
Evaluation of Holmium(III), Erbium(III), and Gadolinium(III) Accumulation by Cyanobacteria Arthrospira platensis Using Neutron Activation Analysis and Elements’ Effects on Biomass Quantity and Biochemical Composition
 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Object of Study
2.2. Experimental Design
2.3. Neutron Activation Analysis (NAA)
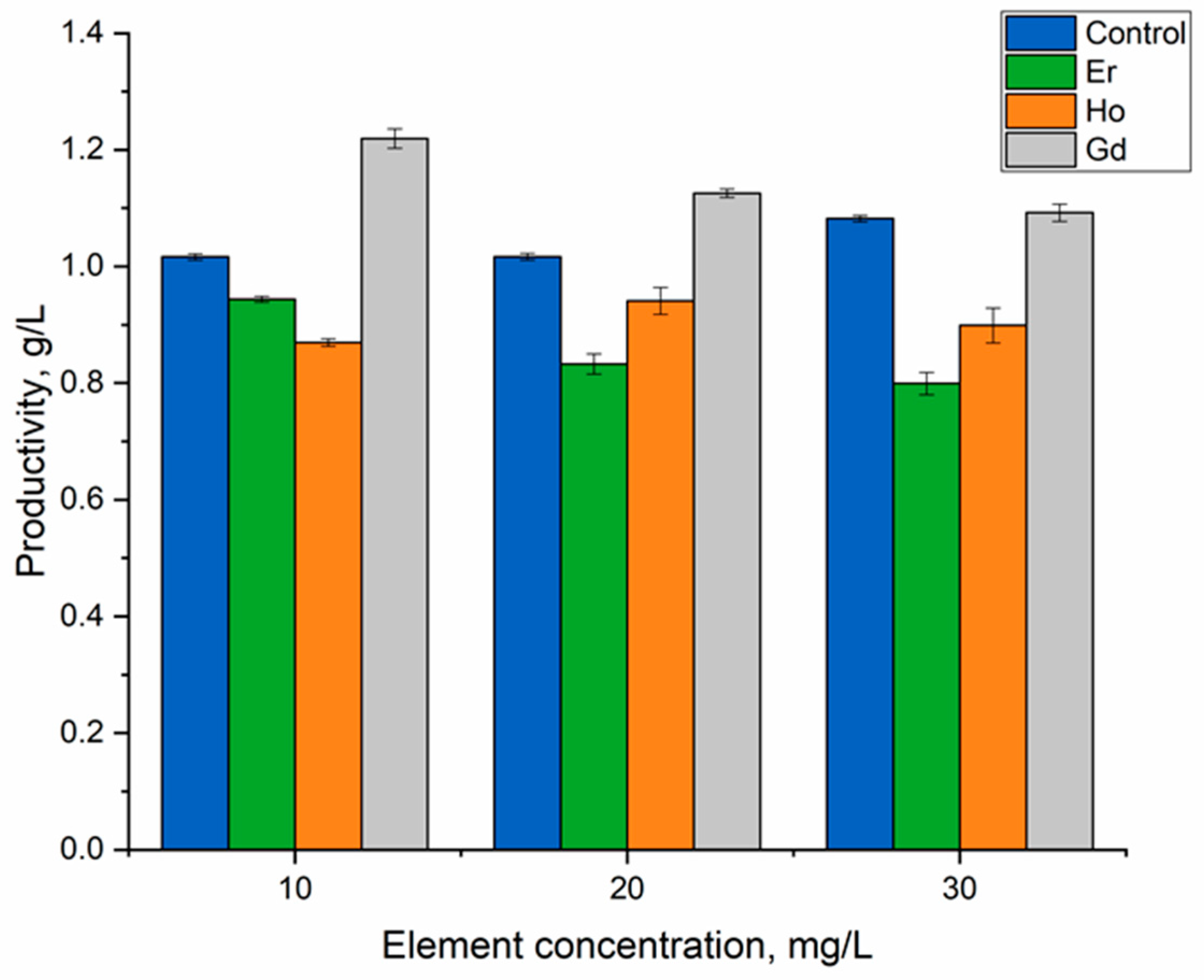
2.4. Biomass Quantity
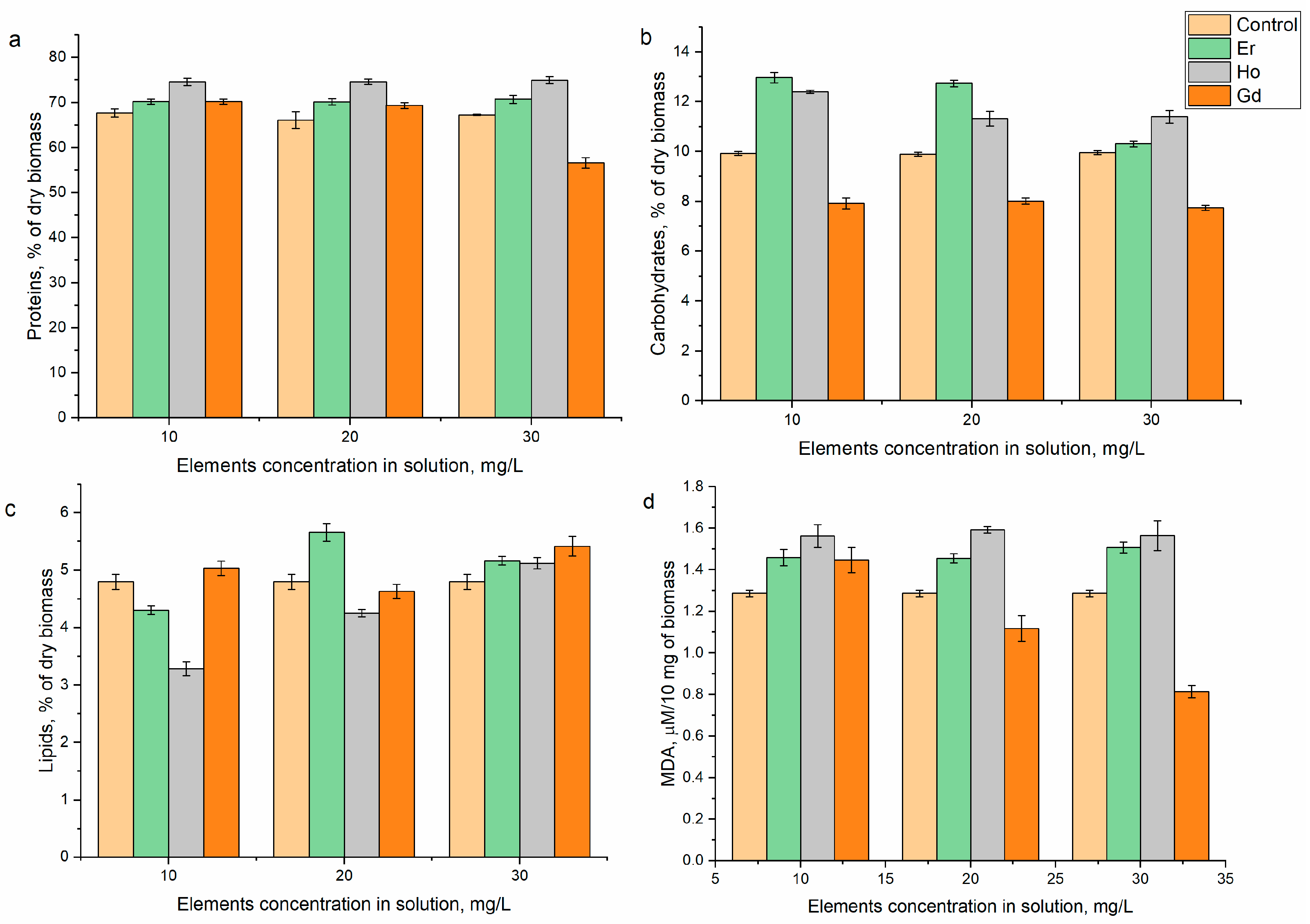
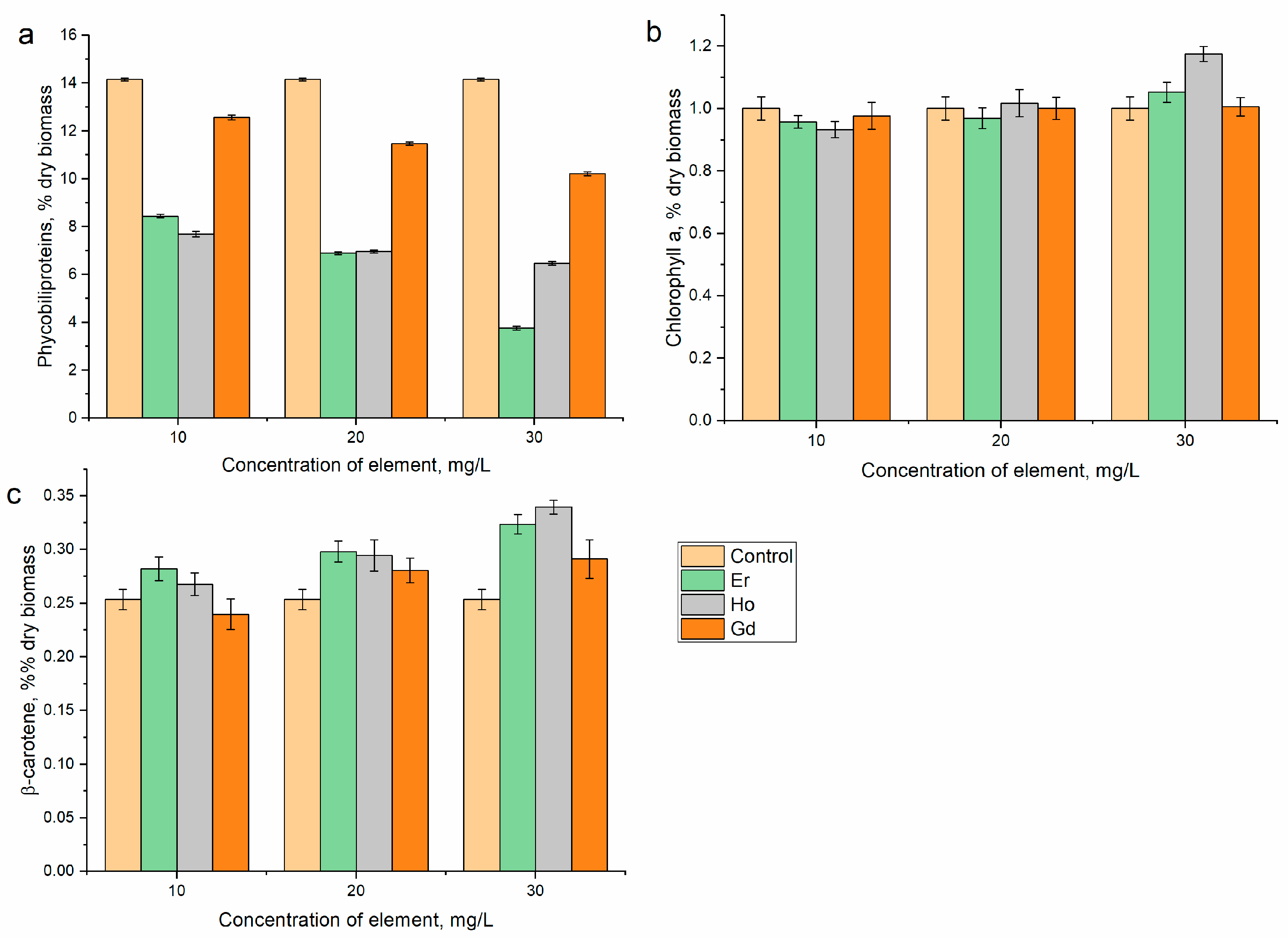
2.5. Biochemical Analysis and Determination of the Antioxidant Activity
2.6. Antioxidant Activity According to the ABTS+ Radical Cation Assay
2.7. Statistical Analysis
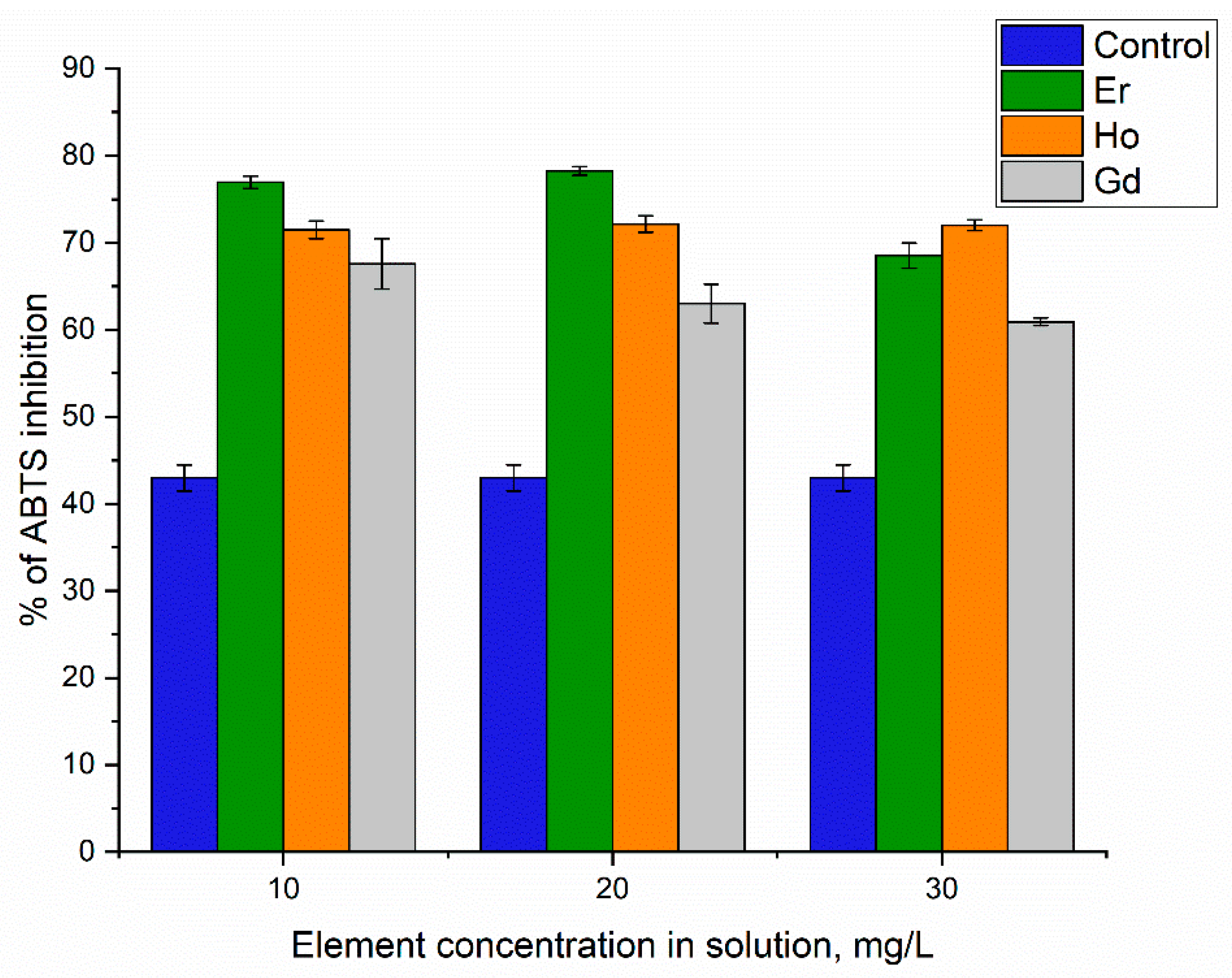
3. Results and Discussion
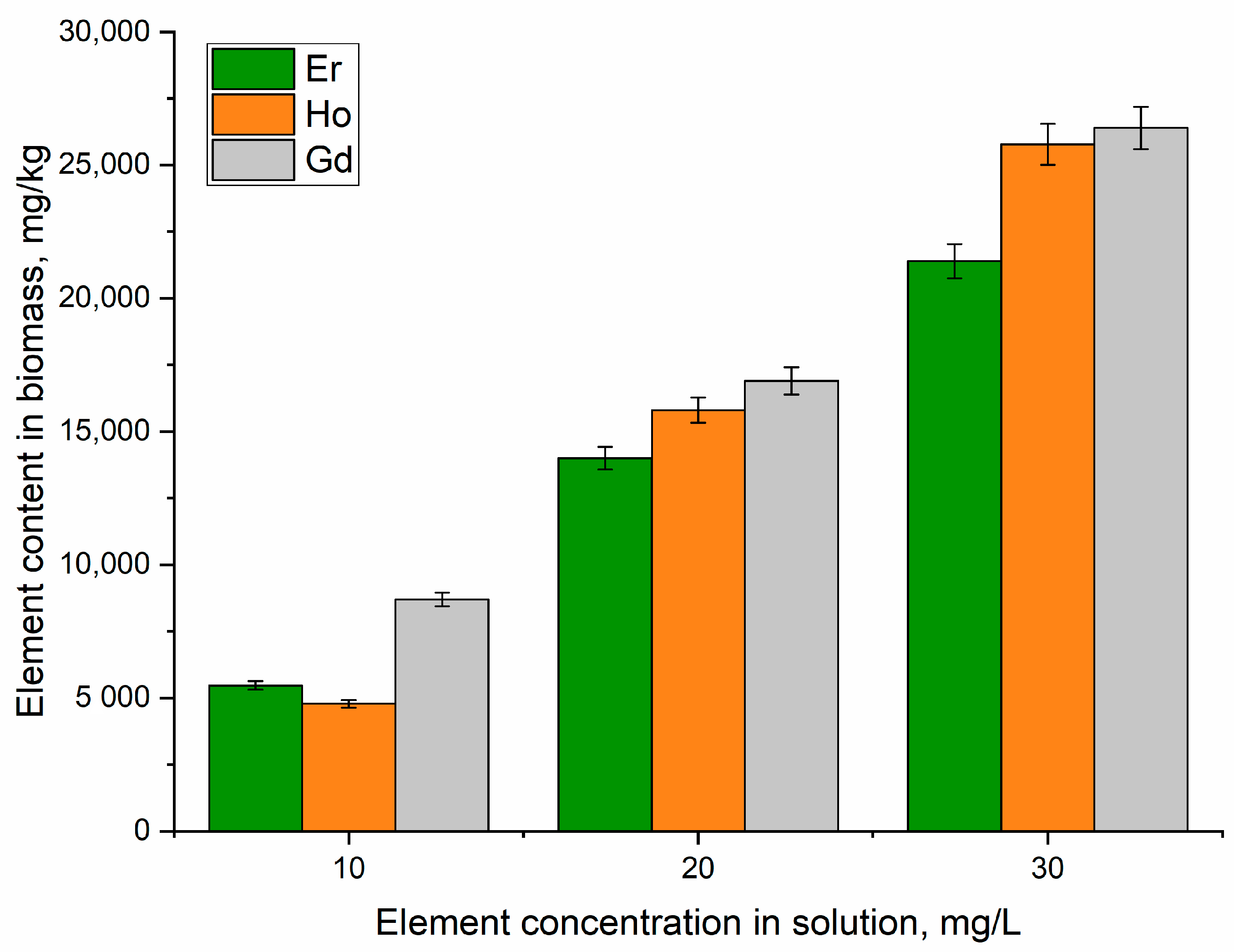
Element Uptake by A. platensis Biomass
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Goecke, F.; Zachleder, V.; Vítová, M. Rare Earth Elements and Algae: Physiological Effects, Biorefi Nery and Recycling. In Algal Biorefineries: Volume 2: Products and Refinery Design; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 339–363. ISBN 9783319202006. [Google Scholar]
