
Evidence on the Preventive Effects of the Postbiotic Derived from Cow’s Milk Fermentation with Lacticaseibacillus paracasei CBA L74 against Pediatric Gastrointestinal Infections
 , and
, and
Abstract
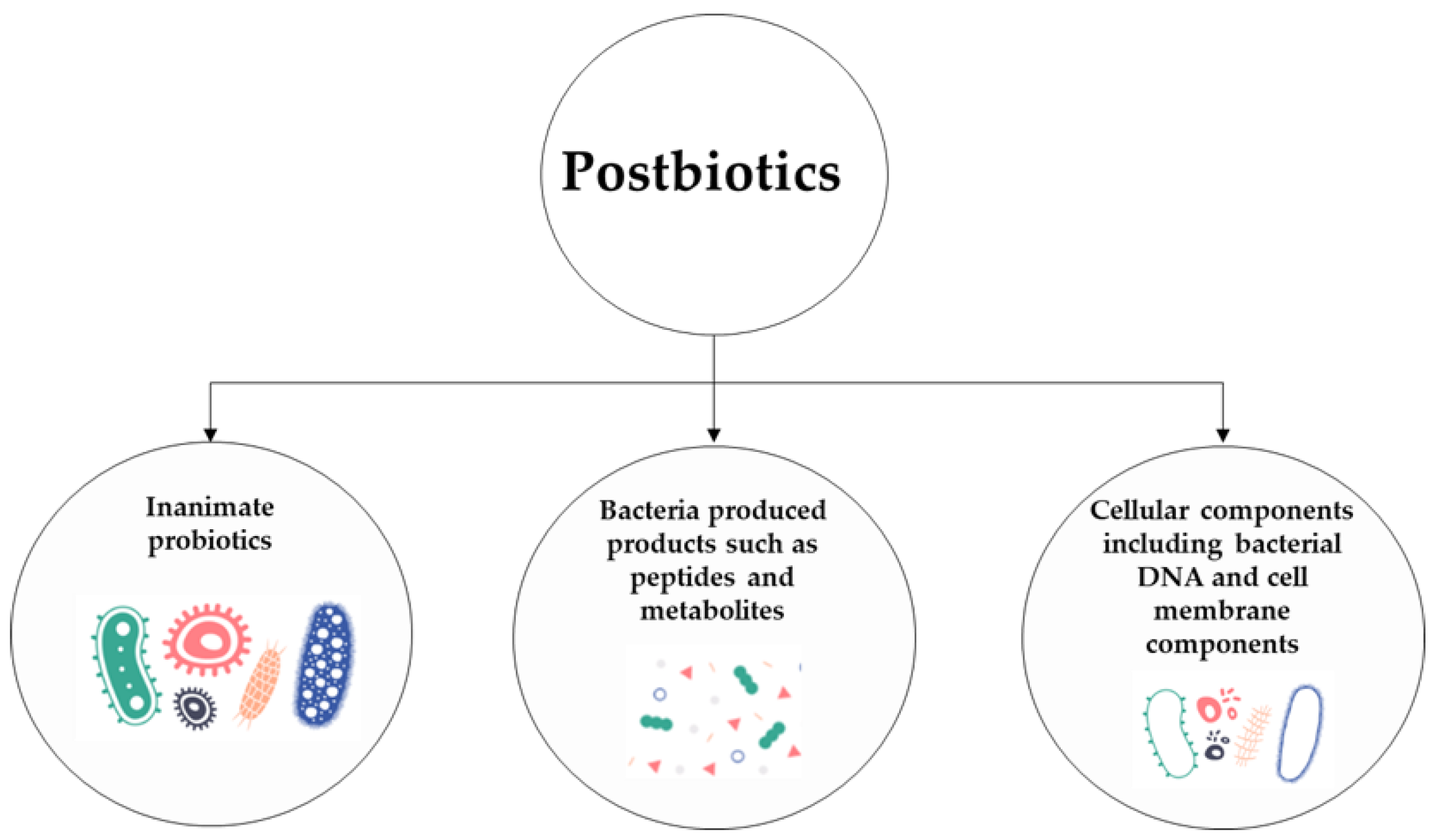
:1. Introduction
|
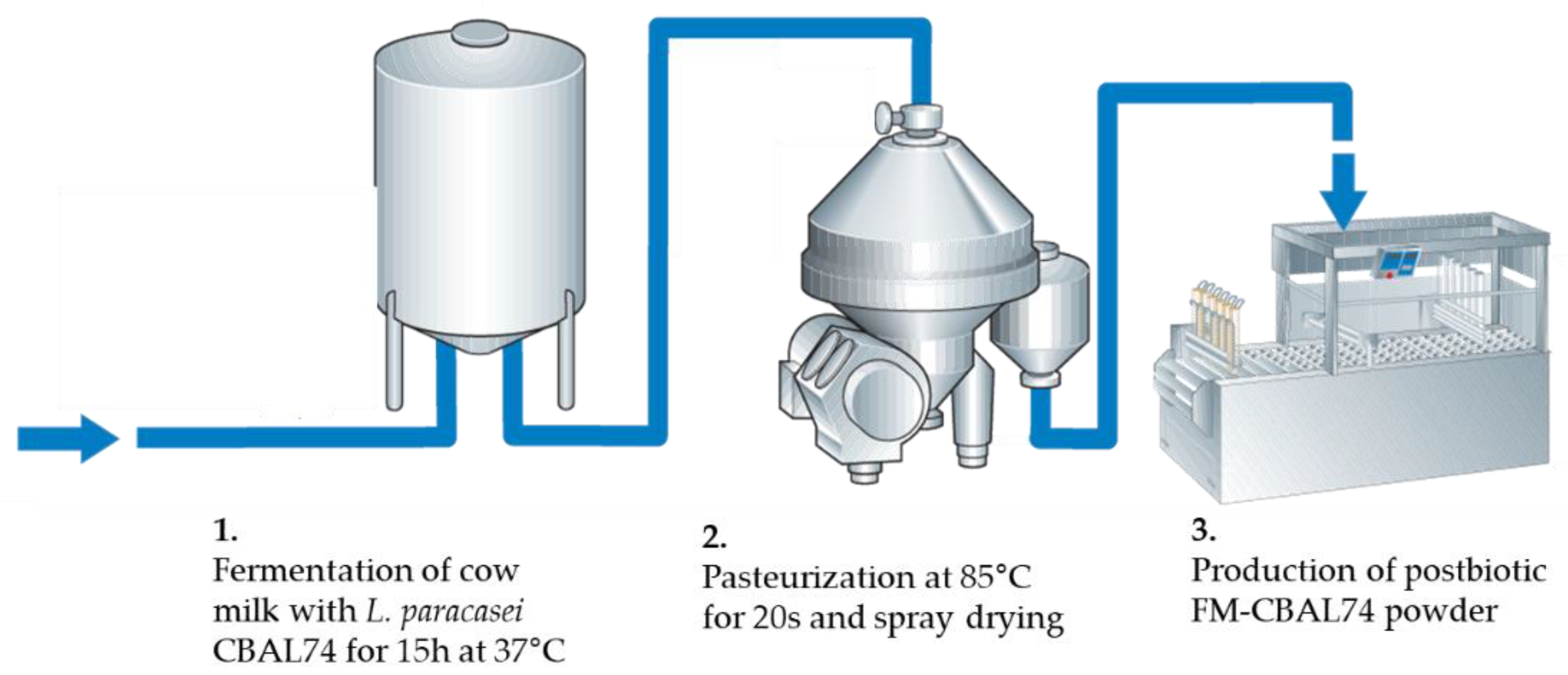
2. The Postbiotic Derived from Cow Milk Fermentation with the Probiotic L. paracasei CBAL74
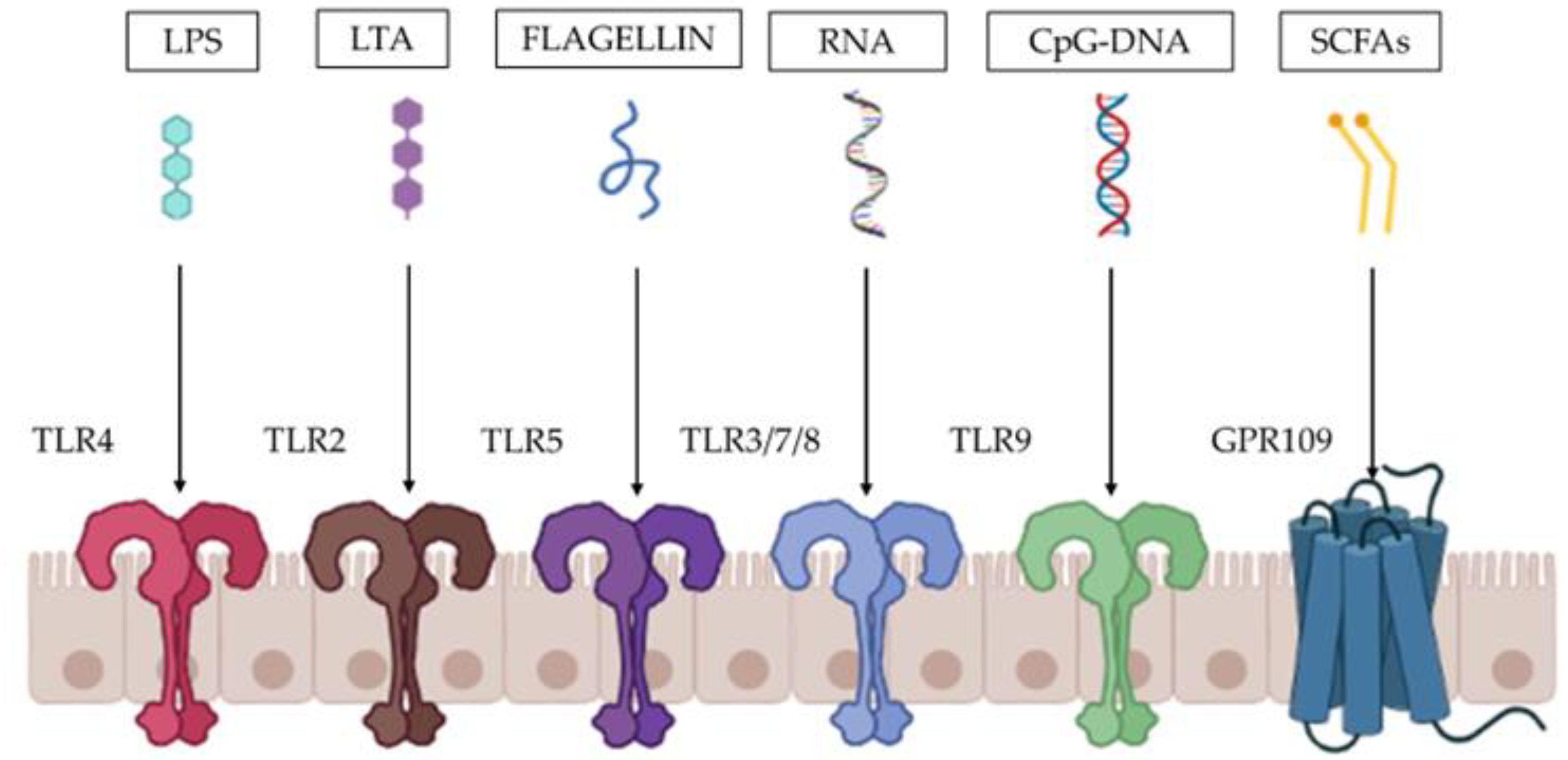
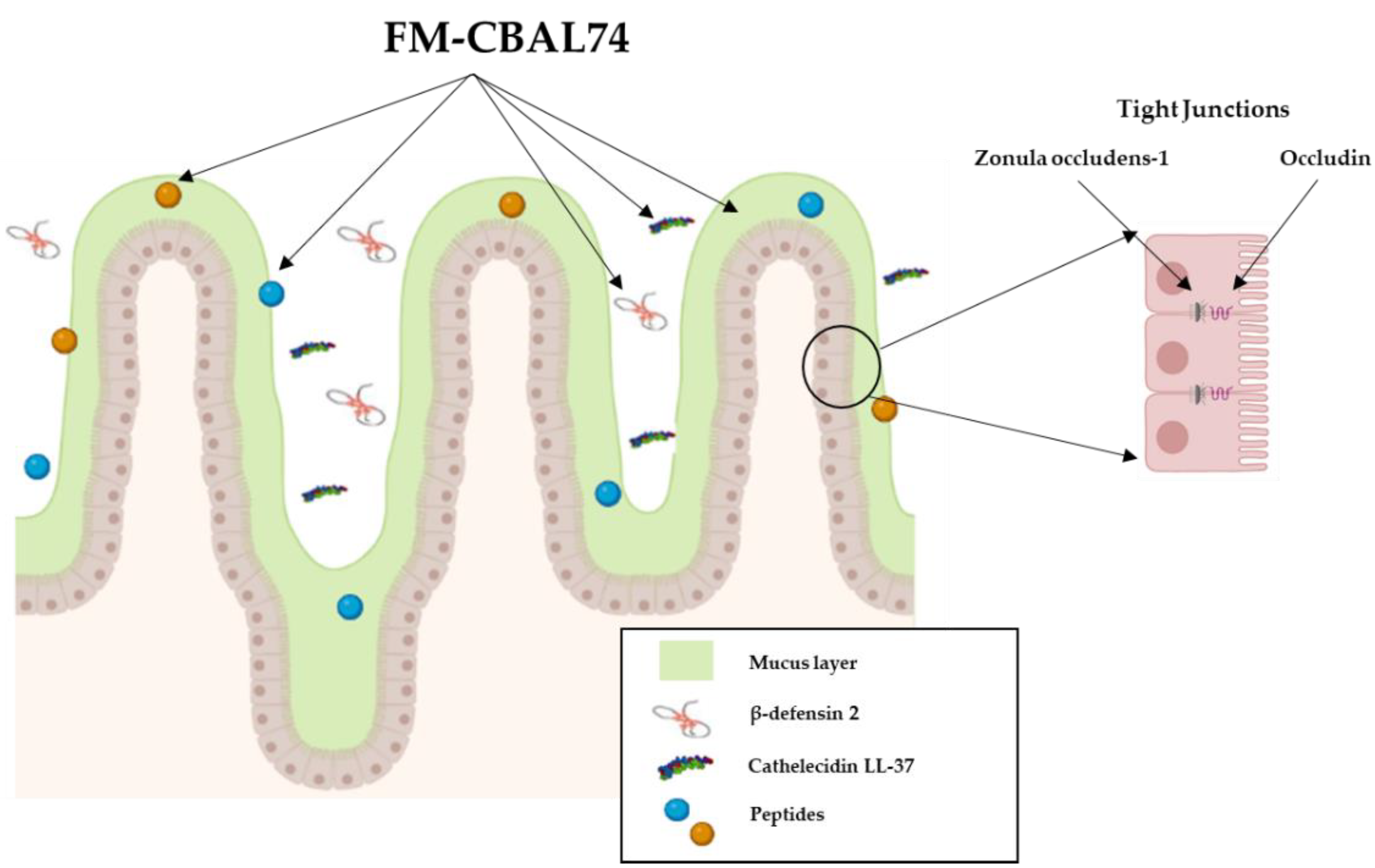
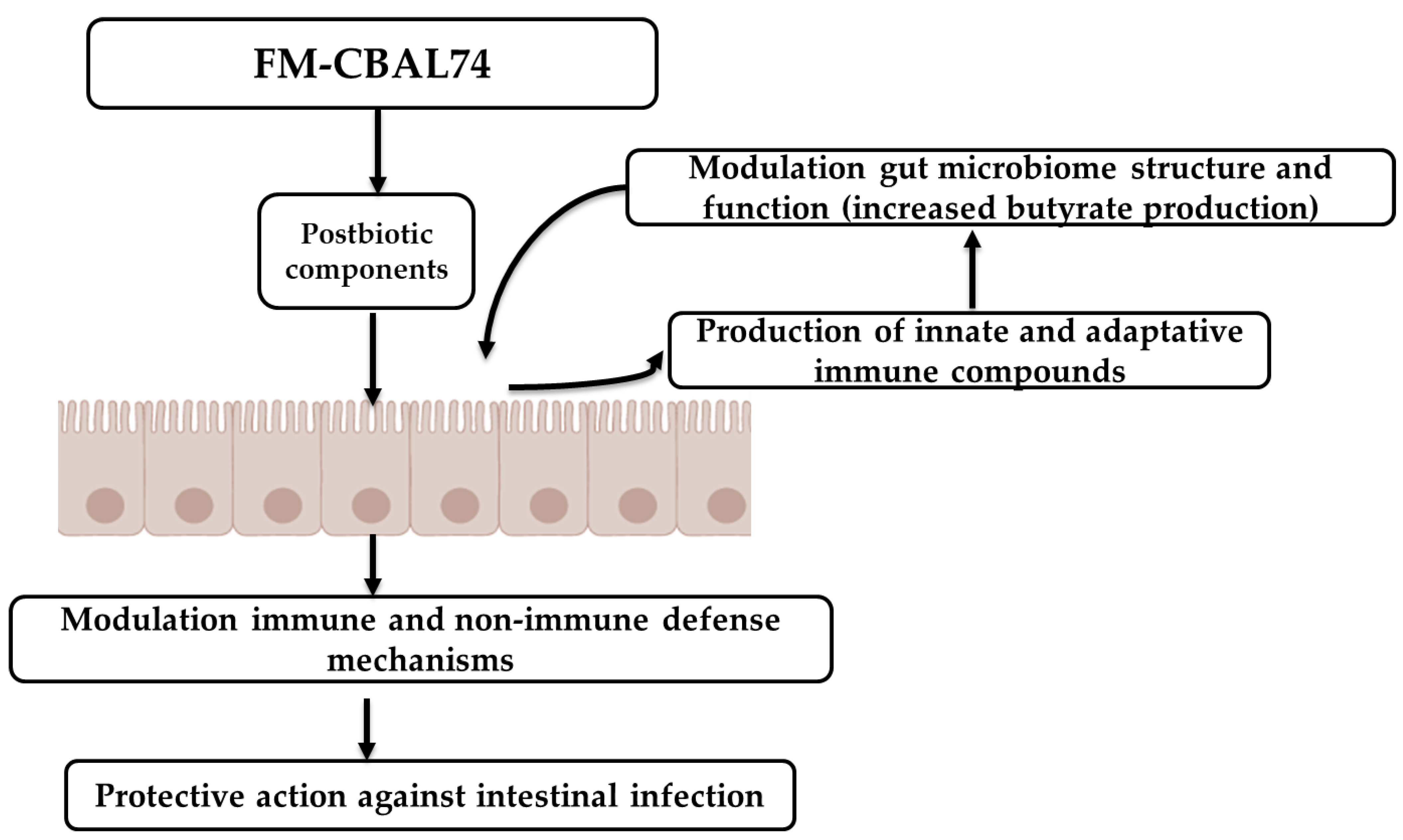
3. Preclinical Evidence on the Mechanisms of Action Elicited by FM-CBAL74
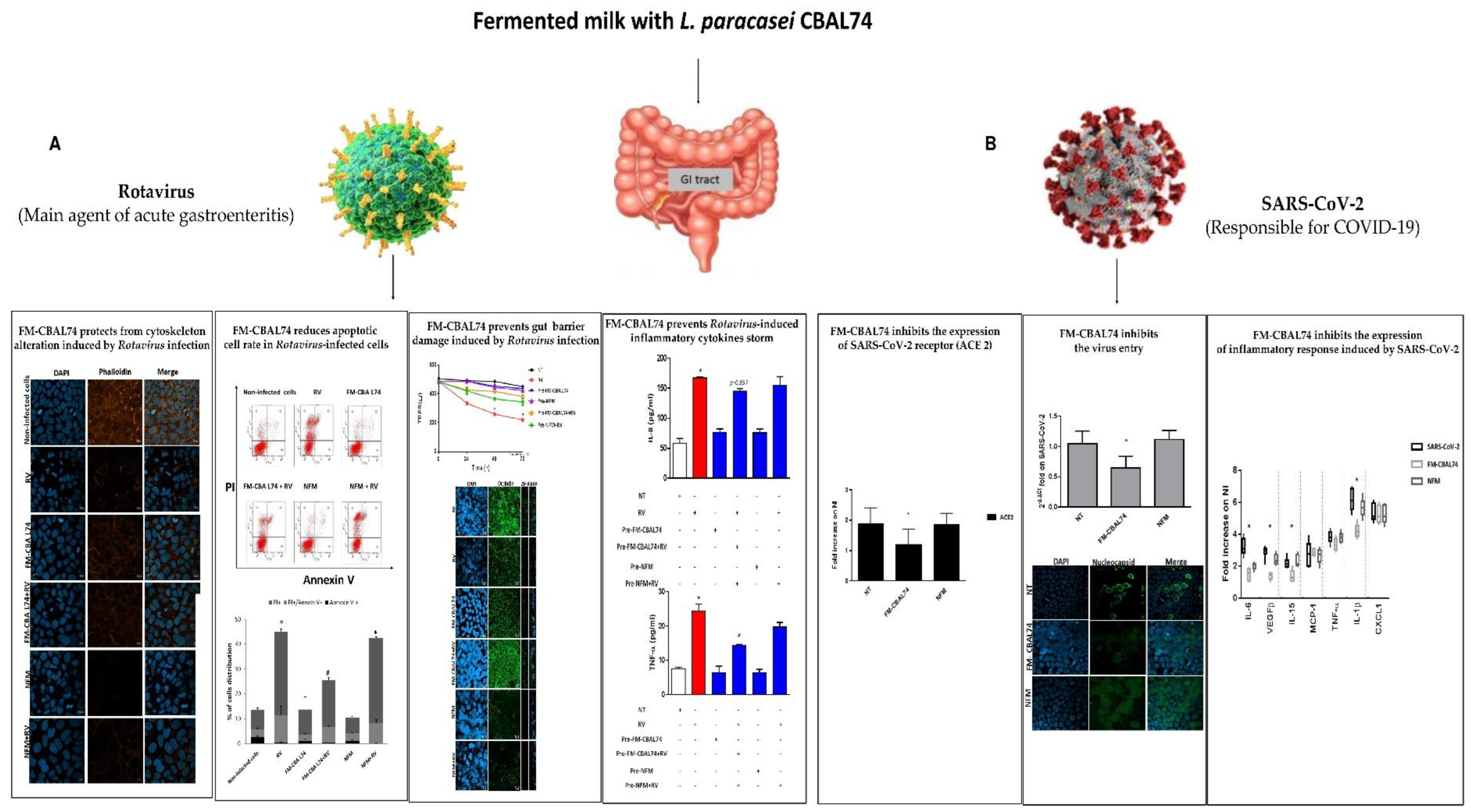
4. Preclinical Evidence on the FM-CBAL74 Effects against Intestinal Viral Infections
5. Clinical Evidence on the Preventive Action of FM-CBAL74 against Pediatric Intestinal Infections
5.1. Clinical Data
5.2. Safety
5.3. Fecal Immune Biomarkers and Microbiome Data
5.4. Limitations of the Available Clinical Data
6. Toward a Research Agenda
