

Loss of Biliverdin Reductase Increases Oxidative Stress in the Cyanobacterium Synechococcus sp. PCC 7002
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
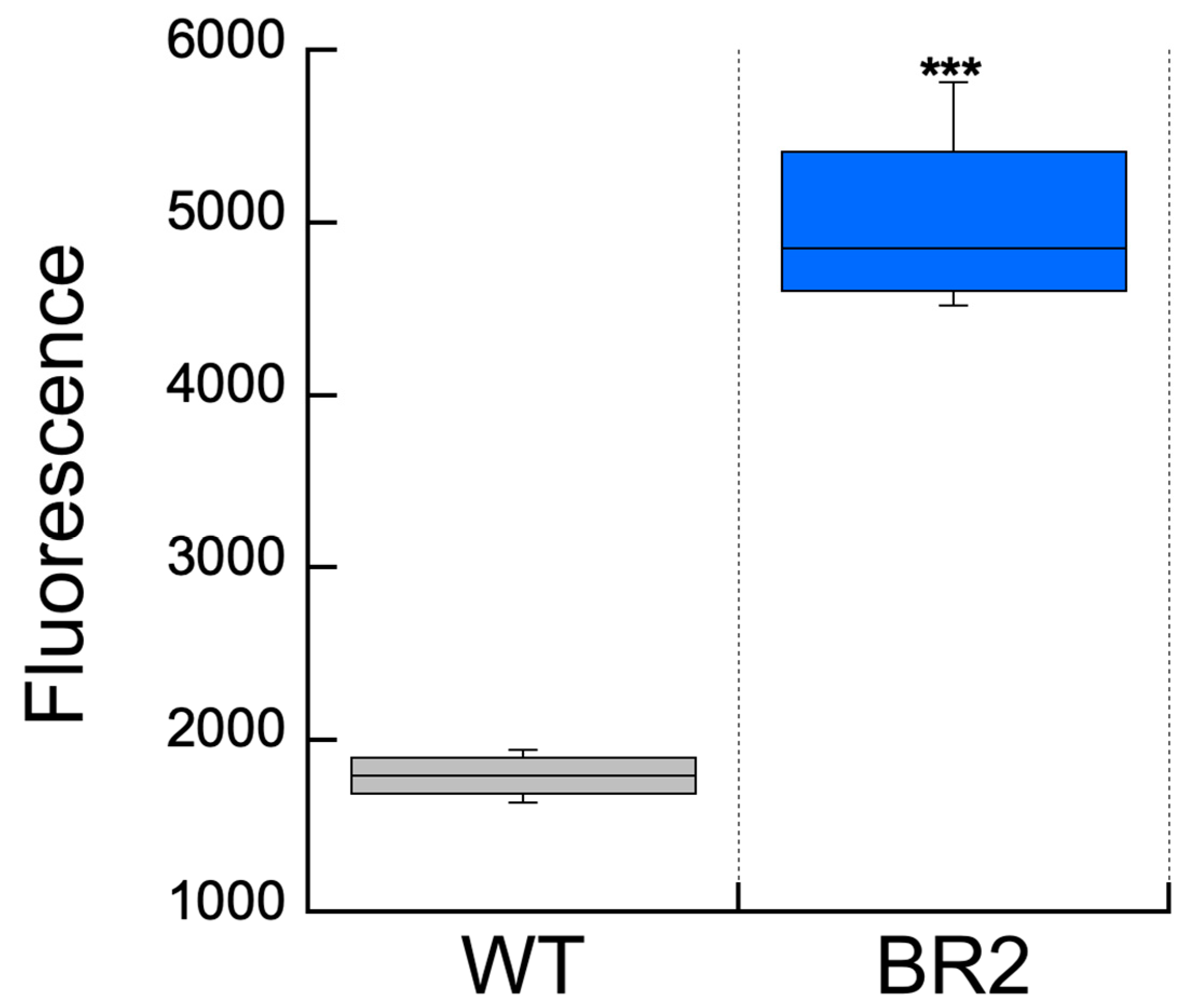
3. Results
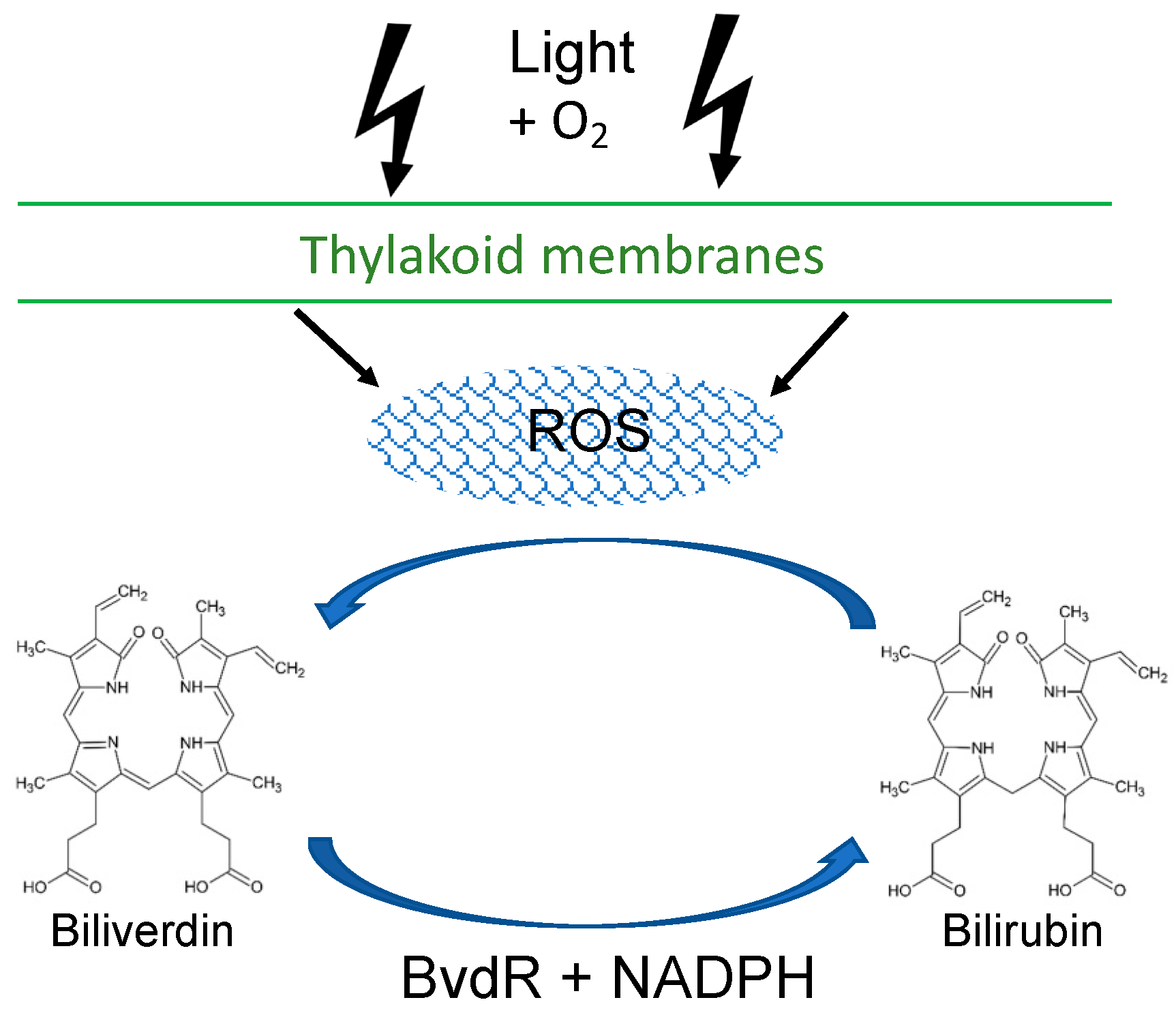
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Billing, B.H. Twenty-five years of progress in bilirubin metabolism (1952-77). Gut 1978, 19, 481–491. [Google Scholar] [CrossRef]
- McDonagh, A.F. Turning green to gold. Nat. Struct. Biol. 2001, 8, 198–200. [Google Scholar] [CrossRef]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.; Glazer, A.N.; Ames, B.N. Antioxidant activity of albumin-bound bilirubin. Proc. Natl. Acad. Sci. USA 1987, 84, 5918–5922. [Google Scholar] [CrossRef] [PubMed]
- Doré, S.; Takahashi, M.; Ferris, C.D.; Zakhary, R.; Hester, L.D.; Guastella, D.; Snyder, S.H. Bilirubin, formed by activation of heme oxygenase-2, protects neurons against oxidative stress injury. Proc. Natl. Acad. Sci. USA 1999, 96, 2445–2450. [Google Scholar] [CrossRef]
- Barañano, D.E.; Rao, M.; Ferris, C.D.; Snyder, S.H. Biliverdin reductase: A major physiologic cytoprotectant. Proc. Natl. Acad. Sci. USA 2002, 99, 16093–16098. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.W.; Saleh, M.; Higginson, D.S.; Paul, B.D.; Juluri, K.R.; Snyder, S.H. Bilirubin and glutathione have complementary antioxidant and cytoprotective roles. Proc. Natl. Acad. Sci. USA 2009, 106, 5171–5176. [Google Scholar] [CrossRef]
- Maghzal, G.J.; Leck, M.C.; Collinson, E.; Li, C.; Stocker, R. Limited role for the bilirubin-biliverdin redox amplification cycle in the cellular antioxidant protection by biliverdin reductase. J. Biol. Chem. 2009, 284, 29251–29259. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.W.; Snyder, S.H. Cycling the wagons for biliverdin reductase. J. Biol. Chem. 2009, 284, LE11. [Google Scholar] [CrossRef]
- McDonagh, A.F. The biliverdin-bilirubin antioxidant cycle of cellular protection: Missing a wheel? Free Radic. Biol. Med. 2010, 49, 814–820. [Google Scholar] [CrossRef]
- Mancuso, C.; Barone, E.; Guido, P.; Miceli, F.; Di Domenico, F.; Perluigi, M.; Santangelo, R.; Preziosi, P. Inhibition of lipid peroxidation and protein oxidation by endogenous and exogenous antioxidants in rat brain microsomes in vitro. Neurosci. Lett. 2012, 518, 101–105. [Google Scholar] [CrossRef]
- Jayanti, S.; Vitek, L.; Tiribelli, C.; Gazzin, S. The role of bilirubin and the other “yellow players” in neurodegenerative diseases. Antioxidants 2020, 9, 900. [Google Scholar] [CrossRef]
