
Gut Dysbiosis in Children with Cystic Fibrosis: Development, Features and the Role of Gut–Lung Axis on Disease Progression


Abstract
:1. Introduction
2. Gut Microbiome in CF: Development and Influences
2.1. CFTR Dysfunction
2.2. Delivery Method
2.3. Breastfeeding
2.4. Antibiotic Treatment
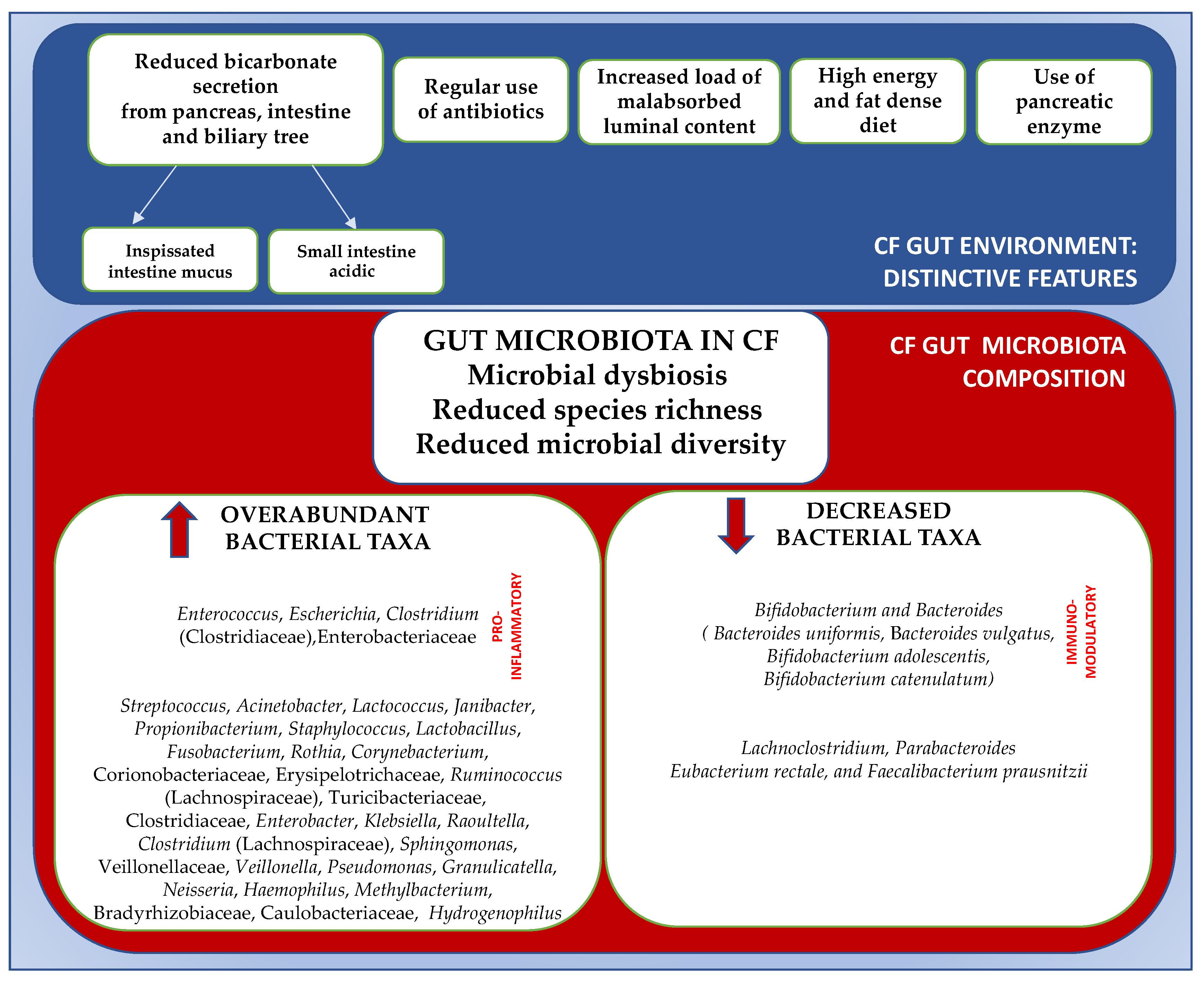
3. Gut Microbiome in Cystic Fibrosis: Composition and Features
4. Gut Microbiota in Cystic Fibrosis: The Gut–lung Axis
5. Gut Microbiota in CF: Possible Influence on Growth and Lung Function
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meoli, A.; Fainardi, V.; Deolmi, M.; Chiopris, G.; Marinelli, F.; Caminiti, C.; Esposito, S.; Pisi, G. State of the Art on Approved Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Modulators and Triple-Combination Therapy. Pharmaceuticals 2021, 14, 928. [Google Scholar] [CrossRef] [PubMed]
- Fink, A.K.; Loeffler, D.R.; Marshall, B.C.; Goss, C.H.; Morgan, W.J. Data that empower: The success and promise of CF patient registries. Pediatr. Pulmonol. 2017, 52, S44–S51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riordan, J.R. Identification of the cystic fibrosis gene: Cloning and characterization of complementary DNA. Science 1989, 245, 1066–1073. [Google Scholar] [CrossRef]
- Riordan, J.R. CFTR function and prospects for therapy. Annu. Rev. Biochem. 2008, 77, 701–726. [Google Scholar] [CrossRef]
- Nielsen, S.; Needham, B.; Leach, S.T.; Day, A.S.; Jaffe, A.; Thomas, T.; Ooi, C.Y. Disrupted progression of the intestinal microbiota with age in children with cystic fibrosis. Sci. Rep. 2016, 6, 24857. [Google Scholar] [CrossRef] [Green Version]
- Rogers, G.B.; Narkewicz, M.R.; Hoffman, R.H. The CF Gastrointestinal Microbiome: Structure and Clinical Impact. Pediatr. Pulmonol. 2016, 51 (Suppl. S44), S35–S44. [Google Scholar] [CrossRef] [Green Version]
- Frati, F.; Salvatori, C.; Incorvaia, C.; Bellucci, A.; Di Cara, G.; Marcucci, F.; Esposito, S. The Role of the Microbiome in Asthma: The Gut-Lung Axis. Int. J. Mol. Sci. 2019, 20, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell. Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Rautava, S.; Luoto, R.; Salnimen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef]
- Berrington, J.E.; Stewart, C.J.; Embleton, N.D.; Cummings, S.R. Gut microbiota in preterm infants: Assessment and relevance to health and disease. Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F286–F290. [Google Scholar] [CrossRef]
- Ballarini, S.; Rossi, G.A.; Principi, N.; Esposito, S. Dysbiosis in Pediatrics Is Associated with Respiratory Infections: Is There a Place for Bacterial-Derived Products? Microorganisms 2021, 9, 448. [Google Scholar] [CrossRef]
- Loman, B.R.; Shrestha, C.L.; Thompson, R.; Kopp, B.T. Age and Environment Exposure Influence the Fecal Bacteriome of Young Childre with Cystic Fibrosis. Pediatr. Pulmonol. 2020, 55, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Meeker, S.M.; Mears, K.S.; Sangwan, N.; Brittnacher, M.J.; Weiss, E.J.; Treuting, P.M.; Tolley, N.; Pope, C.E.; Hager, K.R.; Vo, A.T.; et al. 2020 Jan, CFTR dysregulation drives active selection of the gut microbiome. PLoS Pathog. 2020, 16, e1008251. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.G.; Fouhy, F.; Harrison, M.J. The altered gut microbiota in adults with cystic fibrosis. BMC Microbiol. 2017, 17, 58. [Google Scholar]