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguilar-Toalá, J.E.; Arioli, S.; Behare, P.; Belzer, C.; Berni Canani, R.; Chatel, J.M.; D’Auria, E.; de Freitas, M.Q.; Elinav, E.; Esmerino, E.A.; et al. Postbiotics—When simplification fails to clarify. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 825–826. [Google Scholar] [CrossRef]
- Morniroli, D.; Vizzari, G.; Consales, A.; Mosca, F.; Giannì, M.L. Postbiotic Supplementation for Children and Newborn’s Health. Nutrients 2021, 13, 781. [Google Scholar] [CrossRef]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and Their Potential Applications in Early Life Nutrition and Beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [Green Version]
- Warshakoon, H.J.; Hood, J.D.; Kimbrell, M.R.; Malladi, S.; Wu, W.Y.; Shukla, N.M.; Agnihotri, G.; Sil, D.; David, S.A. Potential adjuvantic properties of innate immune stimuli. Hum. Vaccin. 2009, 5, 381–394. [Google Scholar] [CrossRef]
- Chandler, C.E.; Ernst, R.K. Bacterial lipids: Powerful modifiers of the innate immune response. F1000Research 2017, 6, F1000 Faculty Rev-1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [Green Version]
- Mantziari, A.; Salminen, S.; Szajewska, H.; Malagon-Rojas, J.-N. Postbiotics against Pathogens Commonly Involved in Pediatric Infection Disease. Microorganisms 2020, 8, 1510. [Google Scholar] [CrossRef]
- Ricke, S.C. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics-A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizonsin microbial biotherapy and functional foods. Microb. Cell Fact. 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Paparo, L.; Aitoro, R.; Nocerino, R.; Fierro, C.; Bruno, C.; Berni Canani, R. Direct effects of fermented cow's milk product with Lactobacillus paracasei CBA L74 on human enterocytes. Benef. Microbes 2018, 9, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Bruno, C.; Paparo, L.; Pisapia, L.; Romano, A.; Cortese, M.; Punzo, E.; Berni Canani, R. Protective effects of the postbiotic deriving from cow’s milk fermentation with L. paracasei CBA L74 against Rotavirus infection in human enterocytes. Sci. Rep. 2022, 12, 6268. [Google Scholar] [CrossRef] [PubMed]