- Ramos, S.J.; Dinali, G.S.; Oliveira, C.; Martins, G.C.; Moreira, C.G.; Siqueira, J.O.; Guilherme, L.R.G. Rare Earth Elements in the Soil Environment. Curr. Pollut. Rep. 2016, 2, 28–50. [Google Scholar] [CrossRef]
- Das, N.; Das, D. Recovery of Rare Earth Metals through Biosorption: An Overview. J. Rare Earths 2013, 31, 933–943. [Google Scholar] [CrossRef]
- Gallardo, K.; Castillo, R.; Mancilla, N.; Remonsellez, F. Biosorption of Rare-Earth Elements From Aqueous Solutions Using Walnut Shell. Front. Chem. Eng. 2020, 2, 4. [Google Scholar] [CrossRef]
- Heilmann, M.; Breiter, R.; Becker, A.M. Towards Rare Earth Element Recovery from Wastewaters: Biosorption Using Phototrophic Organisms. Appl. Microbiol. Biotechnol. 2021, 105, 5229–5239. [Google Scholar] [CrossRef]
- Malhotra, N.; Hsu, H.S.; Liang, S.T.; Roldan, M.J.M.; Lee, J.S.; Ger, T.R.; Hsiao, C.D. An Updated Review of Toxicity Effect of the Rare Earth Elements (REEs) on Aquatic Organisms. Animals 2020, 10, 1663. [Google Scholar] [CrossRef] [PubMed]
- Piarulli, S.; Hansen, B.H.; Ciesielski, T.; Zocher, A.L.; Malzahn, A.; Olsvik, P.A.; Sonne, C.; Nordtug, T.; Jenssen, B.M.; Booth, A.M.; et al. Sources, Distribution and Effects of Rare Earth Elements in the Marine Environment: Current Knowledge and Research Gaps. Environ. Pollut. 2021, 291, 118230. [Google Scholar] [CrossRef] [PubMed]