- Salim, M.; Brown-Kipphut, B.A.; Maines, M.D. Human biliverdin reductase is autophosphorylated, and phosphorylation is required for bilirubin formation. J. Biol. Chem. 2001, 276, 10929–10934. [Google Scholar] [CrossRef]
- Ahmad, Z.; Salim, M.; Maines, M.D. Human biliverdin reductase is a leucine zipper-like DNA-binding protein and functions in transcriptional activation of heme oxygenase-1 by oxidative stress. J. Biol. Chem. 2002, 277, 9226–9232. [Google Scholar] [CrossRef]
- Kravets, A.; Hu, Z.; Miralem, T.; Torno, M.D.; Maines, M.D. Biliverdin reductase, a novel regulator for induction of activating transcription factor-2 and heme oxygenase-1. J. Biol. Chem. 2004, 279, 19916–19923. [Google Scholar] [CrossRef]
- Maines, M.D. New insights into biliverdin reductase functions: Linking heme metabolism to cell signaling. Physiology 2005, 20, 382–389. [Google Scholar] [CrossRef]
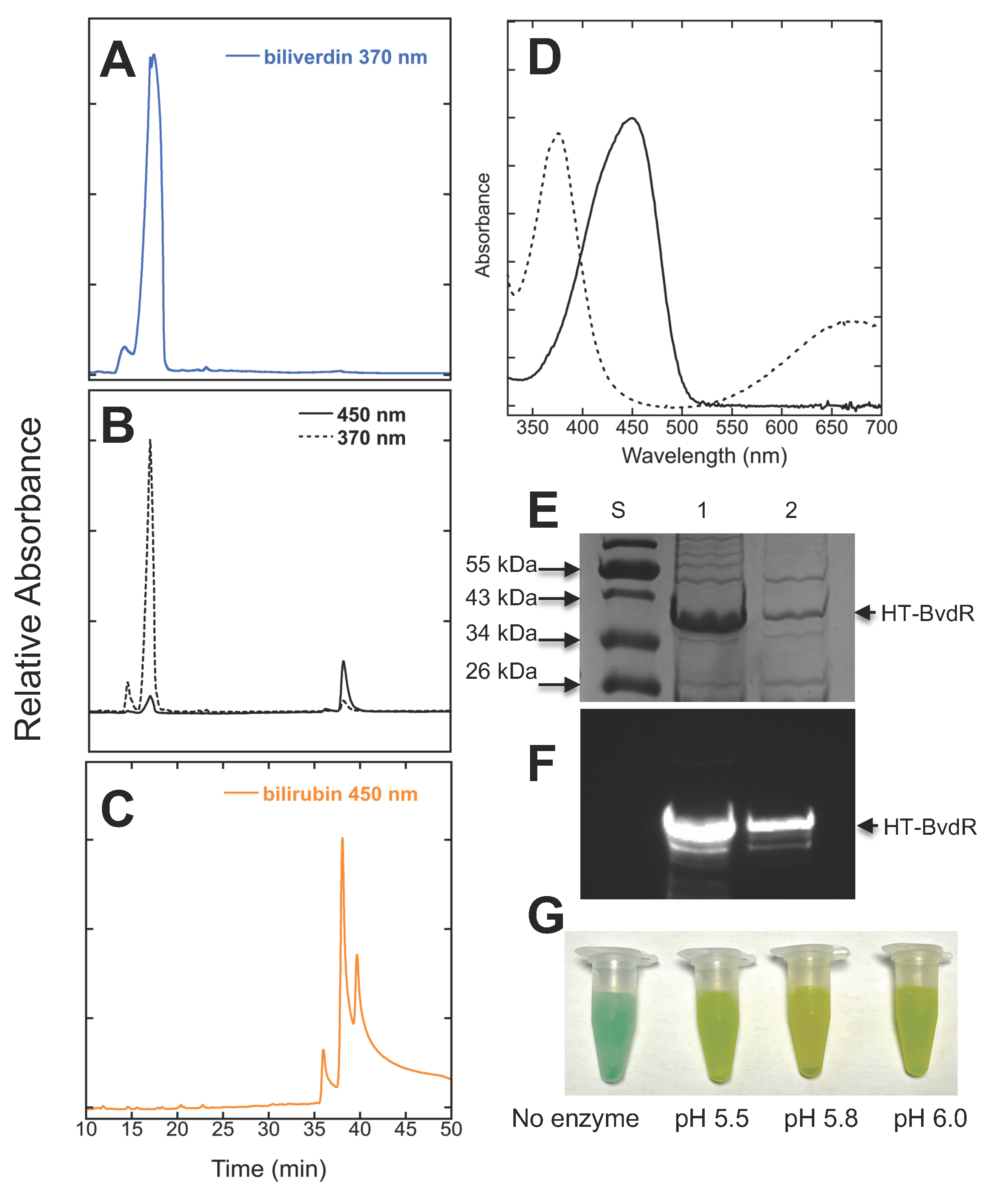
- Schluchter, W.M.; Glazer, A.N. Characterization of cyanobacterial biliverdin reductase: Conversion of biliverdin to bilirubin is important for normal phycobiliprotein biosynthesis. J. Biol. Chem. 1997, 272, 13562–13569. [Google Scholar] [CrossRef]
- Hayes, J.M.; Mantle, T.J. The effect of pH on the initial rate kinetics of the dimeric biliverdin-IX alpha reductase from the cyanobacterium Synechocystis PCC 6803. FEBS J. 2009, 276, 4414–4425. [Google Scholar] [CrossRef]
- Takao, H.; Hirabayashi, K.; Nishigaya, Y.; Kouriki, H.; Nakaniwa, T.; Hagiwara, Y.; Harada, J.; Sato, H.; Yamazaki, T.; Sakakibara, Y.; et al. A substrate-bound structure of cyanobacterial biliverdin reductase identifies stacked substrates as critical for activity. Nat. Commun. 2017, 8, 14397. [Google Scholar] [CrossRef]
- Ludwig, M.; Bryant, D. Transcription profiling of the model cyanobacterium Synechococcus sp. strain PCC 7002 by next-gen (SOLiD™) sequencing of cDNA. Front. Microbiol. 2011, 2, 41. [Google Scholar] [CrossRef]
- Garczarek, L.; Guyet, U.; Doré, H.; Farrant, G.K.; Hoebeke, M.; Brillet-Guéguen, L.; Bisch, A.; Ferrieux, M.; Siltanen, J.; Corre, E.; et al. Cyanorak v2.1: A scalable information system dedicated to the visualization and expert curation of marine and brackish picocyanobacteria genomes. Nucleic Acids Res. 2021, 49, D667–D676. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold protein structure database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2021, 50, D439–D444. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Shen, G.Z.; Bryant, D.A. Characterization of a Synechococcus sp. PCC 7002 mutant lacking Photosystem I. Protein assembly and energy distribution in the absence of the Photosystem I reaction center core complex. Photosynth. Res. 1995, 41, 41–53. [Google Scholar] [CrossRef]
