; Okamoto, R.; Tsuchiya, K.; Nakamura, T.; Sakamoto, N.; Totsuka, T.; et al. Colitogenic CD4+ effector-memory T cells actively recirculate in chronic colitic mice. Inflamm. Bowel Dis. 2008, 14, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Otsuka, S.; Wong, N.; Abbasi, A.; Gaida, M.M.; Fan, Y.; Meerzaman, D.; Ashwell, J.D. TNF plays a crucial role in inflammation by signaling via T cell TNFR2. Proc. Natl. Acad. Sci. USA 2021, 118, e2109972118. [Google Scholar] [CrossRef]
- Schreiber, S.; Ben-Horin, S.; Leszczyszyn, J.; Dudkowiak, R.; Lahat, A.; Gawdis-Wojnarska, B.; Pukitis, A.; Horynski, M.; Farkas, K.; Kierkus, J.; et al. Randomized Controlled Trial: Subcutaneous vs Intravenous Infliximab CT-P13 Maintenance in Inflammatory Bowel Disease. Gastroenterology 2021, 160, 2340–2353. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Lee, M.; Chang, E.B. Inflammatory Bowel Diseases (IBD) and the Microbiome-Searching the Crime Scene for Clues. Gastroenterology 2021, 160, 524–537. [Google Scholar] [CrossRef]
- Xu, X.; Ocansey, D.K.W.; Hang, S.; Wang, B.; Amoah, S.; Yi, C.; Zhang, X.; Liu, L.; Mao, F. The gut metagenomics and metabolomics signature in patients with inflammatory bowel disease. Gut Pathog. 2022, 14, 26. [Google Scholar] [CrossRef]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 573–584. [Google Scholar] [CrossRef]
- Vrakas, S.; Mountzouris, K.C.; Michalopoulos, G.; Karamanolis, G.; Papatheodoridis, G.; Tzathas, C.; Gazouli, M. Intestinal Bacteria Composition and Translocation of Bacteria in Inflammatory Bowel Disease. PLoS ONE 2017, 12, e0170034. [Google Scholar] [CrossRef]
- Crouch, L.I.; Liberato, M.V.; Urbanowicz, P.A.; Baslé, A.; Lamb, C.A.; Stewart, C.J.; Cooke, K.; Doona, M.; Needham, S.; Brady, R.R.; et al. Prominent members of the human gut microbiota express endo-acting O-glycanases to initiate mucin breakdown. Nat. Commun. 2020, 11, 4017. [Google Scholar] [CrossRef]
- Ganesh, B.P.; Klopfleisch, R.; Loh, G.; Blaut, M. Commensal Akkermansia muciniphila exacerbates gut inflammation in Salmonella Typhimurium-infected gnotobiotic mice. PLoS ONE 2013, 8, e74963. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lu, W.; Tian, F.; Zhao, J.; Zhang, H.; Hong, K.; Yu, L. Akkermansia muciniphila Exerts Strain-Specific Effects on DSS-Induced Ulcerative Colitis in Mice. Front. Cell. Infect. Microbiol. 2021, 11, 698914. [Google Scholar] [CrossRef]
- Seregin, S.S.; Golovchenko, N.; Schaf, B.; Chen, J.; Pudlo, N.A.; Mitchell, J.; Baxter, N.T.; Zhao, L.; Schloss, P.D.; Martens, E.C.; et al. NLRP6 Protects Il10-/- Mice from Colitis by Limiting Colonization of Akkermansia muciniphila. Cell Rep. 2017, 19, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wu, W.; Wang, Q.; Yang, L.; Bian, X.; Jiang, X.; Lv, L.; Yan, R.; Xia, J.; Han, S.; et al. The negative effect of Akkermansia muciniphila-mediated post-antibiotic reconstitution of the gut microbiota on the development of colitis-associated colorectal cancer in mice. Front. Microbiol. 2022, 13, 932047. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Lan, C.; Li, H.; Ouyang, Q.; Kong, F.; Wu, A.; Ren, Z.; Tian, G.; Cai, J.; Yu, B.; et al. Rational consideration of Akkermansia muciniphila targeting intestinal health: Advantages and challenges. NPJ Biofilms Microbiomes 2022, 8, 81. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences 5′–3′ | |
|---|---|---|
| Actinobacteria | Sense | TGTAGCGGTGGAATGCGC |
| Antisense | AATTAAGCCACATGCTCCGCT | |
| Bacteroidetes | Sense | GTTTAATTCGATGATACGCGAG |
| Antisense | TTAASCCGACACCTCACGG | |
| Firmicutes | Sense | ATGTGGTTTAATTCGAAGCA |
| Antisense | AGCTGACGACAACCATGCAC | |
| Verrucomicrobia | Sense | TCAKGTCAGTATGGCCCTTAT |
| Antisense | CAGTTTTYAGGATTTCCTCCGCC | |
| Eubacteria | Sense | ACTCCTACGGGAGGCAGCAGT |
| Antisense | ATTACCGCGGCTGCTGGC | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, J.L.; Barbosa, L.V.; Pinzan, C.F.; Nardini, V.; Brigo, I.S.; Sebastião, C.A.; Elias-Oliveira, J.; Brazão, V.; Júnior, J.C.d.P.; Carlos, D.; et al. The Microbiota-Dependent Worsening Effects of Melatonin on Gut Inflammation. Microorganisms 2023, 11, 460. https://doi.org/10.3390/microorganisms11020460
da Silva JL, Barbosa LV, Pinzan CF, Nardini V, Brigo IS, Sebastião CA, Elias-Oliveira J, Brazão V, Júnior JCdP, Carlos D, et al. The Microbiota-Dependent Worsening Effects of Melatonin on Gut Inflammation. Microorganisms. 2023; 11(2):460. https://doi.org/10.3390/microorganisms11020460
Chicago/Turabian Styleda Silva, Jefferson Luiz, Lia Vezenfard Barbosa, Camila Figueiredo Pinzan, Viviani Nardini, Irislene Simões Brigo, Cássia Aparecida Sebastião, Jefferson Elias-Oliveira, Vânia Brazão, José Clóvis do Prado Júnior, Daniela Carlos, and et al. 2023. "The Microbiota-Dependent Worsening Effects of Melatonin on Gut Inflammation" Microorganisms 11, no. 2: 460. https://doi.org/10.3390/microorganisms11020460