
Analysis of Industrial Bacillus Species as Potential Probiotics for Dietary Supplements
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria, Reagents and Equipment
2.2. Bacterial Isolation and Cultivation
2.3. Microscopic and Quantitative Morphological Analysis of Bacteria
2.4. MALDI-TOF Bacterial Species Identification
2.5. Antagonistic Activities
2.6. Antibiotics Susceptibility
2.7. TP-84 Bacteriophage Sensitivity Tests
2.8. Statistical Analysis
3. Results
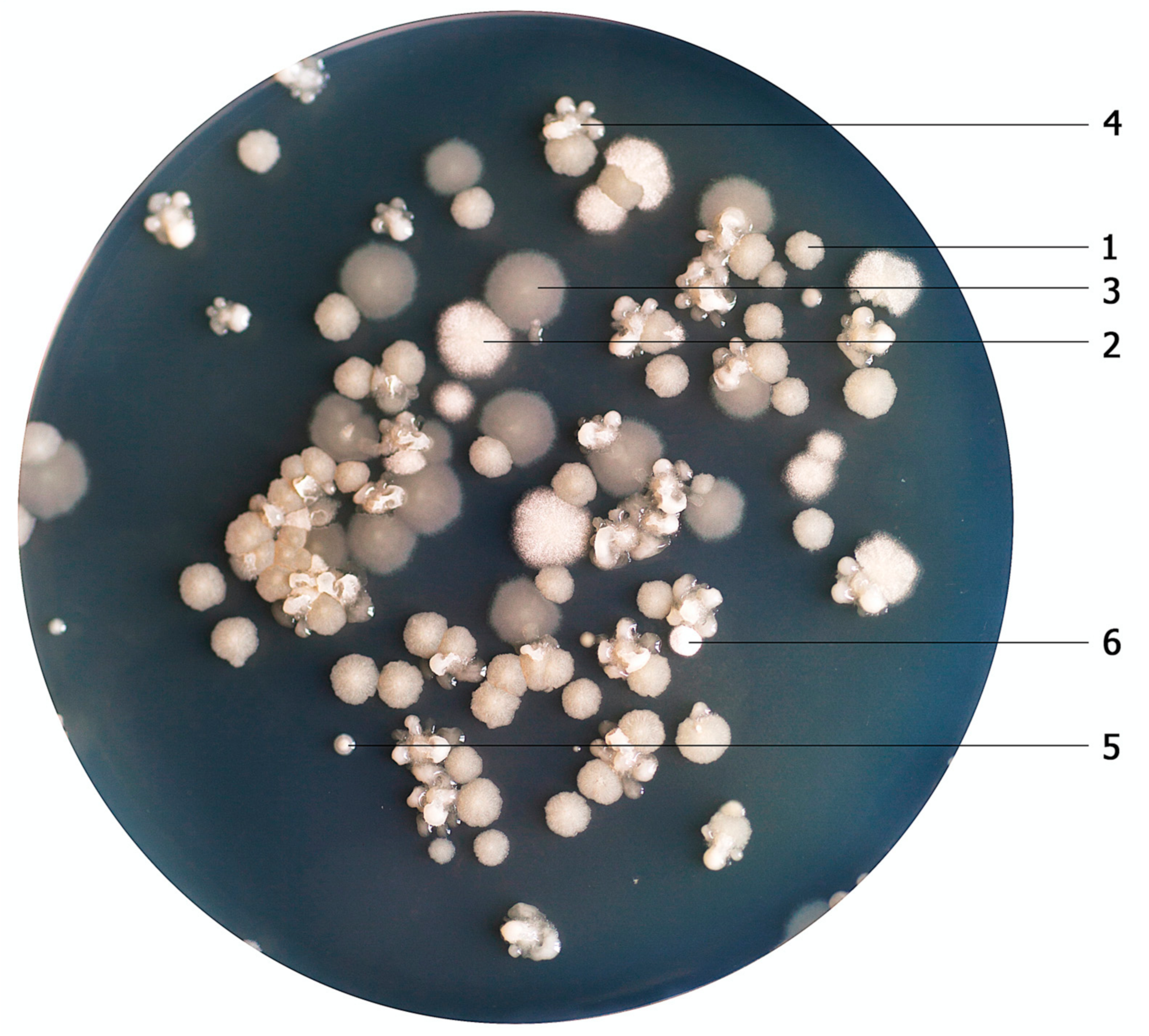
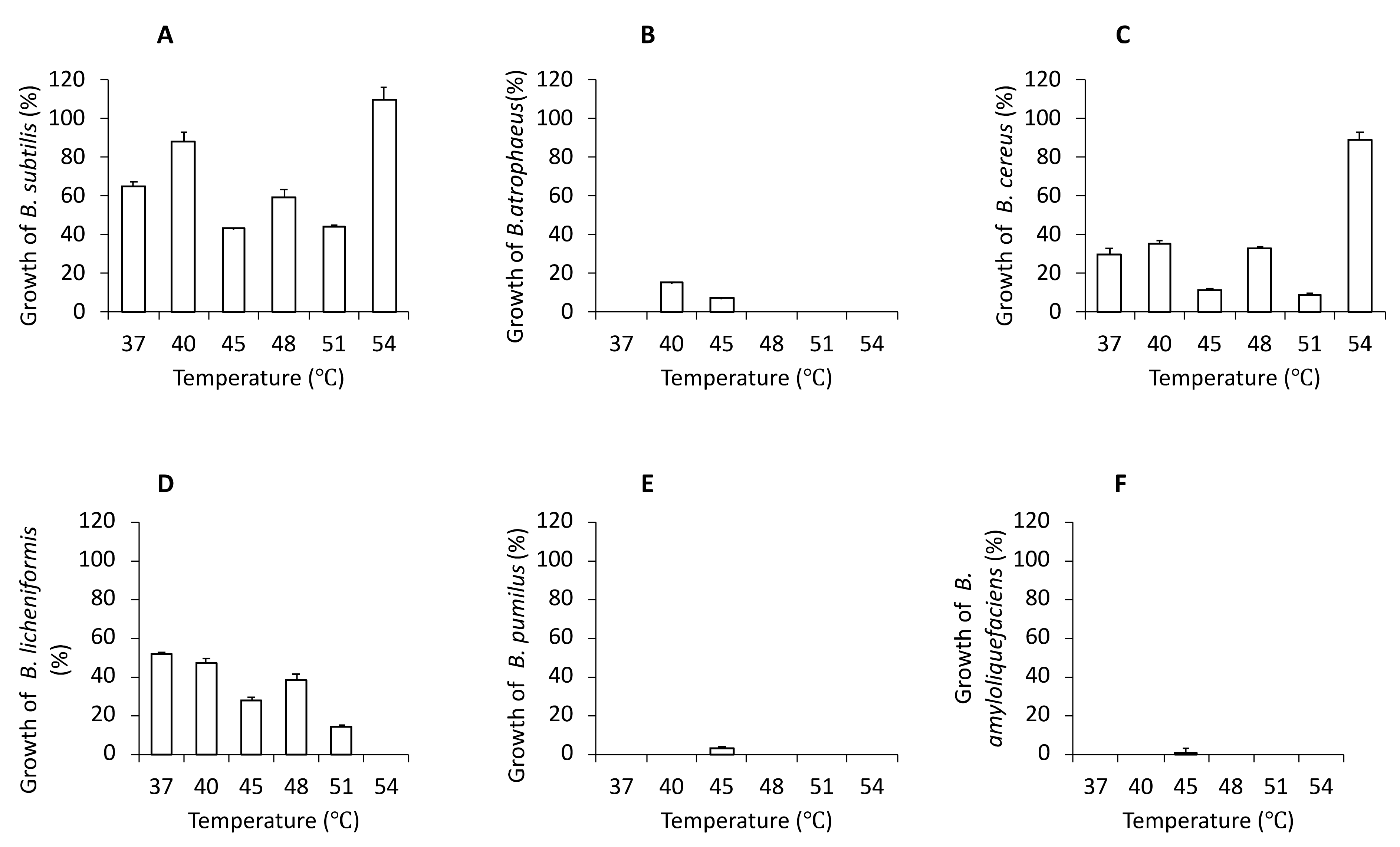
3.1. Selection and Quantification of Bacterial Cultures
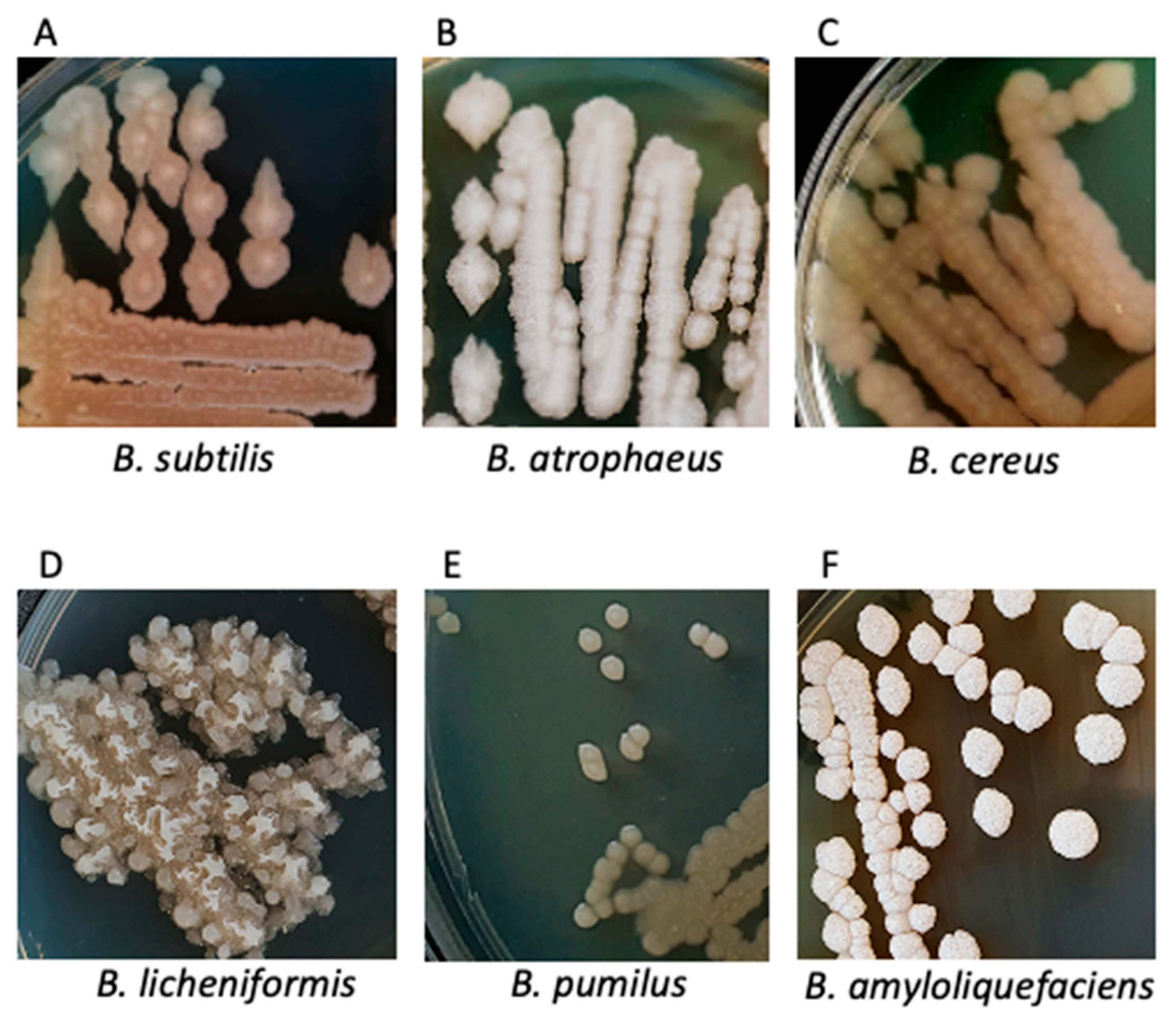
3.2. Characteristics of Bacterial Cultures, Colonies and Bacterial Cells