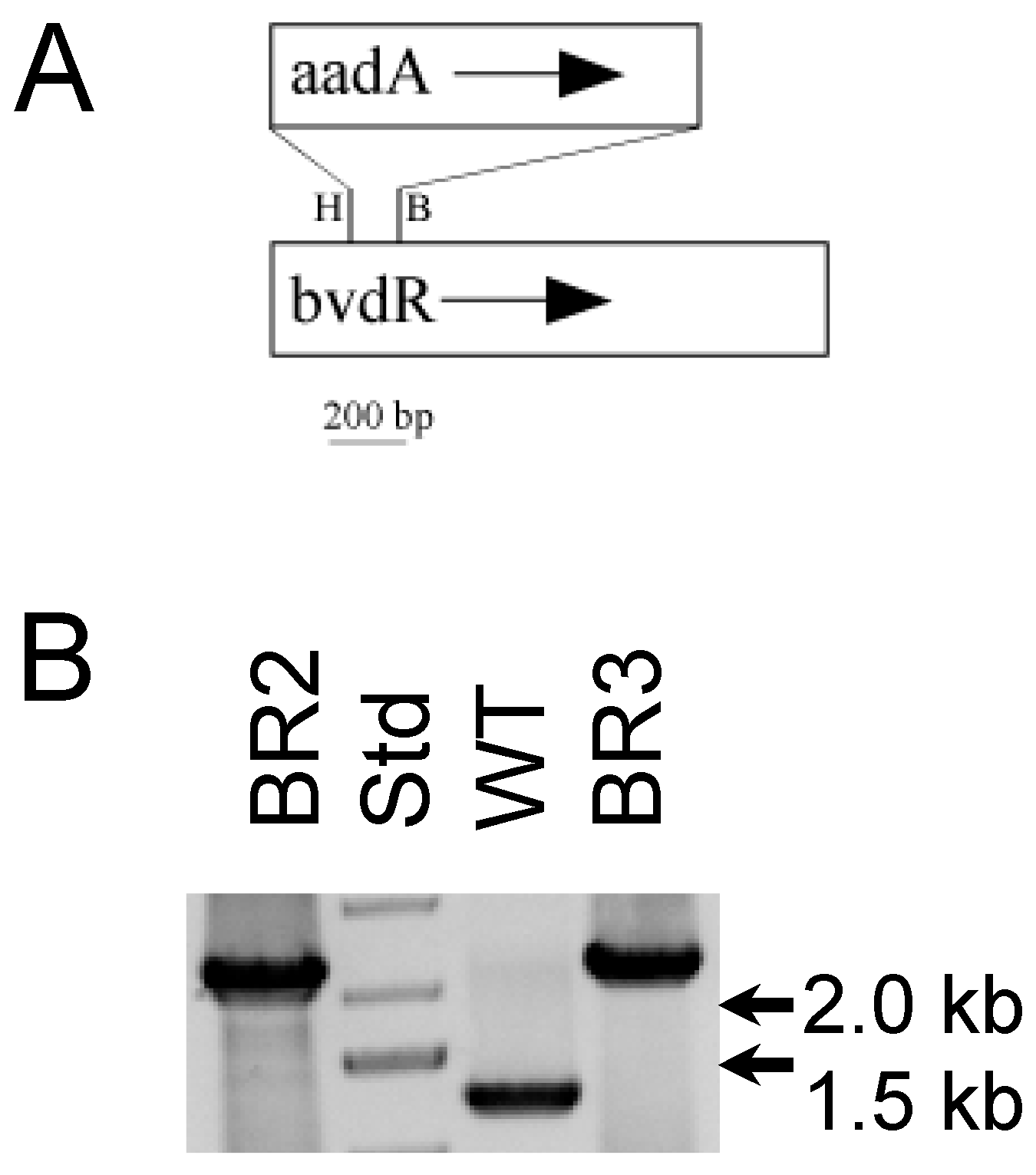
- Frigaard, N.U.; Sakuragi, Y.; Bryant, D.A. Gene inactivation in the cyanobacterium Synechococcus sp. PCC 7002 and the green sulfur bacterium Chlorobium tepidum using in vitro-made DNA constructs and natural transformation. Methods Mol. Biol. 2004, 274, 325–340. [Google Scholar] [CrossRef]
- Studier, F.W. Protein production by auto-induction in high-density shaking cultures. Prot. Exp. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef]
- Carrigee, L.A.; Frick, J.P.; Karty, J.A.; Garczarek, L.; Partensky, F.; Schluchter, W.M. MpeV is a lyase isomerase that ligates a doubly linked phycourobilin on the β-subunit of phycoerythrin I and II in marine Synechococcus. J. Biol. Chem. 2021, 296, 100031. [Google Scholar] [CrossRef]
- Biswas, A.; Vasquez, Y.M.; Dragomani, T.M.; Kronfel, M.L.; Williams, S.R.; Alvey, R.M.; Bryant, D.A.; Schluchter, W.M. Biosynthesis of cyanobacterial phycobiliproteins in Escherichia coli: Chromophorylation efficiency and specificity of all bilin lyases from Synechococcus sp strain PCC 7002. Appl. Environ. Microbiol. 2010, 76, 2729–2739. [Google Scholar] [CrossRef]
- Miller, C.A.; Leonard, H.S.; Pinsky, I.G.; Turner, B.M.; Williams, S.R.; Harrison, L., Jr.; Fletcher, A.F.; Shen, G.; Bryant, D.A.; Schluchter, W.M. Biogenesis of phycobiliproteins.III. CpcM is the asparagine methyltransferase for phycobiliprotein β-subunits in cyanobacteria. J. Biol. Chem. 2008, 283, 19293–19300. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of Photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- MacKinney, G. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Shen, G.; Schluchter, W.M.; Bryant, D.A. Biogenesis of phycobiliproteins. I. cpcS-I and cpcU mutants of the cyanobacterium Synechococcus sp. PCC 7002 define a heterodimeric phycocaynobilin lyase specific for beta -phycocyanin and allophycocyanin subunits. J. Biol. Chem. 2008, 283, 7503–7512. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Brand, J.J. Sequential events in the photoinhibition of Synechocystis under sodium stress. Plant Physiol. 1989, 91, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Inoue-Sakamoto, K.; Bryant, D.A. A novel nitrate/nitrite permease in the marine cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacteriol. 1999, 181, 7363–7372. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Bryant, D.A. The biosynthetic pathway for synechoxanthin, an aromatic carotenoid synthesized by the euryhaline, unicellular cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacteriol. 2008, 190, 7966–7974. [Google Scholar] [CrossRef]
