
Geographic and Ecological Diversity of Green Sulfur Bacteria in Hot Spring Mat Communities
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Sample Collection
2.3. Light Microscopy
2.4. Enrichment Conditions
2.5. Isolation of GSB Strain MV4-Y
2.6. Chlorophyll and Carotenoid Analysis
2.7. Transmission Electron Microscopy
2.8. DNA Extraction and PCR Amplification
2.9. GSB Primer Design and PCR Optimization
2.10. PCR of Eubacterial 16S rRNA Genes
2.11. Cloning and Sequencing
2.12. Phylogenetic Analysis
2.13. Ecotype Simulation: Demarcation of Putative Ecotypes
2.14. Genome Comparison
3. Results
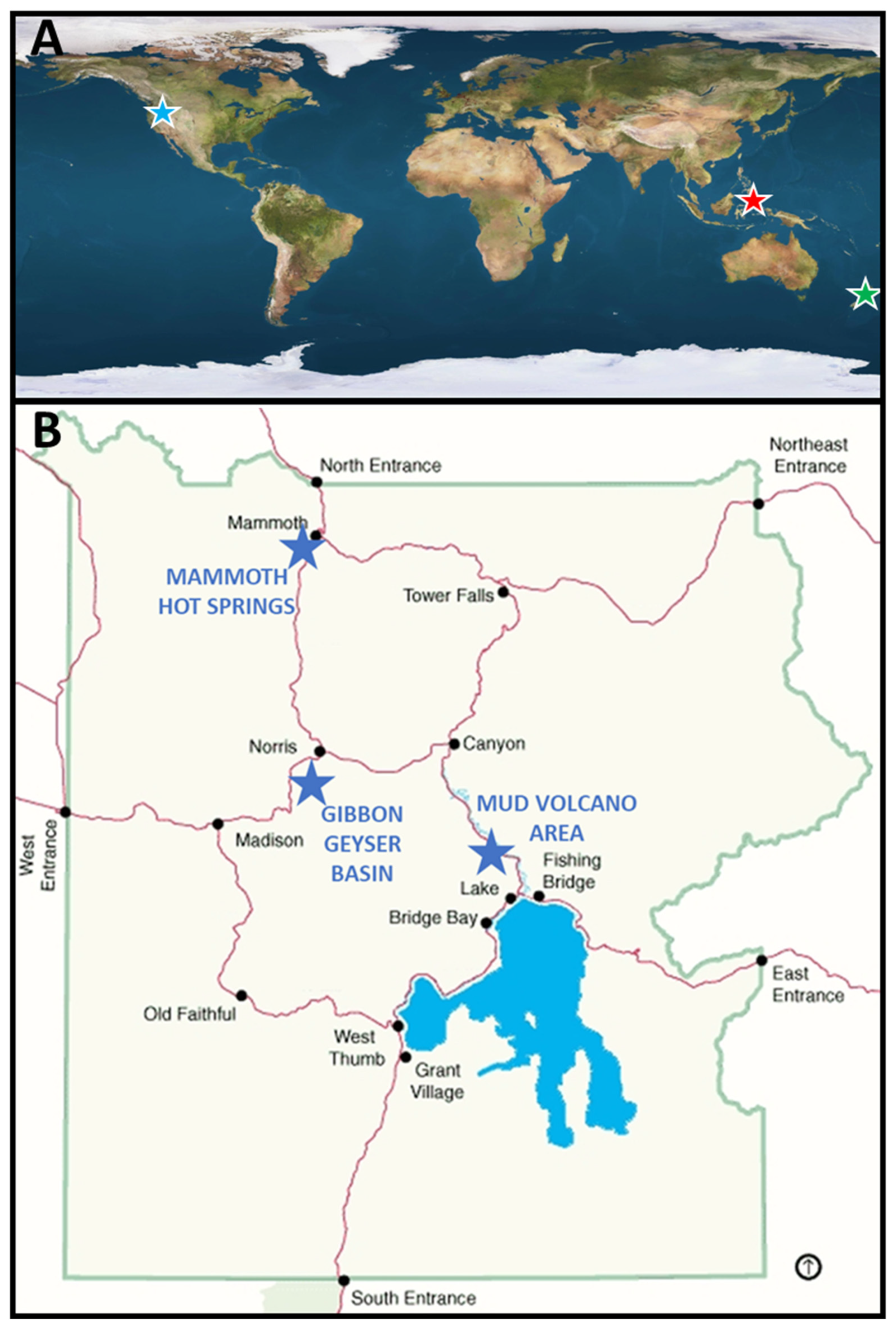
3.1. Site and Phototrophic Mat Descriptions
3.1.1. Travel Lodge Spring, Rotorua, NZ
3.1.2. Lipayo Hot Spring, Negros Island, PHL
3.1.3. Yellowstone National Park, WY, USA
Gibbon Geyser Basin
Mud Volcano Area
Mammoth Hot Springs
3.2. GSB Enrichments
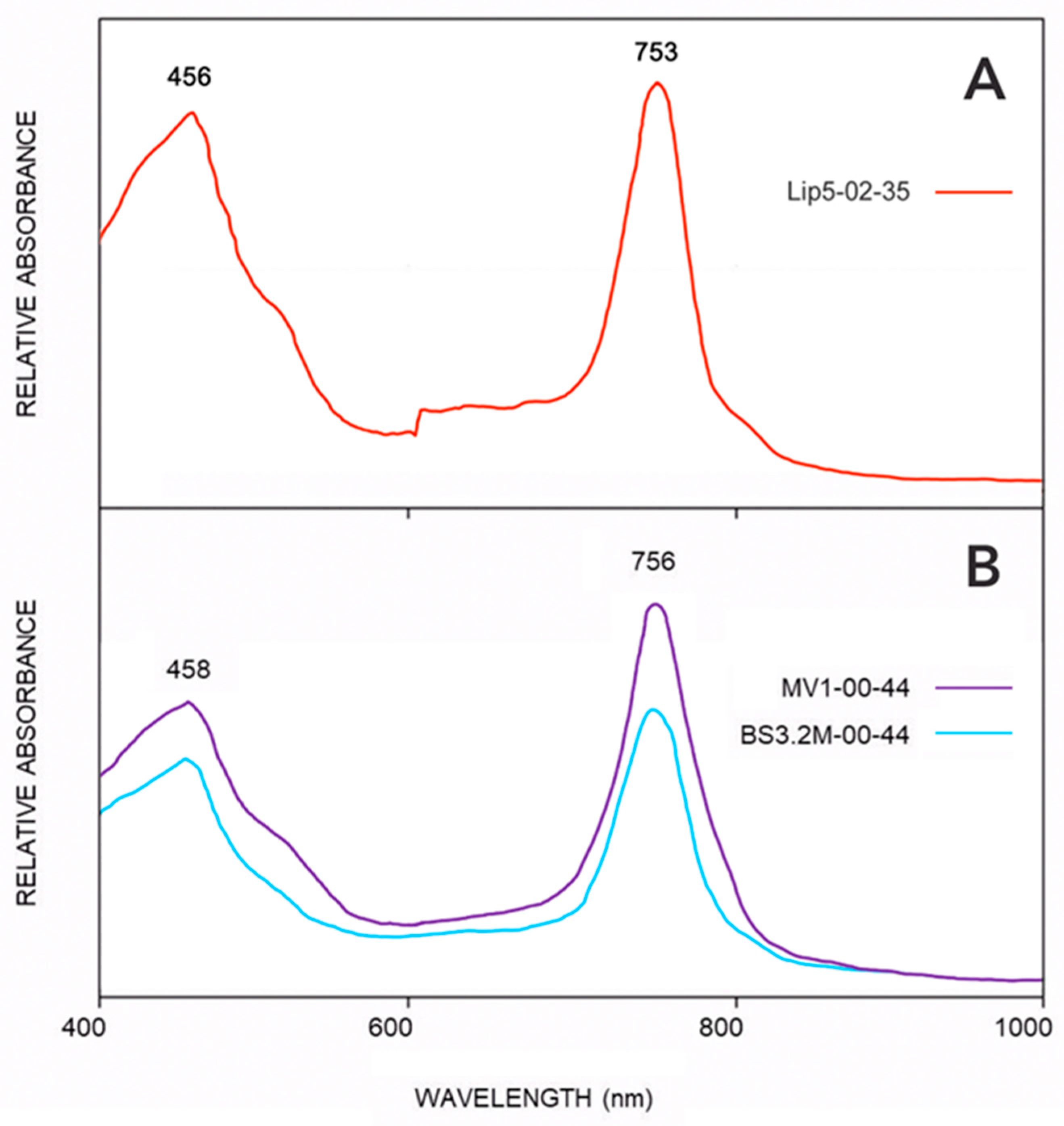
3.3. Absorption Spectra
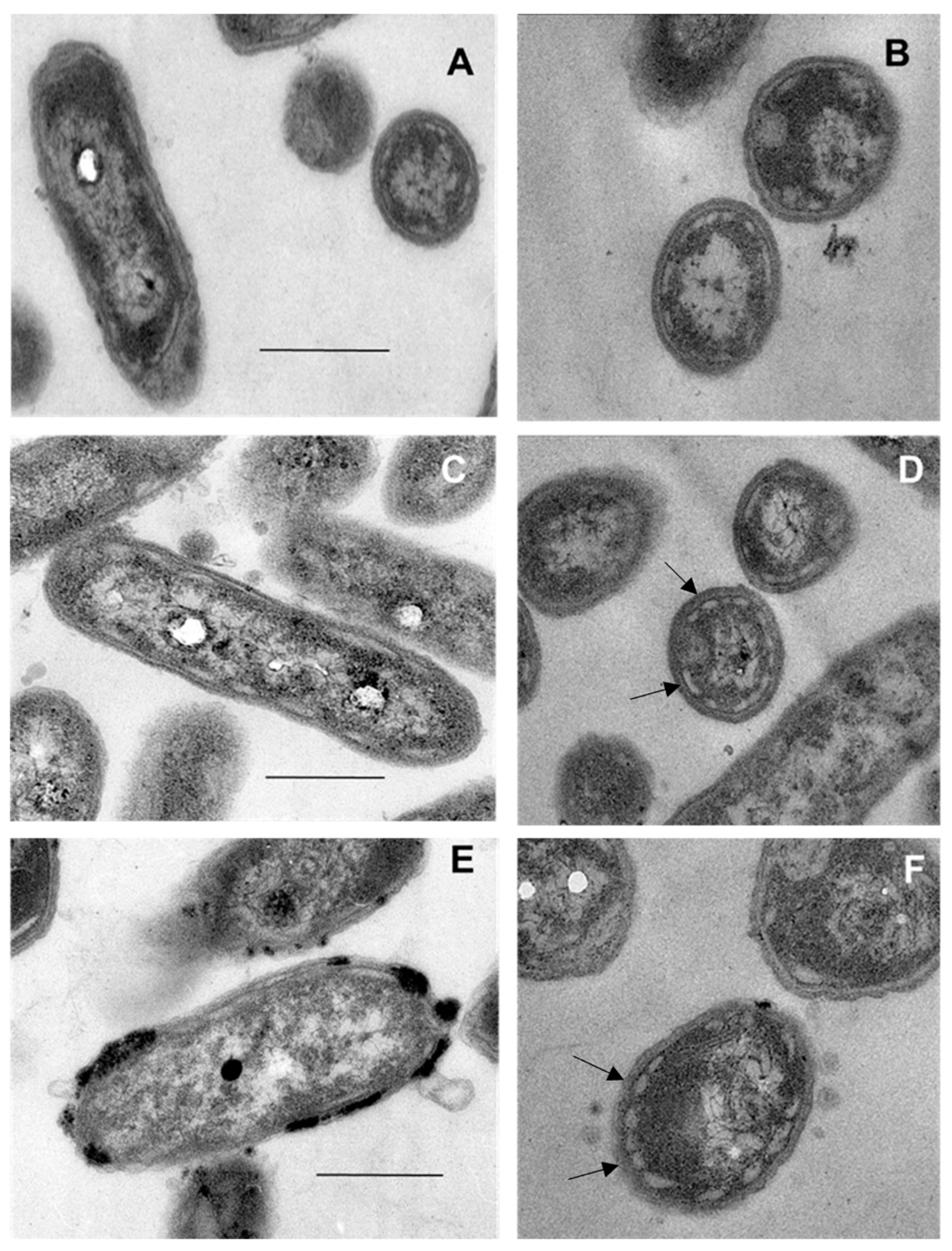
3.4. Transmission Electron Microscopy
3.5. Isolation of the Putative Moderately Thermophilic Chlorobaculum sp. Strain MV4-Y
3.6. Molecular Studies
3.6.1. 16S rRNA Sequences Amplified from Mat and GSB Enrichment DNA
3.6.2. Sequencing Results
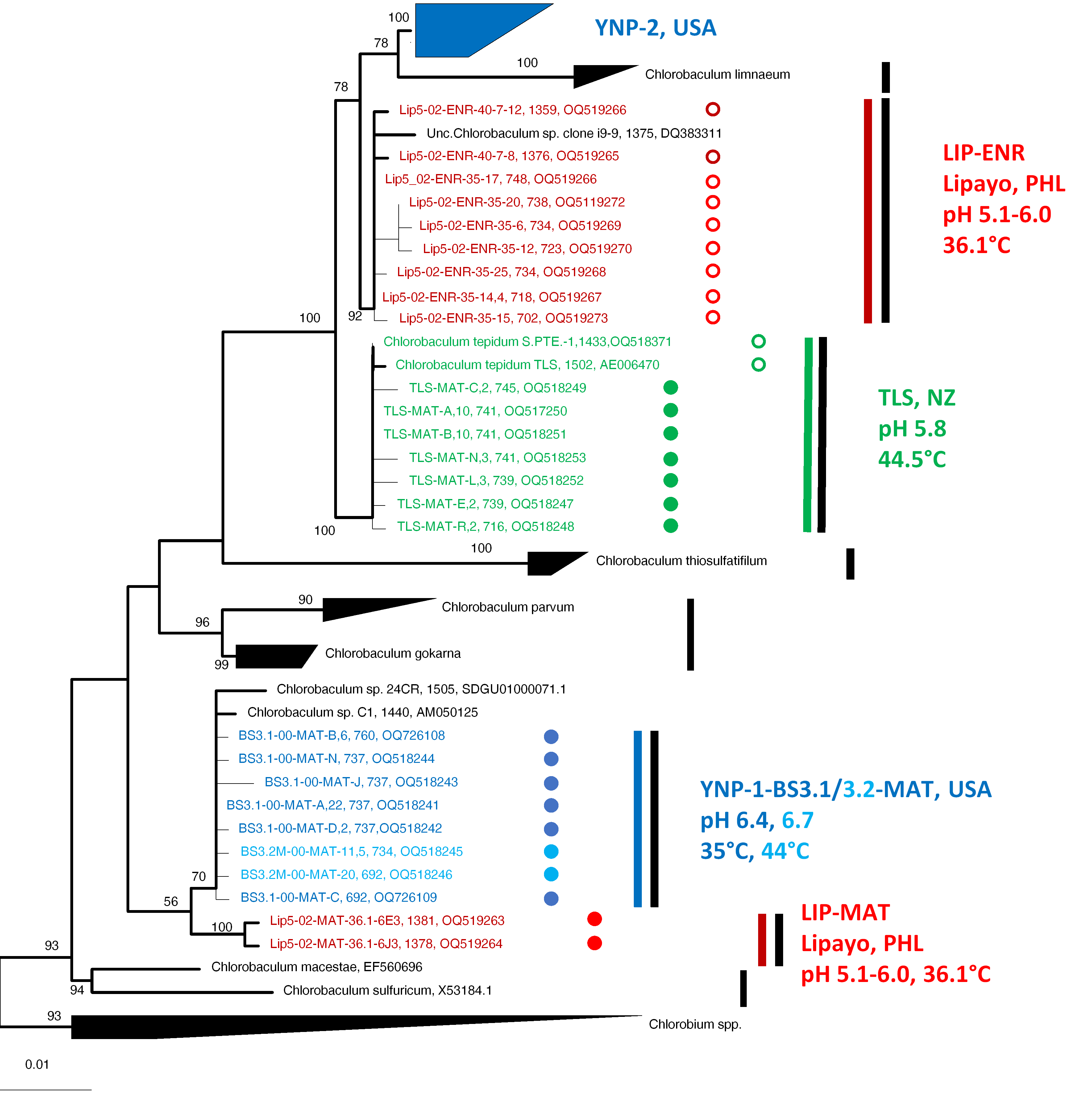
Phylogenetic Analyses
Correspondence between Clades, Subclades, and Environmental Conditions
Sequence Relatedness and Ecotype Simulation 2 Modeling
Enrichment Bias
4. Discussion
4.1. Putative Novel Chlorobaculum Species
4.2. Are the New GSB Species Thermophilic?
4.3. Cultivation Bias
4.4. Conditions in Which Hot Spring-Associated GSB May Be Found
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, G. Alternative pathways of carbon dioxide fixation: Insights into the early evolution of life? Ann. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Arnon, D.I. A reverse KREBS cycle in photosynthesis: Consensus at last. Photosynth. Res. 1990, 24, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Beh, M.; Strauss, G.; Huber, R.; Stetter, K.-O.; Fuchs, G. Enzymes of the reductive citric acid cycle in the autotrophic eubacterium Aquifex pyrophilus and in the archaebacterium Thermoproteus neutrophilus. Arch. Microbiol. 1993, 160, 306–311. [Google Scholar] [CrossRef]
- Ishii, M.; Igarashi, Y.; Kodama, T. Purification and isolation of ATP: Citrate lyase from Hydrogenobacter thermophilus TK-6. J. Bacteriol. 1989, 171, 1788–1792. [Google Scholar] [CrossRef]
- Shiba, H.; Kawasumi, T.; Igarashi, Y.; Kodama, T.; Minoda, Y. The CO2 assimilation via the reductive tricarboxylic acid cycle in an obligately autotrophic, aerobic hydrogen-oxidizing bacterium, Hydrogenobacter thermophilus. Arch. Microbiol. 1985, 141, 198–203. [Google Scholar] [CrossRef]
- Shima, S.; Suzuki, K.-I. Hydrogenobacter acidophilus sp. nov., a thermoacidophilic, aerobic, hydrogen-oxidizing bacterium requiring elemental sulfur for growth. Int. J. Syst. Evol. Microbiol. 1993, 43, 703–708. [Google Scholar] [CrossRef]