- Rim, K.T.; Koo, K.H.; Park, J.S. Toxicological Evaluations of Rare Earths and Their Health Impacts to Workers: A Literature Review. Saf. Health Work 2013, 4, 12–26. [Google Scholar] [CrossRef]
- Brouziotis, A.A.; Giarra, A.; Libralato, G.; Pagano, G.; Guida, M.; Trifuoggi, M. Toxicity of Rare Earth Elements: An Overview on Human Health Impact. Front. Environ. Sci. 2022, 10, 1617. [Google Scholar] [CrossRef]
- Blinova, I.; Vija, H.; Lukjanova, A.; Muna, M.; Syvertsen-Wiig, G.; Kahru, A. Assessment of the Hazard of Nine (Doped) Lanthanides-Based Ceramic Oxides to Four Aquatic Species. Sci. Total Environ. 2018, 612, 1171–1176. [Google Scholar] [CrossRef]
- Bergsten-Torralba, L.R.; Magalhães, D.P.; Giese, E.C.; Nascimento, C.R.S.; Pinho, J.V.A.; Buss, D.F. Toxicity of Three Rare Earth Elements, and Their Combinations to Algae, Microcrustaceans, and Fungi. Ecotoxicol. Environ. Saf. 2020, 201, 110795. [Google Scholar] [CrossRef]
- Xiong, J.Q.; Ru, S.; Zhang, Q.; Jang, M.; Kurade, M.B.; Kim, S.H.; Jeon, B.H. Insights into the Effect of Cerium Oxide Nanoparticle on Microalgal Degradation of Sulfonamides. Bioresour. Technol. 2020, 309, 123452. [Google Scholar] [CrossRef] [PubMed]