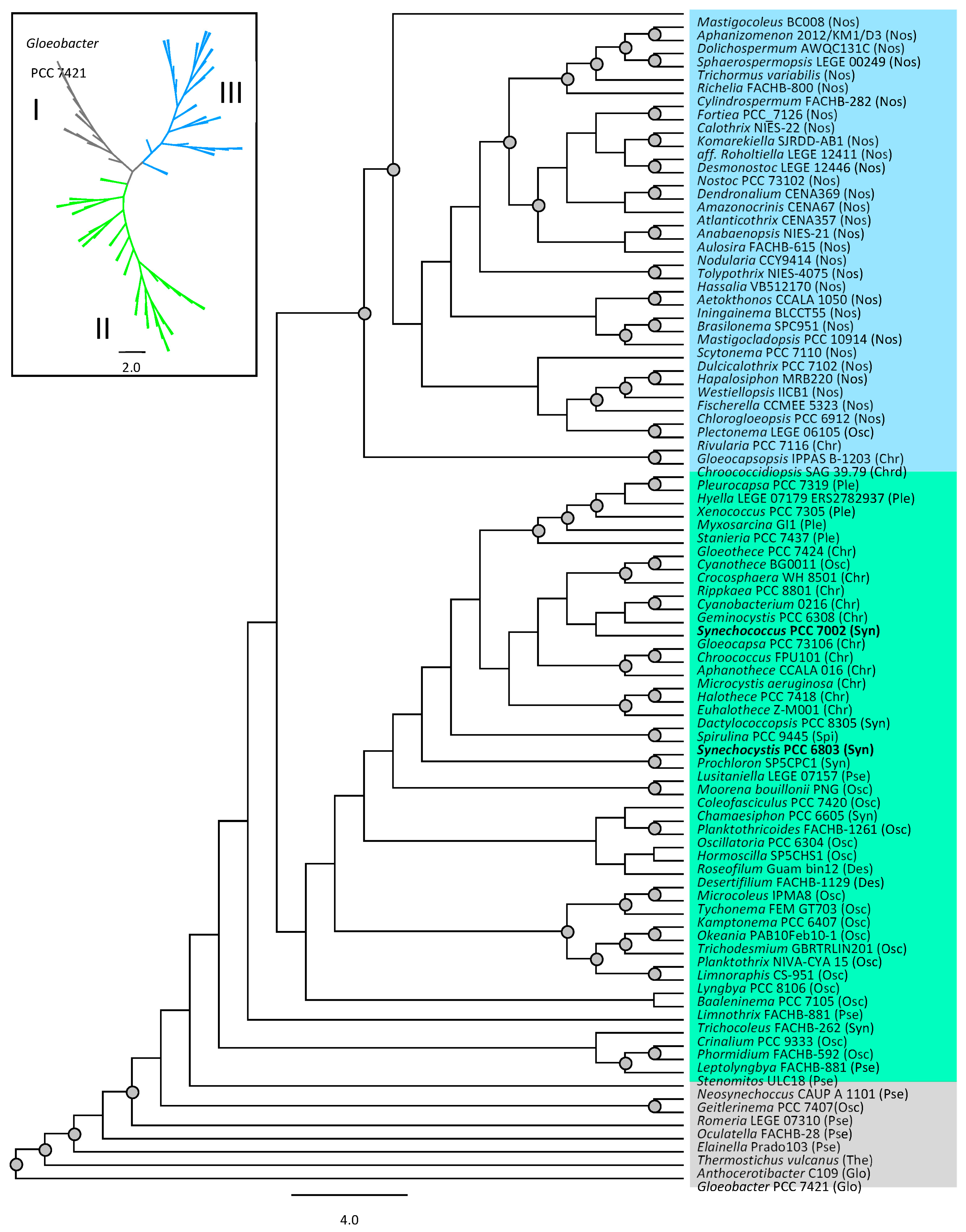
- Moore, K.R.; Magnabosco, C.; Momper, L.; Gold, D.A.; Bosak, T.; Fournier, G.P. An expanded ribosomal phylogeny of cyanobacteria supports a deep placement of plastids. Front. Microbiol. 2019, 10, 1612. [Google Scholar] [CrossRef]
- Komárek, J.; Johansen, J.R.; Šmarda, J.; Strunecký, O. Phylogeny and taxonomy of Synechococcus-like cyanobacteria. Fottea 2020, 20, 171–191. [Google Scholar] [CrossRef]
- Strunecký, O.; Kopejtka, K.; Goecke, F.; Tomasch, J.; Lukavský, J.; Neori, A.; Kahl, S.; Pieper, D.H.; Pilarski, P.; Kaftan, D.; et al. High diversity of thermophilic cyanobacteria in Rupite hot spring identified by microscopy, cultivation, single-cell PCR and amplicon sequencing. Extremophiles 2019, 23, 35–48. [Google Scholar] [CrossRef]
- Strunecký, O.; Ivanova, A.P.; Mareš, J. An updated classification of cyanobacterial orders and families based on phylogenomic and polyphasic analysis. J. Phycol. 2023, 59, 12–51. [Google Scholar] [CrossRef]
- Hirose, Y.; Ohtsubo, Y.; Misawa, N.; Yonekawa, C.; Nagao, N.; Shimura, Y.; Fujisawa, T.; Kanesaki, Y.; Katoh, H.; Katayama, M.; et al. Genome sequencing of the NIES cyanobacteria collection with a focus on the heterocyst-forming clade. DNA Res. 2021, 28, dsab024. [Google Scholar] [CrossRef]
- Ho, M.-Y.; Niedzwiedzki, D.M.; MacGregor-Chatwin, C.; Gerstenecker, G.; Hunter, C.N.; Blankenship, R.E.; Bryant, D.A. Extensive remodeling of the Photosynthetic apparatus alters energy transfer among Photosynthetic complexes when cyanobacteria acclimate to far-red light. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148064. [Google Scholar] [CrossRef]
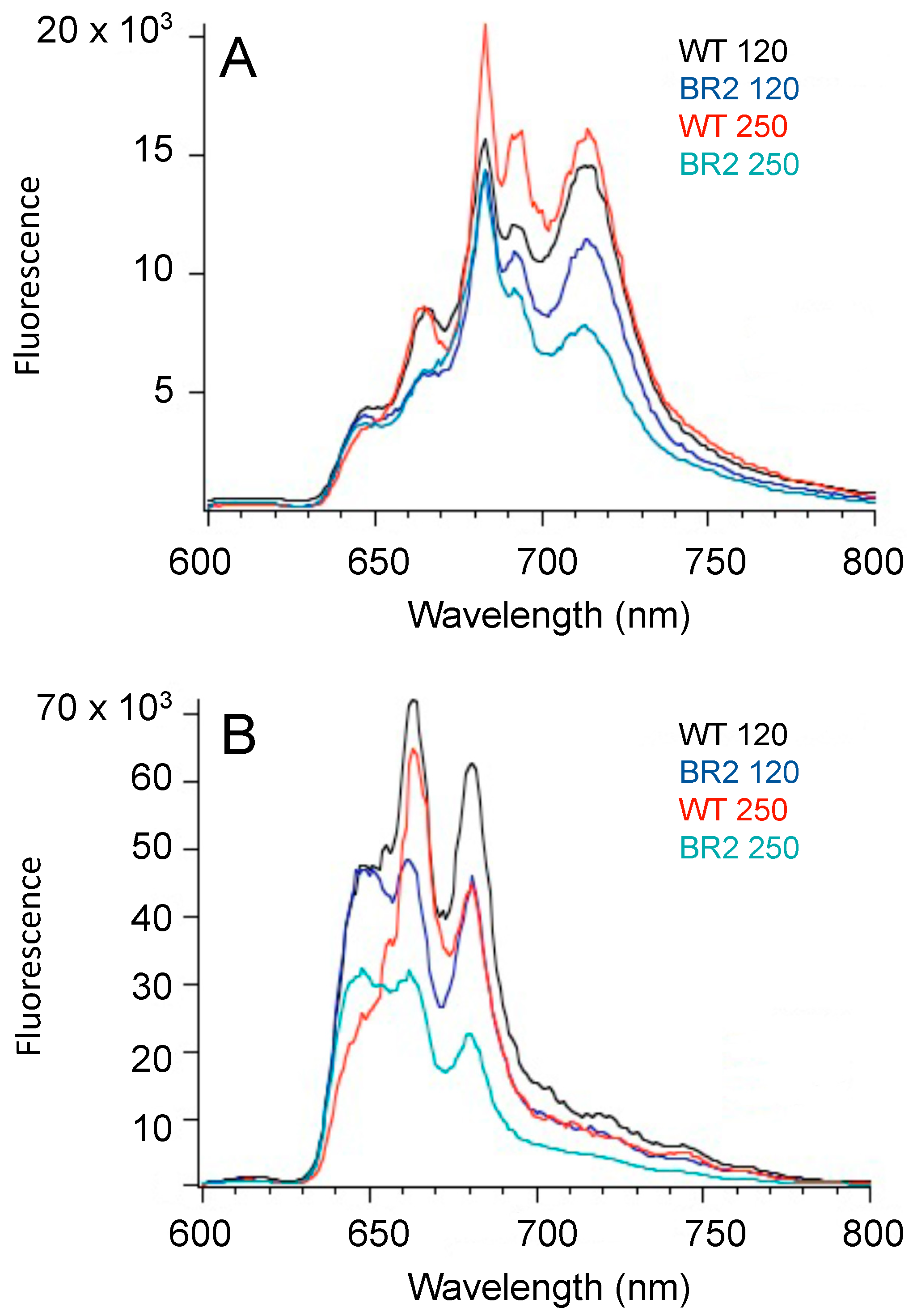
- Gindt, Y.M.; Zhou, J.; Bryant, D.A.; Sauer, K. Spectroscopic studies of phycobilisome subcore preparations lacking key core chromophores: Assignment of excited state energies to the Lcm, β18 and ⍺AP-B chromophores. Biochim. Biophys. Acta 1994, 1186, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Page, L.E.; Liberton, M.; Pakrasi, H.B. Reduction of photoautotrophic productivity in the cyanobacterium Synechocystis sp. strain PCC 6803 by phycobilisome antenna truncation. Appl. Environ. Microbiol. 2012, 78, 6349–6351. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.A.; Lorimier, R.; Guglielmi, G.; Stevens, S.E. Structural and compositional analyses of the phycobilisomes of Synechococcus sp. PCC 7002. Analyses of the wild-type strain and a phycocyanin-less mutant constructed by interposon mutagenesis. Arch. Microbiol. 1990, 153, 550–560. [Google Scholar] [CrossRef]