3.3. MALDI-TOF Mass Spectrometry Analysis of Bacteria
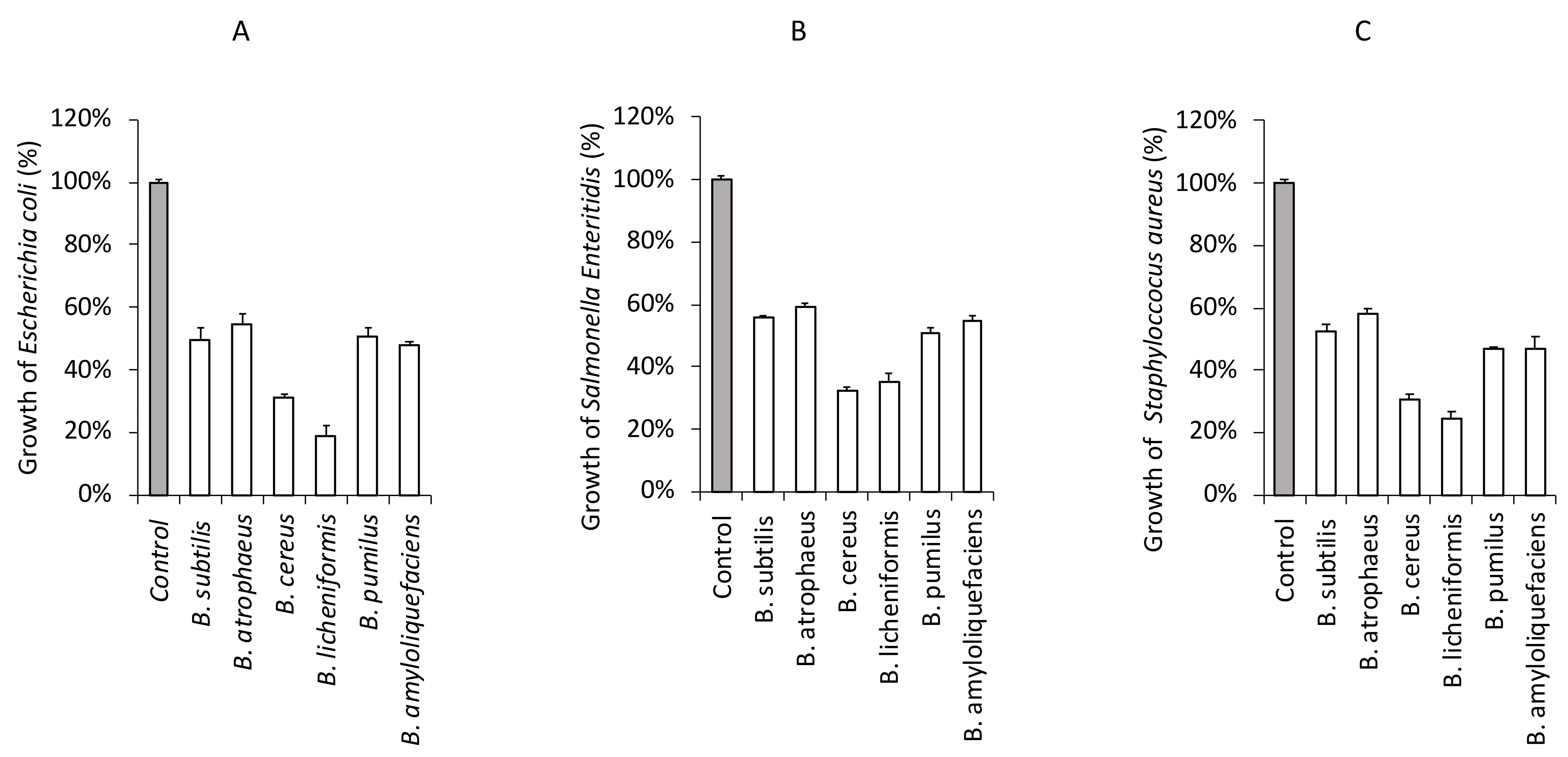
3.4. Antagonistic Effect of Foodborne Pathogenic Bacteria in the Presence of CFS from Bacillus Strains
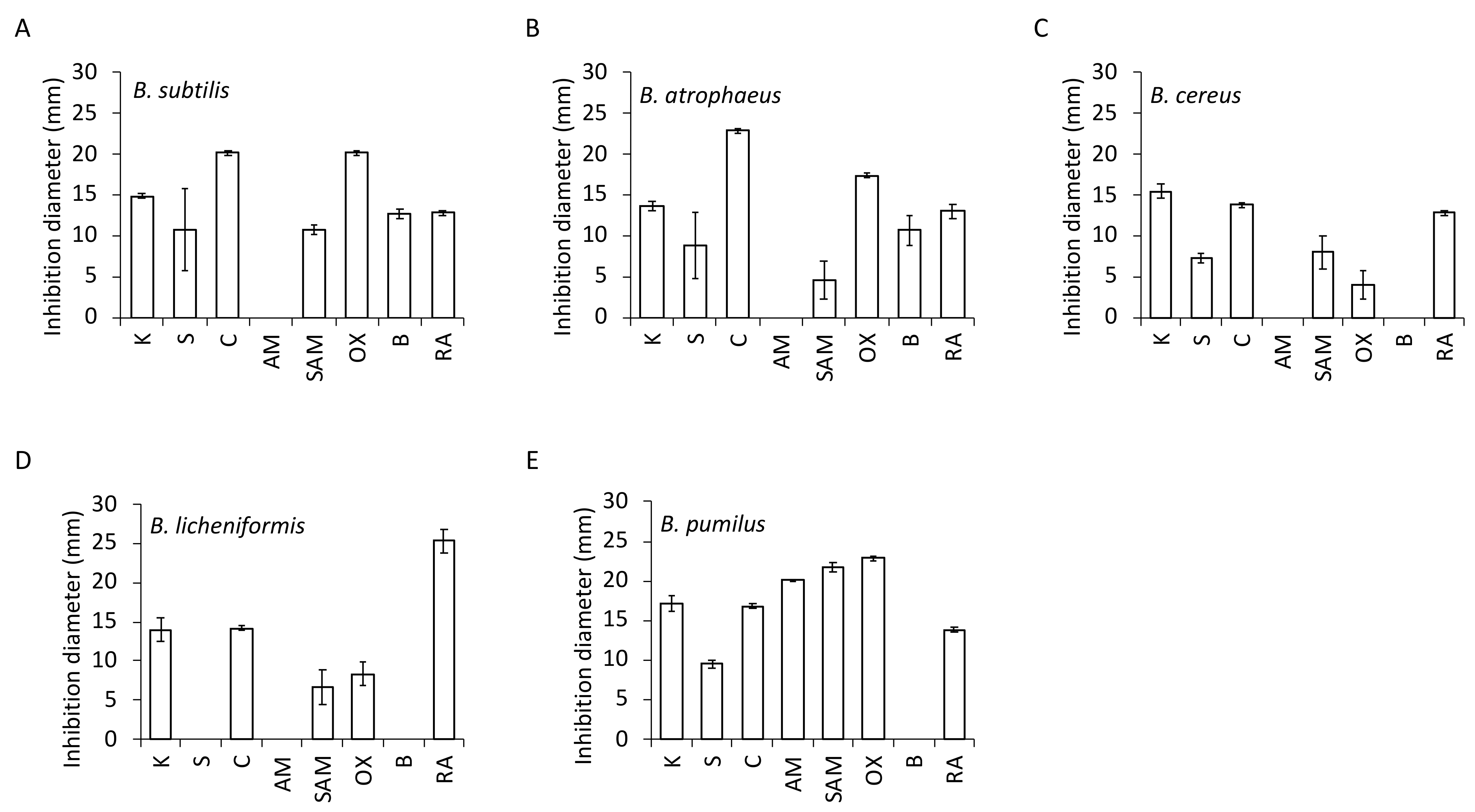
3.5. Antibiotic Resistance
3.6. TP-84 Bacteriophage Sensitivity Tests
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jeżewska-Frąckowiak, J.; Seroczyńska, K.; Banaszczyk, J.; Jedrzejczak, G.; Żylicz-Stachula, A.; Skowron, P.M. The promises and risks of probiotic Bacillus species. Acta Biochim. Pol. 2018, 65, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Gopikrishna, T.; Suresh Kumar, H.K.; Perumal, K.; Elangovan, E. Impact of Bacillus in fermented soybean foods on human health. Ann. Microbiol. 