- Mahaye, N.; Musee, N. Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella Subcapitata to Cerium Oxide Nanoparticles. Toxics 2023, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Cardon, P.Y.; Triffault-Bouchet, G.; Caron, A.; Rosabal, M.; Fortin, C.; Amyot, M. Toxicity and Subcellular Fractionation of Yttrium in Three Freshwater Organisms: Daphnia Magna, Chironomus Riparius, and Oncorhynchus Mykiss. ACS Omega 2019, 4, 13747–13755. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Walton, M.; Das, G.; Dohnalkova, A.; Vanzin, G.; Anderko, A. Impacts of Anthropogenic Gadolinium on the Activity of the Ammonia Oxidizing Bacterium Nitrosomonas Europaea. Chemosphere 2020, 257, 127250. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, H.; Nolde, J.; Berger, S.; Heise, S. Aquatic Ecotoxicity of Lanthanum—A Review and an Attempt to Derive Water and Sediment Quality Criteria. Ecotoxicol. Environ. Saf. 2016, 124, 213–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, Y.; Zhang, R.; Guo, G.; Zhang, K.; Fan, Y.; Feng, C.; Li, H. Water Quality Criteria for Lanthanum for Freshwater Aquatic Organisms Derived via Species Sensitivity Distributions and Interspecies Correlation Estimation Models. Ecotoxicology 2022, 31, 897–908. [Google Scholar] [CrossRef]