- Paparo, L.; Bruno, C.; Ferrucci, V.; Punto, E.; Viscardi, M.; Fusco, G.; Cerino, P.; Romano, A.; Zollo, M.; Berni Canani, R. Protective effects elicited by cow milk fermented with L. paracasei CBAL74 against SARS-CoV-2 infection in human enterocytes. J. Funct. Foods. 2021, 87, 104787. [Google Scholar] [CrossRef] [PubMed]
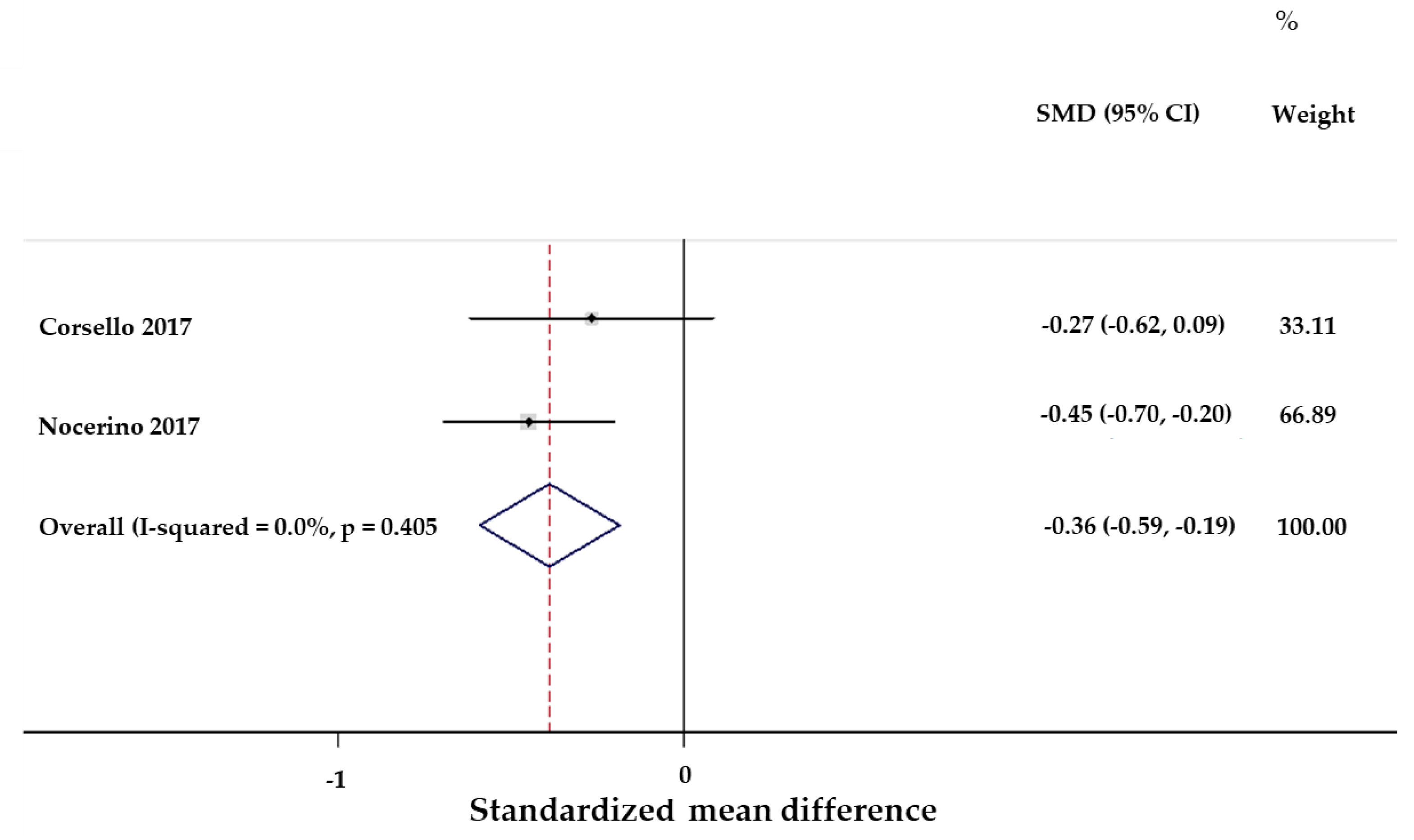
- Nocerino, R.; Paparo, L.; Terrin, G.; Pezzella, V.; Amoroso, A.; Cosenza, L.; Cecere, G.; De Marco, G.; Micillo, M.; Albano, F.; et al. Cow’s milk and rice fermented with Lactobacillus paracasei CBA L74 prevent infectious diseases in children: A randomized controlled trial. Clin. Nutr. 2017, 36, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Corsello, G.; Carta, M.; Marinello, R.; Picca, M.; De Marco, G.; Micillo, M.; Ferrara, D.; Vigneri, P.; Cecere, G.; Ferri, P.; et al. Preventive Effect of Cow's Milk Fermented with Lactobacillus paracasei CBA L74 on Common Infectious Diseases in Children: A Multicenter Randomized Controlled Trial. Nutrients 2017, 9, 669. [Google Scholar] [CrossRef] [PubMed]