2021, 71, 30. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: A review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef] [PubMed]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef]
- Raul, D.; Biswas, T.; Mukhopadhyay, S.; Kumar Das, S.; Gupta, S. Production and Partial Purification of Alpha Amylase from Bacillus subtilis (MTCC 121) Using Solid State Fermentation. Biochem. Res. Int. 2014, 2014, 568141. [Google Scholar] [CrossRef]
- Jezewska-Frackowiak, J.; Seroczynska, K.; Banaszczyk, J.; Wozniak, D.; Skowron, M.; Ozog, A.; Zylicz-Stachula, A.; Ossowski, T.; Skowron, P.M. Detection of endospore producing Bacillus species from commercial probiotics and their preliminary microbiological characterization. J. Environ. Biol. 2017, 38, 1435–1440. [Google Scholar] [CrossRef]
- Yao, M.; Xie, J.; Du, H.; McClements, D.J.; Xiao, H.; Li, L. Progress in microencapsulation of probiotics: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 857–874. [Google Scholar] [CrossRef]
- Goderska, K. Different methods of probiotics stabilization. In Probiotics; Rigobelo, E., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Kiepś, J.; Dembczyński, R. Current Trends in the Production of Probiotic Formulations. Foods 2022, 11, 2330. [Google Scholar] [CrossRef]