- Al-Amin, A.; Parvin, F.; Chakraborty, J.; Kim, Y.I. Cyanobacteria Mediated Heavy Metal Removal: A Review on Mechanism, Biosynthesis, and Removal Capability. Environ. Technol. Rev. 2021, 10, 44–57. [Google Scholar] [CrossRef]
- Fischer, C.B.; Körsten, S.; Rösken, L.M.; Cappel, F.; Beresko, C.; Ankerhold, G.; Schönleber, A.; Geimer, S.; Ecker, D.; Wehner, S. Cyanobacterial Promoted Enrichment of Rare Earth Elements Europium, Samarium and Neodymium and Intracellular Europium Particle Formation. RSC Adv. 2019, 9, 32581–32593. [Google Scholar] [CrossRef]
- Rakic, I.; Kevresan, Z.; Kovac, R.; Kravic, S.; Svircev, Z.; Djurovic, A.; Stojanovic, Z. Bioaccumulation and Biosorption Study of Heavy Metals Removal by Cyanobacteria Nostoc sp. Chem. Ind. Chem. Eng. Q 2023, 29, 291–298. [Google Scholar] [CrossRef]
- Lima, É.C.; Pinto, D.; Schadeck Netto, M.; Dos Reis, G.S.; Silva, L.F.O.; Dotto, G.L. Biosorption of Neodymium (Nd) from Aqueous Solutions Using Spirulina Platensis Sp. Strains. Polymers 2022, 14, 4585. [Google Scholar] [CrossRef]
- Wenqiang, Z.; Qing, S.; Baoquan, X.; Wenqiang, Z.; Qing, S.; Baoquan, X. Study on Biosorption Characteristics of Spirulina to Rare Earth Ytterbium Ions in Simulated Mine Wastewater. Nonferrous Met. Sci. Eng. 2019, 10, 103–112. [Google Scholar] [CrossRef]