- Nomura, C.T.; Persson, S.; Shen, G.Z.; Inoue-Sakamoto, K.; Bryant, D.A. Characterization of two cytochrome oxidase operons in the marine cyanobacterium Synechococcus sp PCC 7002: Inactivation of ctaDI affects the PSI:PS II ratio. Photosynth. Res. 2006, 87, 215–228. [Google Scholar] [CrossRef]
- Cassier-Chauvat, C.; Chauvat, F. Responses to oxidative and heavy metal stresses in cyanobacteria: Recent advances. Int. J. Mol. Sci. 2014, 16, 871–886. [Google Scholar] [CrossRef]
- Boden, J.S.; Konhauser, K.O.; Robbins, L.J.; Sánchez-Baracaldo, P. Timing the evolution of antioxidant enzymes in cyanobacteria. Nat. Commun. 2021, 12, 4742. [Google Scholar] [CrossRef]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Asada, K. THE WATER-WATER CYCLE IN CHLOROPLASTS: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Rinalducci, S.; Pedersen, J.Z.; Zolla, L. Generation of reactive oxygen species upon strong visible light irradiation of isolated phycobilisomes from Synechocystis PCC 6803. Biochim. Biophys. Acta-Bioenerg. 2008, 1777, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.S.; Brusslan, J.; Haselkorn, R. Expression of a family of psbA genes encoding a photosystem II polypeptide in the cyanobacterium Anacystis nidulans R2. EMBO J. 1986, 5, 2789–2798. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Sakuragi, Y.; Bryant, D.A.; Dellapenna, D. Tocopherols protect Synechocystis sp. strain PCC 6803 from lipid peroxidation. Plant Physiol. 2005, 138, 1422–1435. [Google Scholar] [CrossRef]
- Sakuragi, Y.; Maeda, H.; Dellapenna, D.; Bryant, D.A. ⍺-Tocopherol plays a role in Photosynth.thesis and macronutrient homeostasis of the cyanobacterium Synechocystis sp. PCC 6803 that is independent of its antioxidant function. Plant Physiol. 2006, 141, 508–521. [Google Scholar] [CrossRef]
- Schluchter, W.M.; Bryant, D.A. Molecular characterization of ferredoxin-NADP+ oxidoreductase in cyanobacteria: Cloning and sequence of the petH gene of Synechococcus sp. PCC 7002 and studies on the gene product. Biochemistry 1992, 31, 3092–3102. [Google Scholar] [CrossRef]
- Cornejo, J.; Willows, R.D.; Beale, S.I. Phytobilin biosynthesis: Cloning and expression of a gene encoding soluble ferredoxin-dependent heme oxygenase from Synechocystis sp. PCC 6803. Plant J. 1998, 15, 99–107. [Google Scholar] [CrossRef]