- Hohmann-Marriott, M.F.; Blankenship, R.E. Evolution of photosynthesis. Annu. Rev. Plant Biol. 2011, 62, 515–548. [Google Scholar] [CrossRef]
- Martin, W.F.; Bryant, D.A.; Beatty, J.T. A physiological perspective on the origin and evolution of photosynthesis. FEMS Microbiol. Rev. 2018, 42, 205–231. [Google Scholar] [CrossRef]
- Gisriel, C.J.; Azai, C.; Cardona, T. Recent advances in the structural diversity of reaction centers. Photosynth. Res. 2021, 149, 329–343. [Google Scholar] [CrossRef]
- Cui, X.; Liu, X.-L.; Shen, G.; Ma, J.; Husain, F.; Rocher, D.; Zumberge, J.E.; Bryant, D.A.; Summons, R.E. Niche expansion for phototrophic sulfur bacteria at the Proterozoic–Phanerozoic transition. Proc. Natl. Acad. Sci. USA 2020, 117, 17599–17606. [Google Scholar] [CrossRef]
- Ma, J.; French, K.L.; Cui, X.; Bryant, D.A.; Summons, R.E. Carotenoid biomarkers in Namibian shelf sediments: Anoxygenic photosynthesis during sulfide eruptions in the Benguela Upwelling System. Proc. Natl. Acad. Sci. USA 2021, 118, e2106040118. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.; Andersen, J.H.; Cox, R.P.; Imhoff, J.F. Phylogeny of green sulfur bacteria on the basis of gene sequences of 16S rRNA and of the Fenna-Matthews-Olson protein. Arch. Microbiol. 2002, 178, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. Phylogenetic taxonomy of the family Chlorobiaceae on the basis of 16S rRNA and fmo (Fenna-Matthews-Olson protein) gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Bello, S.; Howard-Azzeh, M.; Schellhorn, H.E.; Gupta, R.S. Phylogenomic analyses and molecular signatures elucidating the evolutionary relationships amongst the Chlorobia and Ignavibacteria species: Robust demarcation of two family-level clades within the order Chlorobiales and proposal for the family Chloroherpetonaceae fam. nov. Microorganisms 2022, 10, 1312. [Google Scholar]
- Gregersen, L.H.; Bryant, D.A.; Frigaard, N.-U. Mechanisms and evolution of oxidative sulfur metabolism in green sulfur bacteria. Front. Microbiol. 2011, 2, 116. [Google Scholar] [CrossRef]
- Trüper, H.G.; Pfennig, N. The Family Chlorobiaceae. In The Prokaryotes, 2nd ed.; Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.-H., Eds.; Springer: New York, NY, USA, 1992; Volume IV, pp. 3583–3592. [Google Scholar]
- Overmann, J. The green sulfur bacteria. In Bergey’s Manual of Determinative Bacteriology, 2nd ed.; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; Volume 1, pp. 601–620. [Google Scholar]
- Imhoff, J.F. The family Chlorobiaceae. In The Prokaryotes, 4th ed.; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin, Germany, 2014; pp. 501–514. [Google Scholar]
- Castenholz, R.W.; Bauld, J.; Jørgenson, B.B. Anoxygenic microbial mats of hot springs: Thermophilic Chlorobium sp. FEMS Microbiol. Ecol. 1990, 74, 325–336. [Google Scholar] [CrossRef]