- Šipailienė, A.; Petraitytė, S. Encapsulation of Probiotics: Proper Selection of the Probiotic Strain and the Influence of Encapsulation Technology and Materials on the Viability of Encapsulated Microorganisms. Probiotics Antimicrob. Proteins 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Mantzouridou, F.; Spanou, A.; Kiosseoglou, V. An inulin-based dressing emulsion as a potential probiotic food carrier. Food Res. Int. 2012, 46, 260–269. [Google Scholar] [CrossRef]
- Skowron, P.M.; Kropinski, A.M.; Zebrowska, J.; Janus, L.; Szemiako, K.; Czajkowska, E.; Maciejewska, N.; Skowron, M.; Łoś, J.; Łoś, M.; et al. Correction: Sequence, genome organization, annotation and proteomics of the thermophilic, 47.7-kb Geobacillus stearothermophilus bacteriophage TP-84 and its classification in the new Tp84virus genus. PLoS ONE 2018, 13, e0196798, Erratum in PLoS ONE 2018, 13, e0195449. [Google Scholar] [CrossRef]
- Skowron, P.M.; Krawczun, N.; Żebrowska, J.; Krefft, D.; Żołnierkiewicz, O.; Bielawa, M.; Jeżewska-Frąckowiak, J.; Janus, Ł.; Witkowska, M.; Palczewska, M.; et al. A vector-enzymatic DNA fragment amplification-expression technology for construction of artificial, concatemeric DNA, RNA and proteins for novel biomaterials, biomedical and industrial applications. Mater. Sci. Eng. C 2020, 108, 110426. [Google Scholar] [CrossRef]
- Available online: https://science.umd.edu/CBMG/faculty/asmith/BSCI223/lab3.pdf (accessed on 2 November 2022).