- Di Rienzi, S.C.; Sharon, I.; Wrighton, K.C.; Koren, O.; Hug, L.A.; Thomas, B.C.; Goodrich, J.K.; Bell, J.T.; Spector, T.D.; Banfield, J.F.; et al. The human gut and groundwater harbor non-Photosynthetic bacteria belonging to a new candidate phylum sibling to cyanobacteria. eLife 2013, 2, e01102. [Google Scholar] [CrossRef]
- Coutinho, F.; Tschoeke, D.A.; Thompson, F.; Thompson, C. Comparative genomics of Synechococcus and proposal of the new genus Parasynechococcus. PeerJ 2016, 4, e1522. [Google Scholar] [CrossRef]
- Falcón, L.I.; Magallón, S.; Castillo, A. Dating the cyanobacterial ancestor of the chloroplast. ISME J. 2010, 4, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Terry, M.J.; Maines, M.D.; Lagarias, J.C. Inactivation of phytochrome-chromophore and phycobiliprotein-chromophore precursors by rat-liver biliverdin reductase. J. Biol. Chem. 1993, 268, 26099–26106. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, B.L.; Franklin, K.A.; Terry, M.J.; Thomas, B.; Jackson, S.D.; Crepeau, M.W.; Lagarias, J.C. Biliverdin reductase-induced phytochrome chromophore deficiency in transgenic tobacco. Plant Physiol. 2001, 125, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Lagarias, D.M.; Crepeau, M.W.; Maines, M.D.; Lagarias, J.C. Regulation of photomorphogenesis by expression of mammalian biliverdin reductase in transgenic Arabidopsis plants. Plant Cell 1997, 9, 675–688. [Google Scholar] [CrossRef]
- Ahmed, F.H.; Mohamed, A.E.; Carr, P.D.; Lee, B.M.; Condic-Jurkic, K.; O’Mara, M.L.; Jackson, C.J. Rv2074 is a novel F420 H2-dependent biliverdin reductase in Mycobacterium tuberculosis. Protein Sci. 2016, 25, 1692–1709. [Google Scholar] [CrossRef]
- Perez-Ortiz, G.; Sidda, J.D.; Peate, J.; Ciccarelli, D.; Ding, Y.; Barry, S.M. Production of copropophyrin III, biliverdin and bilirubin by the rufomycin producer, Streptomyces atratus. Front. Microbiol. 2023, 14, 1092166. [Google Scholar] [CrossRef]
- Frankenberg, N.; Mukougawa, K.; Kohchi, T.; Lagarias, J.C. Functional genomic analysis of the HY2 family of ferredoxin-dependent bilin reductases from oxygenic Photosynthetic organisms. Plant Cell 2001, 13, 965–978. [Google Scholar] [CrossRef]
- Quail, P.H.; Boylan, M.T.; Parks, B.M.; Short, T.W.; Xu, Y.; Wagner, D. Phytochromes: Photosensory perception and signal-transduction. Science 1995, 268, 675–680. [Google Scholar] [CrossRef]
- Ishikawa, K.; Xie, X.; Osaki, Y.; Miyawaki, A.; Numata, K.; Kodama, Y. Bilirubin is produced nonenzymatically in plants to maintain chloroplast redox status. Sci. Adv. 2023, 9, eadh4787. [Google Scholar] [CrossRef]