- Wahlund, T.M.; Woese, C.R.; Castenholz, R.W.; Madigan, M.T. A thermophilic green sulfur bacterium from New Zealand hot springs, Chlorobium tepidum sp. nov. Arch. Microbiol. 1991, 156, 81–90. [Google Scholar] [CrossRef]
- Castenholz, R.W. The green sulfur and nonsulfur bacteria of hot springs. In Green Photosynthetic Bacteria; Olson, J.M., Ormerod, J.G., Ames, Z.J., Stackebrandt, E., Trüper, H.G., Eds.; Plenum Press: New York, NY, USA, 1988; pp. 243–255. [Google Scholar]
- Castenholz, R.W. The effect of sulfide on the blue-green algae of Hot Springs II. Yellowstone National Park. Microb. Ecol. 1977, 3, 79–105. [Google Scholar] [CrossRef]
- Madigan, M.T. A novel photosynthetic purple bacterium isolated from a Yellowstone hot spring. Science 1984, 225, 313–315. [Google Scholar] [CrossRef]
- Klatt, C.G.; Inskeep, W.P.; Herrgard, M.; Jay, Z.J.; Rusch, D.B.; Tringe, S.G.; Parenteau, M.N.; Ward, D.M.; Boomer, S.M.; Bryant, D.A.; et al. Community structure and function of high-temperature phototrophic microbial mats inhabiting diverse geothermal environments. Front. Microbiol. 2013, 4, 106. [Google Scholar] [CrossRef]
- Ward, D.M.; Ferris, M.J.; Nold, S.C.; Bateson, M.M. A natural view of microbial biodiversity within hot spring cyanobacterial mat communities. Microbiol. Mol. Biol. Rev. 1998, 62, 1353–1370. [Google Scholar] [CrossRef]
- Liu, Z.; Klatt, C.G.; Ludwig, M.; Rusch, D.B.; Jensen, S.I.; Kühl, M.; Ward, D.M.; Bryant, D.A. “Candidatus Thermochlorobacter aerophilum”: An aerobic chlorophotoheterotrophic member of the phylum Chlorobi. ISME J. 2012, 6, 1869–1882. [Google Scholar] [CrossRef]
- Koeppel, A.; Perry, E.B.; Sikorski, J.; Krizanc, D.; Warner, A.; Ward, D.M.; Rooney, A.P.; Brambilla, E.; Connor, N.; Nevo, E.; et al. Identifying the fundamental units of bacterial diversity: A paradigm shift to incorporate ecological diversification into bacterial systematics. Proc. Natl. Acad. Sci. USA 2008, 105, 2504–2509. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Eichler, B.; Pfennig, N. A new green sulfur bacterium from a freshwater pond. In Green Photosynthetic Bacteria; Olson, J.M., Ormerod, J.G., Amesz, J., Stackebrandt, E., Trüper, H.G., Eds.; Plenum Press: London, UK, 1988; pp. 233–235. [Google Scholar]
- Pfennig, N.; Trüper, H.G. Anoxygenic phototrophic bacteria. In Bergey’s Manual of Systematic Bacteriology; Staley, J.T., Bryant, M.P., Pfennig, N., Holt, J.G., Eds.; Williams & Wilking: Baltimore, MD, USA, 1989; Volume 3, pp. 1635–1709. [Google Scholar]
- Loeffler, F.E.; Sanford, R.A.; Ritalahti, K.M. Enrichment, cultivation, and detection of reductively dechlorinating bacteria. Meth. Enzymol. 2005, 397, 77–111. [Google Scholar]
- Robertson, N.L.; Carroll, T.W. Electron microscopy of the novel barley yellow streak mosaic virus. J. Ultrastruc. Mol. Struct. Res. 1989, 102, 139–146. [Google Scholar] [CrossRef]