- Schaeffer, A.B.; Fulton, M.D. A Simplified Method of Staining Endospores. Science 1933, 77, 194. [Google Scholar] [CrossRef]
- Maneval, W.E. Staining bacteria and yeast with acid dyes. Stain Technol. 1941, 16, 13–19. [Google Scholar] [CrossRef]
- Kunicki–Goldfinger, W. The Life of Bacteria; PWN Scientific Publishing House: Warsaw, Poland, 1998; ISBN 978-83-01-14378-7. [Google Scholar]
- Jeżewska-Frąckowiak, J.; Żebrowska, J.; Czajkowska, E.; Jasińska, J.; Pęksa, M.; Jędrzejczak, G.; Skowron, P.M. Identification of bacterial species in probiotic consortiums in selected commercial cleaning preparations. Acta Biochim. Pol. 2019, 66, 215–222. [Google Scholar] [CrossRef]
- Wikins, T.D.; Holdeman, L.V.; Abramson, I.J.; Moore, W.E. Standardized single-disc method for antibiotic susceptibility testing of anaerobic bacteria. Antimicrob. Agents Chemother. 1972, 1, 451–459. [Google Scholar] [CrossRef]
- Berkeley, R.M.; Heyndrickx, N.L.; De Vos, P. Applications and Systematics of Bacillus and Relatives; Wiley-Blackwell: Oxford, UK, 2008. [Google Scholar]
- Kearns, D.B.; Losick, R. Cell population heterogeneity during growth of Bacillus subtilis. Genes 2005, 19, 3083–3094. [Google Scholar] [CrossRef]
- Lim, E.S.; Baek, S.Y.; Oh, T.; Koo, M.; Lee, J.Y.; Kim, H.J.; Kim, J.S. Strain variation in Bacillus cereus biofilms and their susceptibility to extracellular matrix-degrading enzymes. PLoS ONE 2021, 16, e0245708. [Google Scholar] [CrossRef]
- Liu, J.; He, D.; Li, X.Z.; Gao, S.; Wu, H.; Liu, W.; Gao, X.; Zhou, T. Gamma-polyglutamic acid (gamma-PGA) produced by Bacillus amyloliquefaciens C06 promoting its colonization on fruit surface. Int. J. Food Microbiol. 2010, 142, 190–197. [Google Scholar] [CrossRef]
- Azarko, J.; Wendt, U. Identyfikacja drobnoustrojów—Porównanie metody biochemicznej i spektrometrii masowej. Diagn. Lab. 2011, 47, 409–417. [Google Scholar]
- Kosikowska, P.; Pikula, M.; Langa, P.; Trzonkowski, P.; Obuchowski, M.; Lesner, A. Synthesis and Evaluation of Biological Activity of Antimicrobial--Pro-Proliferative Peptide Conjugates. PLoS ONE 2015, 10, e0140377. [Google Scholar] [CrossRef] [PubMed]
- Afrin, S.; Bhuiyan, M.N.I. Antagonistic activity of Bacillus amyloliquefaciens subspecies Amyloliquefaciens against multidrug resistant Serratia rubidaea. BioRxiv 2019. [CrossRef]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef]
- Huys, G.; D’Haene, K.; Swings, J. Influence of the culture medium on antibiotic susceptibility testing of food-associated lactic acid bacteria with the agar overlay disc diffusion method. Lett. Appl. Microbiol. 2002, 34, 402–406. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral potential of lactic acid bacteria and their bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef]
- Coppi, F.; Ruoppolo, M.; Mandressi, A.; Bellorofonte, C.; Gonnella, G.; Trinchieri, A. Results of treatment with Bacillus subtilis spores (Enterogermina) after antibiotic therapy in 95 patients with infection calculosis. Chemioterapia 1985, 4, 467–470. [Google Scholar]