- Kumagai, A.; Ando, R.; Miyatake, H.; Greimel, P.; Kobayashi, T.; Hirabayashi, Y.; Shimogori, T.; Miyawaki, A. A bilirubin-inducible fluorescent protein from eel muscle. Cell 2013, 153, 1602–1611. [Google Scholar] [CrossRef]
- Pirone, C.; Johnson, J.V.; Quirke, J.M.E.; Priestap, H.A.; Lee, D. Bilirubin present in diverse angiosperms. AoB Plants 2010, 2010, plq020. [Google Scholar] [CrossRef]
- Duanmu, D.; Casero, D.; Dent, R.M.; Gallaher, S.; Yang, W.Q.; Rockwell, N.C.; Martin, S.S.; Pellegrini, M.; Niyogi, K.K.; Merchant, S.S.; et al. Retrograde bilin signaling enables Chlamydomonas greening and phototrophic survival. Proc. Natl. Acad. Sci. USA 2013, 110, 3621–3626. [Google Scholar] [CrossRef] [PubMed]
- Duanmu, D.; Rockwell, N.C.; Lagarias, J.C. Algal light sensing and photoacclimation in aquatic environments. Plant Cell Environ. 2017, 40, 2558–2570. [Google Scholar] [CrossRef] [PubMed]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylidès, C.; Havaux, M. Carotenoid oxidation products are stress signals that mediate gene responses to singlet oxygen in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5535–5540. [Google Scholar] [CrossRef]
- Munné-Bosch, S. Vitamin E function in stress sensing and signaling in plants. Dev. Cell 2019, 48, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Vitek, L.; Hinds, T.D., Jr.; Stec, D.E.; Tiribelli, C. The physiology of bilirubin: Health and disease equilibrium. Trends Mol. Med. 2023, 29, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Vítek, L.; Tiribelli, C. Bilirubin: The yellow hormone? J. Hepatol. 2021, 75, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Creeden, J.F.; Gordon, D.M.; Stec, D.E.; Hinds, T.D., Jr. Bilirubin as a metabolic hormone: The physiological relevance of low levels. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E191–E207. [Google Scholar] [CrossRef]
- Zhang, W.; Willows, R.D.; Deng, R.; Li, Z.; Li, M.; Wang, Y.; Guo, Y.; Shi, W.; Fan, Q.; Martin, S.S.; et al. Bilin-dependent regulation of chlorophyll biosynthesis by GUN4. Proc. Natl. Acad. Sci. USA 2021, 118, e2104443118. [Google Scholar] [CrossRef]
- Vavilin, D.V.; Vermaas, W.F.J. Regulation of the tetrapyrrole biosynthetic pathway leading to heme and chlorophyll in plants and cyanobacteria. Physiol. Plant 2002, 115, 9–24. [Google Scholar] [CrossRef]
- Kobayashi, K.; Masuda, T. Transcriptional regulation of tetrapyrrole biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1811. [Google Scholar] [CrossRef] [PubMed]