- Nübel, U.; Bateson, M.M.; Vandieken, V.; Wieland, A.; Kühl, M.; Ward, D.M. Microscopic examination of distribution and phe-notypic properties of phylogenetically diverse Chloroflexaceae-related bacteria in hot spring microbial mats. Appl. Environ. Microbiol. 2002, 68, 4593–4603. [Google Scholar] [CrossRef]
- Van der Peer, Y.; Chapelle, S.; De Wachter, R. A quantitative map of nucleotide substitution rates in bacterial rRNA. Nucleic Acid Res. 1996, 24, 3381–3391. [Google Scholar] [CrossRef]
- Achenbach, L.A.; Carey, J.; Madigan, M.T. Photosynthetic and phylogenetic primers for detection of anoxygenic phototrophs in natural environments. Appl. Environ. Microbiol. 2001, 67, 2922–2926. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Lane, D.J.; Pace, B.; Olsen, G.J.; Stahl, D.A.; Sogin, M.L.; Pace, N.R. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analysis. Proc. Natl. Acad. Sci. USA. 1991, 82, 6955–6959. [Google Scholar] [CrossRef]
- Zheng, D.; Alm, E.W.; A Stahl, D.; Raskin, L. Characterization of universal small-subunit rRNA hybridization probes for quantitative molecular microbial ecology studies. Appl. Environ. Microbiol. 1996, 62, 4504–4513. [Google Scholar] [CrossRef]
- Moyer, C.L.; Dobbs, F.C.; Karl, D.M. Estimation of diversity and community structure through restriction fragment length polymorphism distribution analysis of bacterial 16S rRNA genes from a microbial mat at an active, hydrothermal vent system, Loihi Seamount, Hawaii. Appl. Environ. Microbiol. 1994, 60, 871–879. [Google Scholar] [CrossRef]
- Laguerre, G.; Allard, M.R.; Revoy, F.; Amarger, N. Rapid identification of Rhizobia by restriction fragment length polymor-phism analysis of PCR-amplified 16S rRNA genes. Appl. Environ. Microbiol. 1994, 60, 56–63. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T.; Order, I. Campylobacterales ord. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2005; Volume 2 (The Proteobacteria), part C (The Alpha-, Beta-, Delta-, and Epsilonproteobacteria); p. 1145. [Google Scholar]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. System. Evol. Microbiol. 2021, 71, 5056. [Google Scholar] [CrossRef]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar, A.; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; et al. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Westram, R.; Bader, K.; Prüße, E.; Kumar, Y.; Meier, H.; Glöckner, F.O.; Ludwig, W. ARB: A software environment for sequence data. In Handbook of Molecular Microbial Ecology I: Metagenomics and Complementary Approaches; de Bruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Cohan, F.M.; Perry, E.B. A systematics for identifying the fundamental units of bacterial diversity. Curr. Biol. 2008, 18, R1024–R1034. [Google Scholar] [CrossRef] [PubMed]
- Nelder, J.A.; Mead, R. A Simplex Method for Function Minimization. Comput. J. 1965, 7, 308–313. [Google Scholar] [CrossRef]