- Roggero, P.; Liotto, N.; Pozzi, C.; Braga, D.; Troisi, J.; Menis, C.; Giannì, M.L.; Berni Canani, R.; Paparo, L.; Nocerino, R.; et al. Analysis of immune, microbiota and metabolome maturation in infants in a clinical trial of Lactobacillus paracasei CBA L74-fermented formula. Nat. Commun. 2020, 1, 2703. [Google Scholar] [CrossRef] [PubMed]
- Zagato, E.; Mileti, E.; Massimiliano, L.; Fasano, F.; Budelli, A.; Penna, G.; Rescigno, M. Lactobacillus paracasei CBA L74 metabolic products and fermented milk for infant formula have anti-inflammatory activity on dendritic cells in vitro and protective effects against colitis and an enteric pathogen in vivo. PLoS ONE 2014, 9, e87615. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; Raev, S.A.; Chepngeno, J.; Maingo, A.O.; Guo, Y.; Saif, L.; Vlasova, A.N. Rotavirus Interactions with Host Intestinal Epithelial Cells. Front. Immunol. 2022, 12, 793841. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; De Filippis, F.; Nocerino, R.; Laiola, M.; Paparo, L.; Calignano, A.; De Caro, C.; Coretti, L.; Chiariotti, L.; Gilbert, J.A.; et al. Specific Signatures of the Gut Microbiota