- Shu, Q.; Liao, C.-F.; Zou, W.-Q.; Xu, B.-Q.; Tan, Y.-H. Recovery of Rare Earth Element Ytterbium(III) by Dried Powdered Biomass of Spirulina: Adsorption Isotherm, Kinetic and Thermodynamic Study. Trans. Nonferrous Met. Soc. China (Engl. Ed.) 2021, 31, 1127–1139. [Google Scholar] [CrossRef]
- Sadovsky, D.; Brenner, A.; Astrachan, B.; Asaf, B.; Gonen, R. Biosorption Potential of Cerium Ions Using Spirulina Biomass. J. Rare Earths 2016, 34, 644–652. [Google Scholar] [CrossRef]
- Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Biosorption and Bioaccumulation Capacity of Arthospira Platensis toward Europium Ions. Water 2022, 14, 2128. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Cepoi, L.; Rudi, L.; Chiriac, T.; Grozdov, D.; Pavlov, S.; Djur, S. Accumulation of Dysprosium, Samarium, Terbium, Lanthanum, Neodymium and Ytterbium by Arthrospira Platensis and Their Effects on Biomass Biochemical Composition. J. Rare Earths 2021, 39, 1133–1143. [Google Scholar] [CrossRef]
- Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Biosorption and Bioaccumulation Capacity of Arthrospira Platensis toward Yttrium Ions. Metals 2022, 12, 1465. [Google Scholar] [CrossRef]