- Bernardeau, M.; Lehtinen, M.J.; Forssten, S.D.; Nurminen, P. Importance of the gastrointestinal life cycle of Bacillus for probiotic functionality. J. Food Sci. Technol. 2017, 54, 2570–2584. [Google Scholar] [CrossRef]
- UK Standards for Microbiology Investigations. Identification of Bacillus species. In Bacteriology—Identification 9; Standards Unit, PHE, Microbiology Services; Public Health England: London, UK, 2015; pp. 1–27. [Google Scholar]
- Romo-Barrera, C.M.; Castrillón-Rivera, L.E.; Palma-Ramos, A.; Castañeda-Sánchez, J.I.; Luna-Herrera, J. Bacillus licheniformis and Bacillus subtilis, Probiotics That Induce the Formation of Macrophage Extracellular Traps. Microorganisms 2021, 9, 2027. [Google Scholar] [CrossRef]
- Hill, J.E.; Baiano, J.C.F.; Barnes, A.C. Isolation of a novel strain of “B. pumilus” from penaeid shrimp that is inhibitory against marine pathogens. J. Fish Dis. 2009, 32, 1007–1016. [Google Scholar] [CrossRef]
- Zakowska, D.; Bartoszcze, M.; Niemcewicz, M.; Bielawska-Drózd, A.; Kocik, J. New aspects of the infection mechanisms of Bacillus anthracis. Ann. Agric. Environ. Med. 2012, 19, 613–618. [Google Scholar]
- Pals, K.L.; Chang, R.T.; Ryan, A.J.; Gisolfi, C.V. Effect of running intensity on intestinal permeability. J. Appl. Physiol. 1985, 82, 571–576. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacillus Species | Antibiotics | |||||||
|---|---|---|---|---|---|---|---|---|
| K | S | C | AM | SAM | OX | B | RA | |
| B. subtilis | S | ND | S | R | S | S | S | S |
| B. atrophaeus | S * | ND * | S * | R * | ND * | S * | ND * | S * |
| B. cereus | ND | ND | S | R | ND | ND | R | S |
| B. licheniformis | ND * | R * | S * | R * | ND * | ND * | R * | ND * |
| B. pumilus | S * | S * | S * | S * | S * | S * | R * | S * |
| B. amyloliquefaciens | ND | ND | ND | ND | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łubkowska, B.; Jeżewska-Frąckowiak, J.; Sroczyński, M.; Dzitkowska-Zabielska, M.; Bojarczuk, A.; Skowron, P.M.; Cięszczyk, P. Analysis of Industrial Bacillus Species as Potential Probiotics for Dietary Supplements. Microorganisms 2023, 11, 488. https://doi.org/10.3390/microorganisms11020488
Łubkowska B, Jeżewska-Frąckowiak J, Sroczyński M, Dzitkowska-Zabielska M, Bojarczuk A, Skowron PM, Cięszczyk P. Analysis of Industrial Bacillus Species as Potential Probiotics for Dietary Supplements. Microorganisms. 2023; 11(2):488. https://doi.org/10.3390/microorganisms11020488
Chicago/Turabian StyleŁubkowska, Beata, Joanna Jeżewska-Frąckowiak, Michał Sroczyński, Magdalena Dzitkowska-Zabielska, Aleksandra Bojarczuk, Piotr M. Skowron, and Paweł Cięszczyk. 2023. "Analysis of Industrial Bacillus Species as Potential Probiotics for Dietary Supplements" Microorganisms 11, no. 2: 488. https://doi.org/10.3390/microorganisms11020488