- Cackett, L.; Luginbuehl, L.H.; Schreier, T.B.; Lopez-Juez, E.; Hibberd, J.M. Chloroplast development in green plant tissues: The interplay between light, hormone, and transcriptional regulation. New Phytol. 2022, 233, 2000–2016. [Google Scholar] [CrossRef] [PubMed]
- Jan, M.S.; Liu, Z.X.; Rochaix, J.D.; Sun, X.W. Retrograde and anterograde signaling in the crosstalk between chloroplast and nucleus. Front. Plant Sci. 2022, 13, 980237. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.W.U.; Montgomery, B.L. Interdependence of tetrapyrrole metabolism, the generation of oxidative stress and the mitigative oxidative stress response. Redox Biol. 2015, 4, 260–271. [Google Scholar] [CrossRef]
- Holleboom, C.P.; Walla, P.J. The back and forth of energy transfer between carotenoids and chlorophylls and its role in the regulation of light harvesting. Photosynth. Res. 2014, 119, 215–221. [Google Scholar] [CrossRef]
- Brzezowski, P.; Richter, A.S.; Grimm, B. Regulation and function of tetrapyrrole biosynthesis in plants and algae. Biochim. Biophys. Acta-Bioenerg. 2015, 1847, 968–985. [Google Scholar] [CrossRef]
- Larkin, R.M.; Alonso, J.M.; Ecker, J.R.; Chory, J. GUN4, a regulator of chlorophyll synthesis and intracellular signaling. Science 2003, 299, 902–906. [Google Scholar] [CrossRef]
- Jackson, P.J.; Hitchcock, A.; Brindley, A.A.; Dickman, M.J.; Hunter, C.N. Absolute quantification of cellular levels of Photosynth.thesis-related proteins in Synechocystis sp. PCC 6803. Photosynth. Res. 2023, 155, 219–245. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relative PBP Content (%) 1 | μg Chl (OD730 · mL)−1 | Relative Carotenoid Content (%) |
|---|---|---|---|
| WT | 100 | 3.56 0.28 (100%) | 100 |
| BR2 | 32.5 | 1.30 0.40 (36.5%) | 150 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schluchter, W.M.; Babin, C.H.; Liu, X.; Bieller, A.; Shen, G.; Alvey, R.M.; Bryant, D.A. Loss of Biliverdin Reductase Increases Oxidative Stress in the Cyanobacterium Synechococcus sp. PCC 7002. Microorganisms 2023, 11, 2593. https://doi.org/10.3390/microorganisms11102593
Schluchter WM, Babin CH, Liu X, Bieller A, Shen G, Alvey RM, Bryant DA. Loss of Biliverdin Reductase Increases Oxidative Stress in the Cyanobacterium Synechococcus sp. PCC 7002. Microorganisms. 2023; 11(10):2593. https://doi.org/10.3390/microorganisms11102593
Chicago/Turabian StyleSchluchter, Wendy M., Courtney H. Babin, Xindi Liu, Amori Bieller, Gaozhong Shen, Richard M. Alvey, and Donald A. Bryant. 2023. "Loss of Biliverdin Reductase Increases Oxidative Stress in the Cyanobacterium Synechococcus sp. PCC 7002" Microorganisms 11, no. 10: 2593. https://doi.org/10.3390/microorganisms11102593