and Increased Levels of Butyrate in Children Treated with Fermented Cow’s Milk Containing Heat-Killed Lactobacillus paracasei CBA L74. Appl. Environ. Microbiol. 2017, 83, e01206–e01217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calame, W.; van Olderen, D.; Calabretta, V.; Bottari, L.; Paparo, L.; Bruno, C.; Carucci, L.; Voto, L.; Coppola, S.; Budelli, A. Baseline Concentrations of Various Immune Biomarkers Determine Their Increase after Consumption of a Postbiotic Based on Cow’s Milk Fermented with Lactobacillus paracasei CBA L74 in Both Newborns and Young Children. Appl. Sci. 2022, 12, 2009. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preclinical Evidence | ||
|---|---|---|
| Outcomes | Effects | References |
| Modulation of the major defense mechanisms against intestinal infections | Stimulation of enterocytes cell growth and differentiation, tight-junctions proteins expression, mucus layer thickness, and innate immunity peptides production. | [11] |
| Anti-inflammatory activity | Inhibition of pro-inflammatory cytokines release, and modulation of Toll-like receptor 2 and 4, transcriptional factor nuclear factor kappa B1, and trefoil factor 3 expression in enterocytes and dendritic cells. | [11,12,17] |
| Anti-viral action | Protection against Rotavirus-induced gut barrier damage, oxidative stress, and pro-inflammatory cytokines release. Protection against SARS-CoV-2 entry into human enterocytes, and pro-inflammatory cytokines release | [12,13] |
| Clinical evidence | ||
| Protection against pediatric acute gastroenteritis | Significant reduction of the rate of children presenting ≥ 1 episodes of acute gastroenteritis and of the total number of acute gastroenteritis episodes. | [14,15] |
| Stimulation of innate and adaptative immunity | Increased intestinal production of α- and β-defensins, cathelicidin LL-37, and secretory immunoglobulin A. | [14,15,16] |
| Modulation of gut microbiome | Beneficial modulation of gut microbiome structure and function in formula fed neonates, infants, and children with increased abundance of healthy butyrate-producer bacteria. | [16,19] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oglio, F.; Bruno, C.; Coppola, S.; De Michele, R.; Masino, A.; Carucci, L. Evidence on the Preventive Effects of the Postbiotic Derived from Cow’s Milk Fermentation with Lacticaseibacillus paracasei CBA L74 against Pediatric Gastrointestinal Infections. Microorganisms 2023, 11, 10. https://doi.org/10.3390/microorganisms11010010
Oglio F, Bruno C, Coppola S, De Michele R, Masino A, Carucci L. Evidence on the Preventive Effects of the Postbiotic Derived from Cow’s Milk Fermentation with Lacticaseibacillus paracasei CBA L74 against Pediatric Gastrointestinal Infections. Microorganisms. 2023; 11(1):10. https://doi.org/10.3390/microorganisms11010010
Chicago/Turabian StyleOglio, Franca, Cristina Bruno, Serena Coppola, Roberta De Michele, Antonio Masino, and Laura Carucci. 2023. "Evidence on the Preventive Effects of the Postbiotic Derived from Cow’s Milk Fermentation with Lacticaseibacillus paracasei CBA L74 against Pediatric Gastrointestinal Infections" Microorganisms 11, no. 1: 10. https://doi.org/10.3390/microorganisms11010010