
iChip-Inspired Isolation, Bioactivities and Dereplication of Actinomycetota from Portuguese Beach Sediments

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. Identification of the Strain’s Phylogeny and Detection of Secondary-Metabolite-Associated Genes
2.3. Strain Fermentation and Extraction
2.4. Antimicrobial Screening
2.5. Dereplication of Extracts
3. Results
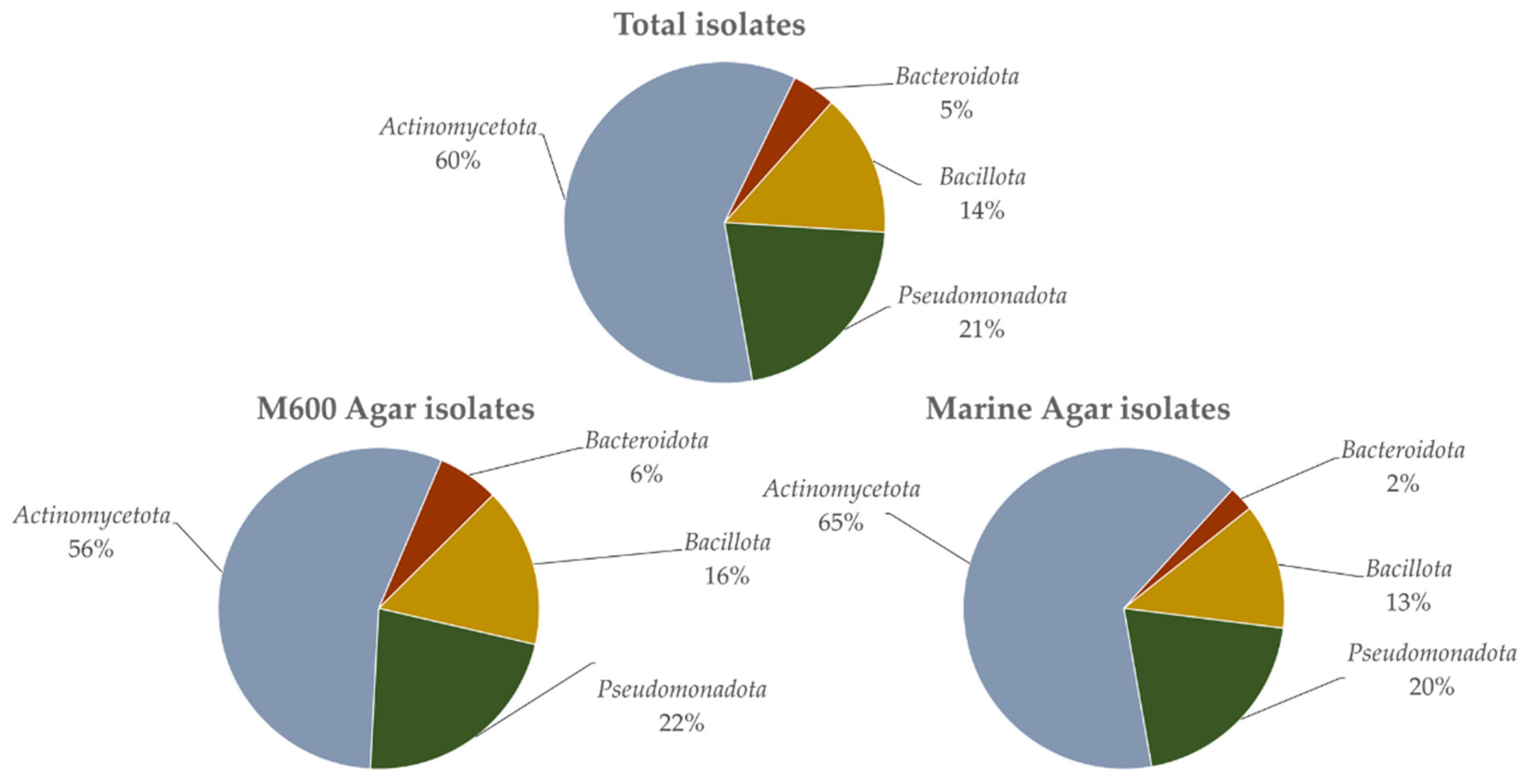
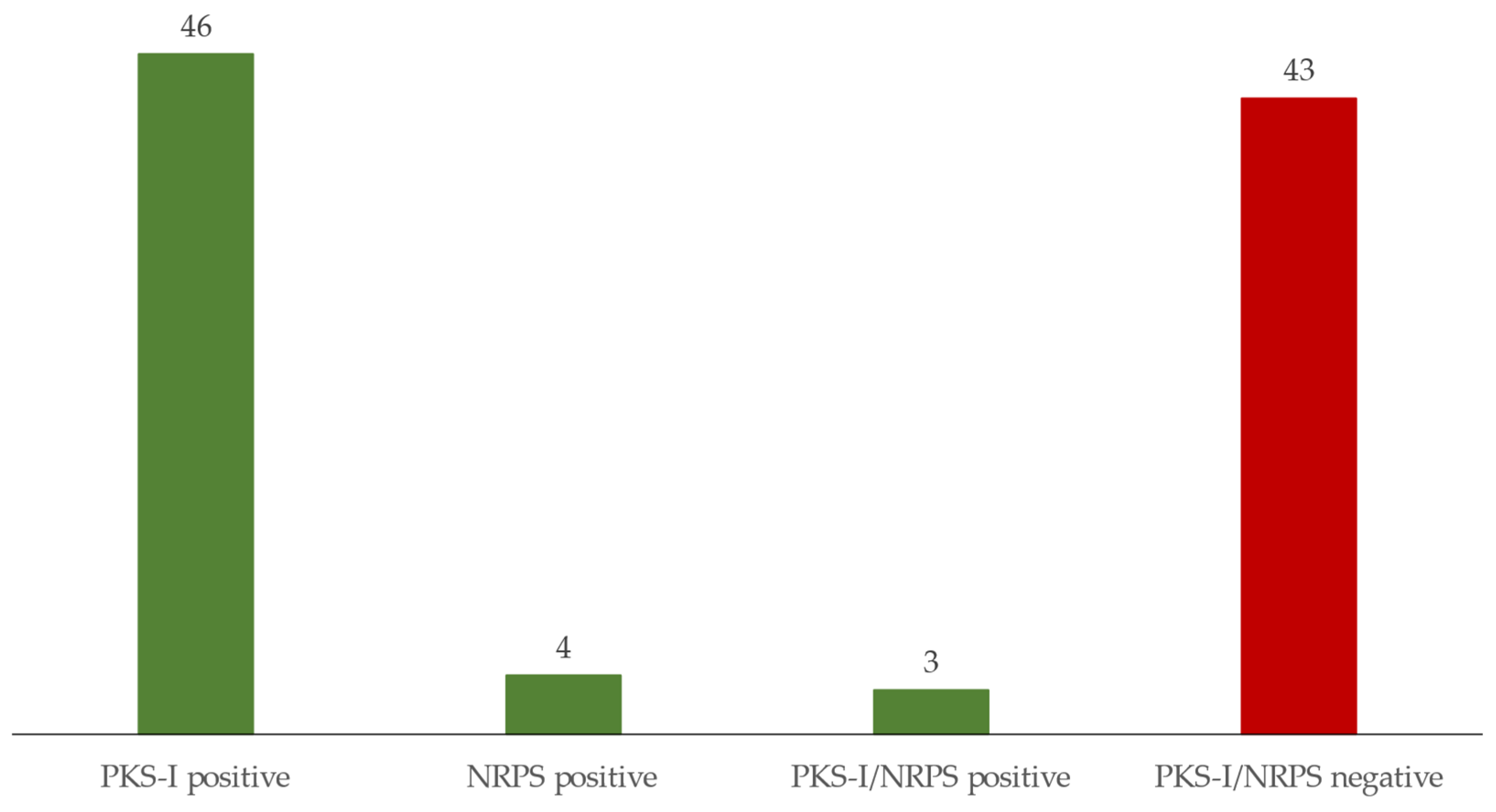
3.1. Isolation and Identification of Strains
3.2. Antimicrobial Screening
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NOAA (Ed.) Ocean Literacy: The Essential Principles and Fundamental Concepts of Ocean Sciences for Learners of All Ages; NOAA: Washington, DC, USA, 2020.
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [Green Version]
- NOAA. How Much of the Ocean Have We Explored? Available online: https://oceanservice.noaa.gov/facts/exploration.html (accessed on 7 March 2021).
- Hunter-Cevera, J.; Karl, D.; Buckley, M. Marine Microbial Diversity: The Key to Earth’s Habitability. In Proceedings of the American Academy of Microbiology, San Francisco, CA, USA, 8–10 April 2005. [Google Scholar]