- Schippa, S.; Iebba, V.; Santangelo, F.; Gagliardi, A.; De Biase, R.V.; Stamano, A.; Bertasi, S.; Lucarelli, M.; Conte, M.P.; Quattrucci, S. Cystic fibrosis transmembrane conductance regulator (CFTR) allelic viariants relate to shifts in faecal microbiota of cystic fibrosis patients. PLoS ONE 2013, 8, e61176. [Google Scholar] [CrossRef]
- Ooi, C.Y.; Syed, S.A.; Rossi, L.; Garg, M.; Needham, B.; Aviolo, J.; Young, K.; Surette, M.G.; Gonska, T. Impact of CFTR modulation with Ivacaftor on Gut Microbiota and Intestinal Inflammation. Sci. Rep. 2018, 8, 17834. [Google Scholar] [CrossRef] [Green Version]
- Ronan, N.J.; Einarsson, G.G.; Deane, J.; Fouhy, F.; Rea, M.; Hill, C.; Shanahan, F.; Elborn, J.S.; Ross, R.P.; McCarthy, M.; et al. Modulation, microbiota and inflammation in the adult CF gut: A prospective study. J. Cyst. Fibros. 2022, 25, 837–843. [Google Scholar] [CrossRef]
- Antosca, K.M.; Chernikova, D.A.; Price, C.E.; Ruoff, K.L.; Li, K.; Guill, M.F.; Sontag, N.R.; Morrison, H.G.; Hao, S.; Drumm, M.L.; et al. Altered Stool Microbiota of Infants with Cystic Fibrosis Shows a Reduction in Genera Associated with Immune Programming from Birth. J. Bacteriol. 2019, 201, e00274-19. [Google Scholar] [CrossRef] [Green Version]
- Madan, J.C.; Koestler, D.C.; Stanton, B.A.; Davidson, L.; Moulton, L.A.; Housman, M.L.; Moore, J.H.; Guill, M.F.; Morrison, H.G.; Sogin, M.L.; et al. Serial analysys of the gut and respiratory microbiome in CF in infancy: The interaction between intestinal and respiratory tracts and the impact of nutritional exposure. mBio 2012, 3, e00251-12. [Google Scholar] [CrossRef] [Green Version]
- Vernocchi, P.; Del Chierico, F.; Russo, A.; Majo, F.; Rossitto, M.; Valerio, M.; Casadei, L.; La Storia, A.; De Filippis, F.; Rizzo, C.; et al. Gut microbiota signatures in cystic fibrosis: Loss of host CFTR function drives the microbiota enterophenotype. PLoS ONE 2018, 13, e0208171. [Google Scholar] [CrossRef] [Green Version]
- De Freitas, M.B.; Moreira, E.A.M.; Tomio, C.; Moreno, Y.M.F.; Daltoe, F.P.; Barbosa, E.; Neto, N.L.; Buccigrossi, V.; Guarino, A. Altered intestinal microbiota composition, antibiotic therapy and intestinal inflammation in children and adolescents with cystic fibrosis. PLoS ONE 2018, 13, e0198457. [Google Scholar] [CrossRef]
- Kristensen, M.; Prevaes, S.M.P.J.; Kalkman, G.; Tramper-Stranders, G.A.; Hasrat, R.; de Winter-de Groot, K.M.; Janseen, H.M.; Tiddens, H.A.; van Westreenen, M.; Sanders, E.A.M.; et al. Development of the gut microbiota in early life: The impact of cistic fibrosis and antibiotic treatment. J. Cyst. Fibros. 2020, 19, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Yatsuneko, T.; Rey, F.E.; Manary, M.J. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albenberg, L.; Kelsen, J. Advances in Gut Microbiome Research and Relevance to Pediatric Diseases. J. Pediatr. 2016, 178, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Wang, A.C.; Parikh, I.; Green, S.J.; Hoffman, J.D.; Chlipala, G.; Murphy, M.P.; Sokola, B.S.; Bauer, B.; Hartz, A.M.S.; et al. Ketogenic diet enhances neurovascular function with altered gut microbiome in young healty mice. Sci. Rep. 2018, 8, 6670. [Google Scholar] [CrossRef] [Green Version]