- Ward, D.M.; Bateson, M.M.; Ferris, M.J.; Kühl, M.; Wieland, A.; Koeppel, A.; Cohan, F.M. Cyanobacterial ecotypes in the microbial mat community of Mushroom Spring (Yellowstone National Park, Wyoming) as species-like units linking microbial community composition, structure and function. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- Becraft, E.D.; Wood, J.M.; Rusch, D.B.; Kühl, M.; Jensen, S.I.; Bryant, D.A.; Roberts, D.W.; Cohan, F.M.; Ward, D.M. The molecular dimension of microbial species: 1. Ecological distinctions among, and homogeneity within, putative ecotypes of Synechococcus inhabiting the cyanobacterial mat of Mushroom Spring, Yellowstone National Park. Front. Microbiol. 2015, 6, 590. [Google Scholar] [CrossRef]
- Becraft, E.D.; Wood, J.M.; Cohan, F.M.; Ward, D.M. Biogeography of American Northwest hot spring A/B’-lineage Synechococcus populations. Front. Microbiol. 2020, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Rodriquez-R, L.M.; Konstantinidis, K.T. Bypassing cultivation to identify bacterial species. Microbe 2014, 9, 111–118. [Google Scholar] [CrossRef]
- Shock, E.L.; Holland, M.; Meyer-Dombard, D.R.; Amend, J.P. Geochemical sources of energy for microbial metabolism in hydro-thermal ecosystems: Obsidian Pool, Yellowstone National Park. In Geothermal Biology and Geochemistry in Yellowstone National Park; Inskeep, W.P., McDermott, T.R., Eds.; Thermal Biology Institute, Montana State University: Bozeman, MT, USA, 2005; pp. 95–109. [Google Scholar]
- Pierson, B.K.; Castenholz, R.W. Bacteriochlorophylls in gliding filamentous prokaryotes from hot springs. Nat. New Biol. 1971, 233, 25–27. [Google Scholar] [CrossRef]
- Pierson, B.K.; Castenholz, R.W. A phototrophic gliding filamentous bacterium of hot springs, Chloroflexus aurantiacus, gen. and sp. nov. Arch. Microbiol. 1974, 100, 5–24. [Google Scholar] [CrossRef]
- Frigaard, N.-U.; Bryant, D.A. Chlorosomes: Antenna organelles in green photosynthetic bacteria. In Complex Intracellular Structures in Prokaryotes, Microbiology Monographs; Shively, J., Ed.; Springer: Berlin, Germany, 2006; Volume 2, pp. 79–114. [Google Scholar]
- Hurse, T.J.; Kappler, U.; Keller, J. Using anoxygenic photosynthetic bacteria for the removal of sulfide from wastewater. In Sulfur Metabolism in Phototrophic Organisms, Advances in Photosynthesis and Respiration; Hell, R., Dahl, C., Knaff, D., Leustel, T., Eds.; Springer: Dordrecht, Germany, 2008; Volume 27, pp. 8–22. [Google Scholar]
- Imhoff, J.F.; Thiel, V. Phylogeny and taxonomy of Chlorobiaceae. Photosynth. Res. 2010, 104, 123–136. [Google Scholar] [CrossRef]
- Manske, A.K. Ecological Diversity and Low Light Adaptation of Green Sulfur Bacteria. Ph.D. Dissertation, Ludwig -Maximilians-Universität München, München, Germany, 2007. [Google Scholar]