- Sayqal, A.; Ahmed, O.B. Advances in Heavy Metal Bioremediation: An Overview. Appl. Bionics Biomech. 2021, 2021, 1609149. [Google Scholar] [CrossRef]
- Cepoi, L.; Zinicovscaia, I.; Rudi, L.; Chiriac, T.; Rotari, I.; Turchenko, V.; Djur, S. Effects of PEG-Coated Silver and Gold Nanoparticles on Spirulina Platensis Biomass during Its Growth in a Closed System. Coatings 2020, 10, 717. [Google Scholar] [CrossRef]
- Tsuruta, T. Selective Accumulation of Light or Heavy Rare Earth Elements Using Gram-Positive Bacteria. Colloids Surf. B Biointerfaces 2006, 52, 117–122. [Google Scholar] [CrossRef]
- Řezanka, T.; Kaineder, K.; Mezricky, D.; Řezanka, M.; Bišová, K.; Zachleder, V.; Vítová, M. The Effect of Lanthanides on Photosynthesis, Growth, and Chlorophyll Profile of the Green Alga Desmodesmus Quadricauda. Photosynth. Res. 2016, 130, 335–346. [Google Scholar] [CrossRef]
- Makhi, V.V.; Ahmad, A.; Chaugule, B.B. Effect of Bismuth, Gadolinium, and Cadmium Nanoparticles on Biomass, Carotenoid, and Lipid Content of Dunaliella salina (Dunal) Teodoresco. Curr. Microbiol. 2022, 79, 30. [Google Scholar] [CrossRef] [PubMed]