- Madan, J.C.; Hoen, A.G.; Lundgren, S.N.; Farzan, S.F.; Cottingham, K.L.; Morrison, H.G.; Sogin, M.L.; Li, H.; Moore, J.H.; Karagas, M.R. Association of Cesarean delivery and formula supplementation with the intestinal microbiome of 6-week-old infants. JAMA Paediatr. 2016, 170, 212–219. [Google Scholar] [CrossRef]
- Esposito, S.; Ballarini, S.; Argentiero, A.; Ruggiero, L.; Rossi, G.A.; Principi, N. Microbiota profiles in pre-school children with respiratory infections: Modifications induced by the oral bacterial lysate OM-85. Front. Cell. Infect. Microbiol. 2022, 12, 1088. [Google Scholar] [CrossRef]
- Fouhy, F.; Ronan, N.J.; O’Sullivan, O.; McCarthy, Y.; Walsh, A.M.; Murphy, D.M.; Daly, M.; Flanagan, E.T.; Fleming, C.; McCarthy, M.; et al. A pilot study demonstrating the altered gut microbiota functionality in stable adults with Cystic Fibrosis. Sci. Rep. 2017, 7, 6685. [Google Scholar] [CrossRef] [Green Version]
- Duytschaever, G.; Huys, G.; Bekaert, M.; Boulanger, L.; De Boeck, K.; Vandamme, P. Cross-Sectional and Longitudinal Composition of Prenominant Fecal Microbiota Composition of a Group of Pediatric Patients with Cystic Fibrosis and Their Healty Siblings. Appl. Environ. Microbiol. 2011, 77, 8015–8024. [Google Scholar] [CrossRef] [Green Version]
- Sommer, M.O.; Church, G.M.; Dantas, G. The human microbiome harbors a diverse reservoiur of antibiotic resistance genes. Virulence 2010, 1, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Duytschaever, G.; Huys, G.; Boulanger, L.; De Boeck, K.; Vandamme, P. Amoxicillin–clavulanic acid resistance in fecal Enterobacteriaceae from patients with cystic fibrosis and healthy siblings. J. Cyst. Fibros. 2013, 12, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.L.; Leong, L.E.X.; Sims, S.K.; Keating, R.L.; Papanicolas, L.E.; Richard, A.; Mobegi, F.M.; Wesselingh, S.; Burr, L.D.; Rogers, G.B. The cystic fibrosis gut as a potential source of multidrug resistant pathogens. J. Cyst. Fibros. 2021, 20, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Thavamani, A.; Salem, I.; Sferra, T.J.; Sankararaman, S. Impact of Altered Gut Microbiota and Its Metabolites in Cystic Fibrosis. Metabolites 2021, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.R.; Pope, C.E.; Hayden, H.S.; Heltshe, S.; Levy, R.; McNamara, S.; Jacobs, M.A.; Rohmer, L.; Radey, M.; Ramsey, B.W.; et al. Escherichia coli Dysbiosis Correlates with Gastrointestinal Dysfunction in Children with Cystic Fibrosis. Clin. Infect. Dis. 2014, 58, 396–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanlan, P.D.; Buckling, A.; Kong, W.; Wild, Y.; Lynch, S.V.; Harrison, F. Gut dysbiosis in cystic fibrosis. J. Cyst. Fibros. 2012, 11, 454–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruzzese, E.; Callegari, M.L.; Raia, V.; Viscovo, S.; Scotto, R.; Ferrari, S.; Morelli, L.; Buccigrossi, V.; Lo Vecchio, A.; Ruberto, E.; et al. Disrupted Intestinal Microbiota and Intestinal Inflammation in Children with Cystic Fibrosis and Its Restoration with Lactobacillus GG: A Randomised Clinical Trial. PLoS ONE 2014, 9, e87796. [Google Scholar] [CrossRef] [Green Version]