- Freed, S.; Robertson, S.; Meyer, T.; Kyndt, J. Draft whole-genome sequence of the green sulfur photosynthetic bacterium Chlorobaculum sp. strain 24CR, isolated from the Carmel River. Microbiol. Resour. Announc. 2019, 8, e00116-19. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, M.; Kakinuma, K.; Kawaguchi, R. Phylogenetic analysis of Salmonella, Shigella, and Escherichia coli strains on the basis of the gyrB gene sequence. J. Clin. Microbiol. 2002, 40, 2779–2785. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pei, T.; Yi, S.; Du, J.; Zhang, X.; Deng, X.; Yao, Q.; Deng, M.-R.; Zhu, H. Phylogenomic analysis substantiates the gyrB gene as a powerful molecular marker to efficiently differentiate the most closely related genera Myxococcus, Corallococcus, and Pyxidicoccus. Front. Microbiol. 2021, 12, 763359. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, E.; Overmann, J. Ecological significance of microdiversity: Identical 16S rRNA gene sequences can be found in bacteria with highly divergent genomes and ecophysiologies. Appl. Environ. Microbiol. 2004, 70, 4831–4839. [Google Scholar] [CrossRef] [PubMed]
- Nowack, S.; Olsen, M.T.; Schaible, G.; Becraft, E.D.; Shen, G.; Klapper, I.; Bryant, D.A.; Ward, D.M. The molecular dimension of microbial species: 2. Synechococcus isolates representative of putative ecotypes inhabiting different depths in the Mushroom Spring microbial mat exhibit different adaptive and acclimative responses to light. Front. Microbiol. 2015, 6, 626. [Google Scholar] [CrossRef] [PubMed]
- Allewalt, J.P.; Bateson, M.M.; Revsbech, N.P.; Slack, K.; Ward, D.M. Effect of temperature and light on growth and photosynthesis of Synechococcus isolates typical of those predominating in the Octopus Spring microbial mat community. Appl. Environ. Microbiol. 2006, 72, 544–550. [Google Scholar] [CrossRef]
- Olsen, M.T.; Nowack, S.; Wood, J.M.; Becraft, E.D.; LaButti, K.; Lipzen, A.; Martin, J.; Schackwitz, W.S.; Rusch, D.B.; Cohan, F.M.; et al. The molecular dimension of microbial species: 3. Comparative genomics of Synechoccocus isolates with different light responses and in situ diel transcription patterns of associated putative ecotypes in the Mushroom Spring microbial mat. Front. Microbiol. 2015, 6, 604. [Google Scholar] [CrossRef]
- Madigan, M.T.; Absher, J.N.; Mayers, J.E.; Asao, M.; Jung, D.O.; Bender, K.S.; Kempher, M.L.; Hayward, M.K.; Sanguedolce, S.A.; Brown, A.C.; et al. Allochromatium tepidum, sp. nov., a hot spring species of purple sulfur bacteria. Arch. Microbiol. 2022, 204, 115. [Google Scholar] [CrossRef]
- Ferris, M.J.; Ruff-Roberts, A.L.; Kopczynski, E.D.; Bateson, M.M.; Ward, D.M. Enrichment culture and microscopy conceal diverse thermophilic Synechococcus populations in a single hot spring microbial mat habitat. Appl. Environ. Microbiol. 1996, 62, 1045–1050. [Google Scholar] [CrossRef]
- Becraft, E.D.; Jackson, B.D.; Nowack, S.; Klapper, I.; Ward, D.M. Relationship between microorganisms inhabiting alkaline siliceous hot spring mat communities and overflowing water. Appl. Environ. Microbiol. 2020, 86, e00194-20. [Google Scholar] [CrossRef] [PubMed]