- Goecke, F.; Vítová, M.; Lukavský, J.; Nedbalová, L.; Řezanka, T.; Zachleder, V. Effects of Rare Earth Elements on Growth Rate, Lipids, Fatty Acids and Pigments in Microalgae. Phycol. Res. 2017, 65, 226–234. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zinicovscaia, I.; Cepoi, L.; Rudi, L.; Chiriac, T.; Grozdov, D. Evaluation of Holmium(III), Erbium(III), and Gadolinium(III) Accumulation by Cyanobacteria Arthrospira platensis Using Neutron Activation Analysis and Elements’ Effects on Biomass Quantity and Biochemical Composition. Microorganisms 2024, 12, 122. https://doi.org/10.3390/microorganisms12010122
Zinicovscaia I, Cepoi L, Rudi L, Chiriac T, Grozdov D. Evaluation of Holmium(III), Erbium(III), and Gadolinium(III) Accumulation by Cyanobacteria Arthrospira platensis Using Neutron Activation Analysis and Elements’ Effects on Biomass Quantity and Biochemical Composition. Microorganisms. 2024; 12(1):122. https://doi.org/10.3390/microorganisms12010122
Chicago/Turabian StyleZinicovscaia, Inga, Liliana Cepoi, Ludmila Rudi, Tatiana Chiriac, and Dmitrii Grozdov. 2024. "Evaluation of Holmium(III), Erbium(III), and Gadolinium(III) Accumulation by Cyanobacteria Arthrospira platensis Using Neutron Activation Analysis and Elements’ Effects on Biomass Quantity and Biochemical Composition" Microorganisms 12, no. 1: 122. https://doi.org/10.3390/microorganisms12010122