- Coffey, M.J.; Nielsen, S.; Wemheuer, B.; Kaakoush, N.O.; Garg, M.; Needham, B.; Pickford, R.; Jaffe, A.; Thomas, T.; Ooi, C.Y. Gut microbiota in children with cystic fibrosis: A taxonomic and functional dysbiosis. Sci. Rep. 2019, 9, 18593. [Google Scholar] [CrossRef] [Green Version]
- Price, C.E.; O’Toole, G.A. The gut-lung axis in cystic fibrosis. J. Bacteriol. 2021, 203, e00311-21. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Wang, N.; Tan, H.Y.; Zhang, Z.; Feng, Y. The Cross-Talk Between Gut Microbiota and Lungs in Common Lung Diseases. Front. Microbiol. 2020, 11, 301. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodriguez, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain faffy acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Françoise, A.; Héry-Arnaud, G. The Microbiome in Cystic Fibrosis Pulmonary Disease. Genes 2020, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.; Gaggar, A.; Bruscia, E.; Hector, A.; Marcos, V.; Jung, A.; Greene, C.; McElvaney, G.; Mall, M.; Döring, G. Innate immunity in cystic fibrosis lung disease. J. Cyst. Fibros. 2012, 11, 363–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoen, A.G.; Li, J.; Moulton, L.A.; O’Toole, G.A.; Housman, M.L.; Koestler, D.C.; Guill, M.F.; Moore, J.H.; Hibberd, P.L.; Morrison, H.G.; et al. Association between gut microbial colonization in early life and respiratory outcomes in cystic fibrosis. J. Pediatr. 2015, 167, 138–147.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkstén, B.; Naaber, P.; Sepp, E.; Mikelsaar, M. The intestinal microflora in allergic Estonian and Swedish 2-year-old children. Clin. Exp. Allergy 1999, 29, 342–346. [Google Scholar] [CrossRef]
- Kalliomäki, M.; Kirjavainen, P.; Eerola, E.; Kero, P.; Salminen, S.; Isolauri, E. Distinct patterns of neonatal gut microflora in infants in whom atopy was and was not developing. J. Allergy Clin. Immunol. 2001, 107, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Duytschaever, G.; Huys, G.; Bekaert, M. Dysbiosis of bifidobacteria and Clostridium cluster XIVa in the cystic fibrosis fecal microbiota. J. Cyst. FIbros. 2013, 12, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Testa, I.; Mariotti Zani, E.; Cunico, D.; Torelli, L.; Grandinetti, R.; Fainardi, V.; Pisi, G.; Principi, N. Probiotics Administration in Cystic Fibrosis: What Is the Evidence? Nutrients 2022, 14, 3160. [Google Scholar] [CrossRef]
- Grier, A.; McDavid, A.; Wang, B.; Qiu, X.; Java, J.; Bandyopadhyay, S.; Yang, H.; Holden-Wiltse, J.; Kessler, H.A.; Gill, A.L.; et al. Neonatal gut and respiratory microbiota: Coordinated development through time and space. Microbiome 2018, 6, 193. [Google Scholar] [CrossRef] [Green Version]
- Hulzebos, E.H.; Bomhof-Roordink, H.; van de Weert-van Leeuwen, P.B.; Twisk, J.W.; Arets, H.G.; van der Ent, C.K.; Takken, T. Prediction of mortality in adolescents with cystic fibrosis. Med. Sci. Sport. Exerc. 2014, 46, 2047–2052. [Google Scholar] [CrossRef]