- Ruff-Roberts, A.L.; Kuenen, J.G.; Ward, D.M. Distribution of cultivated and uncultivated cyanobacteria and Chloroflexus-like bacteria in hot spring microbial mats. Appl. Environ. Microbiol. 1994, 60, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Jørgensen, B.B.; Paden, E.; Shilo, M. Sulfide dependent anoxygenic photosynthesis in the cyanobacterium Oscillatoria limnetica. Nature 1975, 257, 489–492. [Google Scholar] [CrossRef]
- Papke, R.T.; Ramsing, N.B.; Bateson, M.M.; Ward, D.M. Geographical isolation in hot spring cyanobacteria. Environ. Microbiol. 2003, 5, 650–659. [Google Scholar] [CrossRef]
- Gaisin, V.A.; Grouzdev, D.S.; Namsaraev, Z.B.; Sukhacheva, M.V.; Gorlenko, V.M.; Kuznetsov, B.B. Biogeography of thermophilic phototrophic bacteria belonging to Roseiflexus genus. FEMS Microbiol. Ecol. 2016, 92, fiw012. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Sample Date | Site Description | Temp °C | pH | Sulfide µM | Mat DNA GSB PCR product | GSB Enrichment | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 600F | Nested 600F | 619F | 27F | Temp °C | Gr a | PCR Prod | ||||||
| NZ | ||||||||||||
| TLS | 9/9/1986 | Ropey green film atop mud | 44.5 | 5.8 | 280–1850 | + | NT | NT | NT | NT | NT | NT |
| PHL | ||||||||||||
| Lip5 | 22/1/2002 | Grass green growth on the surface at the edge of the stream. See Figure 2D | 36.1 | 5.1–6.0 | 40 | NT | NT | NT | + | 36.5 b | + | + |
| YNP | ||||||||||||
| BS1-00 c | 21/6/2000 | Bright green mat in a muddy area | 41.6 | 6.4 | NT | No | No | + | NT | 36.5 | + | + |
| BS1-01 c | 6/8/2001 | Bright green mat in muddy area | 51.6 | 6.8 | 1.8 | No | No | + | NT | 44 | + | + |
| BS2 | 21/6/2000 | Yellowish-green mat on mud with thin white film (sulfur?) on top | 51.7 d–46 | 4.6 d–6.4 | NT | No | No | + | NT | 50 | No | NT |
| BS3.1 | 21/6/2000 | Pinkish-purple/yellowish green mat, floating on surface | 35.6 | 6.4 | NT | + | NT | NT | NT | 36.5 | + | + |
| BS3.2T e | 29/6/2000 | As BS3.1, but larger | 44.4 | 6.7 | NT | No | + | NT | NT | 44 | + | + |
| BS3.2M e | 29/6/2000 | Deep green layer, and below that, black mud and white filaments | 44.4 | 6.7 | NT | No | + | NT | NT | 44 | + | + |
| MV1 | 29/6/2000 | Bright green mat circling a small bubbling spring in the mud | 49–54 | 5.9 | NT | No | No | + | NT | 44 | + | + |
| MV2 | 29/6/2000 | Bright green mat circling small bubbling spring on mud | 36–44 | 5.7 | NT | No | + | + | NT | 36.5 | + | + |
| MV3 | 7/8/2001 | Yellowish-green mat on mud around bubbling spring. See Figure 3D | 34.3 | 5.8 | 2.7 | No | + | NT | NT | 34 | + | + |
| MV4 | 7/8/2001 | Yellowish-green mat on mud | 50.1 | 5.7 | 5.3 | + | + | NT | NT | 44 | + | + |
| MV5 | 7/8/2001 | Yellowish-green mat on mud | 39–40 | 5.7–5.9 | 4.7 | + | + | + | NT | 38 | + | + |
| MV6 | 31/7/2002 | Yellowish-green mat on mud circling a bubbling spring. See Figure 3E | 37–40 | 5.4–5.7 | 1.3 | + | + | NT | NT | 44 f | + | + |
| AT | 11/8/2001 | Olive green to orange mat sample in a shallow travertine pool near its top | 67.4 | 6.9 | 27.7 | + | NT | + | NT | 55 g | No | No |
| NG2 | 14/6/2000 | Fibrous dark olive green and orange mat in a tiny spring near the midpoint of Narrow Gauge | 52.5 | 6.2 | NT | No | NT | + | NT | 52 | No | No |
| Primer | Specificity | Sequence 5′ to 3′ | Reference |
|---|---|---|---|
| 27F | EUB | AGA GTT TGA TCC TGG CTC AG | [36,37] |
| 1492R | UNIV | GGT TAC CTT GTT ACG ACT T | [36,37] |
| GS.619F | GSB | GGG GTT AAA TCC ATG TGT GCT | [35] |
| GS.1144R | GSB | CAG TTC ART TAG AGT CC | [35] |
| GSB600F | GSB | GGG GGT TAA ATC CAT GTG | This study |
| 1392R | UNIV | ACG GGC GGT GTG TAC | [38] |
| Species/Clade | Cba. parvum | Cba. thiosulfatiphilum | Cba. tepidum | Cba. limnaeum | LIP-MAT | YNP-1-BS3.1/3.2-MAT | TLS | LIP-ENR | YNP-2 | YNP-2-MV4 | YNP-2-MV3/4/5 | YNP-2-BS1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cba. parvum (7) | 0–1.7 | |||||||||||
| Cba. thiosulfatiphilum (2) | 3.4–4.9 | 0.4 | ||||||||||
| Cba. tepidum (2) | 2.8–4.4 | 3.5–3.9 | 0 | |||||||||
| Cba. limnaeum (4) | 3.2–4,9 | 3.3–3.9 | 1.9–2.5 | 0–0.6 | ||||||||
| LIP-MAT | 3.1–4.0 | 4.0–4.4 | 3.6–3.8 | 1.2–2.2 | 0.0–0.2 | |||||||
| YNP-1-BS3.1/3.2-MAT | 2.6–4.7 | 3.5–4.1 | 2.9–3.5 | 1.7–2.6 | 0.3–0.8 | 0.2 | ||||||
| TLS | 2.2–4.1 | 3.5–4.1 | 0.0–0.4 | 1.5–2.5 | 2.7–3.1 | 2.1–3.1 | 0.1–0.6 | |||||
| LIP-ENR | 1.7–4.1 | 3.3–4.1 | 0.8–1.3 | 1.2–2.2 | 1.7––3.6 | 1.9–3.4 | 0.7–1.2 | 0.0–0.4 | ||||
| YNP-2 | 1.5–5.5 | 2.0–5.3 | 1.4–2.8 | 1.2–3.0 | 2.2–4.1 | 2.7–4.5 | 1.2–2.5 | 0.8–2.1 | 0–2.0 | |||
| YNP-2 MV4 | 2.9–4.9 | 2.9–5.0 | 1.5–2.0 | 1.3–2.2 | 2.9–3.6 | 2.8–4.0 | 1.6–2.2 | 0.9–1.8 | 0.0–1.0 | |||
| YNP-2-MV3/4/5 | 1.9–5.0 | 2.3–5.0 | 1.6–2.1 | 1.3–2.2 | 2.5–3.7 | 2.7–4.5 | 1.4–2.5 | 1.0–2.0 | 0.2–1.7 | 0.0–1.2 | ||
| YNP-2-BS1 | 1.6–5.3 | 2.6–3.6 | 1.4–2.0 | 1.7–2.6 | 2.3–3.4 | 2.7–4.2 | 2.3–3.4 | 0.8–1.9 | 0.2–1.2 | 0.4–1.8 | 0.0–1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedard, D.L.; Van Slyke, G.; Nübel, U.; Bateson, M.M.; Brumfield, S.; An, Y.J.; Becraft, E.D.; Wood, J.M.; Thiel, V.; Ward, D.M. Geographic and Ecological Diversity of Green Sulfur Bacteria in Hot Spring Mat Communities. Microorganisms 2023, 11, 2921. https://doi.org/10.3390/microorganisms11122921
Bedard DL, Van Slyke G, Nübel U, Bateson MM, Brumfield S, An YJ, Becraft ED, Wood JM, Thiel V, Ward DM. Geographic and Ecological Diversity of Green Sulfur Bacteria in Hot Spring Mat Communities. Microorganisms. 2023; 11(12):2921. https://doi.org/10.3390/microorganisms11122921
Chicago/Turabian StyleBedard, Donna L., Greta Van Slyke, Ulrich Nübel, Mary M. Bateson, Sue Brumfield, Yong Jun An, Eric D. Becraft, Jason M. Wood, Vera Thiel, and David M. Ward. 2023. "Geographic and Ecological Diversity of Green Sulfur Bacteria in Hot Spring Mat Communities" Microorganisms 11, no. 12: 2921. https://doi.org/10.3390/microorganisms11122921