- Hayden, H.S.; Eng, A.; Pope, C.E.; Brittnacher, M.J.; Vo, A.T.; Weiss, E.J.; Hager, K.R.; Martin, B.D.; Leung, D.H.; Heltshe, S.L.; et al. Fecal dysbiosis in infants with cystic fibrosis is associated with early linear growth failure. Nat. Med. 2020, 26, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Gough, E.K.; Stephens, D.A.; Moodie, E.E.; Prendergast, A.J.; Stoltzfus, R.J.; Humphrey, J.H.; Manges, A.R. Linear growth faltering in infants is associated with Acidaminococcus sp. and community-level changes in the gut microbiota. Microbiome 2015, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werlin, S.L.; Benuri-Silbiger, I.; Kerem, E.; Adler, S.N.; Goldin, E.; Zimmerman, J.; Malka, N.; Cohen, L.; Armoni, S.; Yatzkan-Israelit, Y.; et al. Evidence of intestinal inflammation in patients with cystic fibrosis. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 304–308. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Factors Involved | Reference | Study Evidence | Taxa Imbalance |
|---|---|---|---|
| Age | Loman et al. [12] |
|
|
| CFTR dysfunction and CFTR modulators | Meeker et al. [13] | CFTR dysfunction actively modulates and selects gut microbiome in mice | |
| Burke et al. [14] | No significant differences in species richness or microbial diversity between the CF cohort with class 1–3 mutations and other classes |
| |
| Schippa et al. [15] |
| ||
| Ooi et al. [16] | No significant difference in alpha and beta diversities 6 months after starting ivacaftor |
| |
| Ronan et al. [17] | No significant change in gut microbiota diversity and richness after a year of treatment with ivacaftor | ||
| Delivery method | Loman et al. [12] | ↑ alpha diversity in children born by C-section |
|
| Antosca et al. [18] | Delivery mode not affecting significantly gut microbiota of CF infants | ||
| Breastfeeding | Loman et al. [12] | Similar alpha and beta diversities in formula-fed vs. breastfed or mixed formula- and breastfed |
|
| Madan et al. [19] |
| ||
| Antibiotics use | Vernocchi et al. [20] |
|
|
| de Freitas et al. [21] |
| ||
| Burke et al. [14] | Significant negative correlation between the number of IV antibiotic courses and gut microbiota diversity |
| |
| Kristensen et al. [22] | Independent association between antibiotic treatment (mainly co-trimoxazole) and lower alpha diversity in CF infants |
|
| Gut-Lung Axys: Study Evidence in Children with Cystic Fibrosis | ||
|---|---|---|
| Authors | Study Population | Study Evidence |
| Madan et al. [9] | 7 CF patients from birth to 9-21 months of age |
|
| Hoen et al. [43] | 13 CF children from birth to 34 months of age |
|
| Antosca et al. [18] | 21 CF infants and 409 controls sampled between 6 weeks and 12 months of age |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Testa, I.; Crescenzi, O.; Esposito, S. Gut Dysbiosis in Children with Cystic Fibrosis: Development, Features and the Role of Gut–Lung Axis on Disease Progression. Microorganisms 2023, 11, 9. https://doi.org/10.3390/microorganisms11010009
Testa I, Crescenzi O, Esposito S. Gut Dysbiosis in Children with Cystic Fibrosis: Development, Features and the Role of Gut–Lung Axis on Disease Progression. Microorganisms. 2023; 11(1):9. https://doi.org/10.3390/microorganisms11010009
Chicago/Turabian StyleTesta, Ilaria, Oliviero Crescenzi, and Susanna Esposito. 2023. "Gut Dysbiosis in Children with Cystic Fibrosis: Development, Features and the Role of Gut–Lung Axis on Disease Progression" Microorganisms 11, no. 1: 9. https://doi.org/10.3390/microorganisms11010009