- Haefner, B. Drugs from the deep: Marine natural products as drug candidates. Drug Discov. Today 2003, 8, 536–544. [Google Scholar] [CrossRef]
- Santos, J.D.; Vitorino, I.; Reyes, F.; Vicente, F.; Lage, O.M. From ocean to medicine: Pharmaceutical applications of metabolites from marine bacteria. Antibiotics 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Armelagos, G.J.; Kolbacher, K.; Collins, K.; Cook, J.; Krafeld-Daugherty, M. Tetracycline consumption in prehistory. In Tetracyclines in Biology, Chemistry and Medicine; Nelson, M., Hillen, W., Greenwald, R.A., Eds.; Birkhäuser Basel: Basel, Switzerland, 2001; pp. 219–236. [Google Scholar] [CrossRef]
- Montinari, M.R.; Minelli, S.; De Caterina, R. The first 3500 years of aspirin history from its roots—A concise summary. Vasc. Pharm. 2019, 113, 1–8. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Reynolds, K.A.; Luhavaya, H.; Li, J.; Dahesh, S.; Nizet, V.; Yamanaka, K.; Moore, B.S. Isolation and structure elucidation of lipopeptide antibiotic taromycin B from the activated taromycin biosynthetic gene cluster. J. Antibiot. 2018, 71, 333–338. [Google Scholar] [CrossRef]
- Anjum, K.; Sadiq, I.; Chen, L.; Kaleem, S.; Li, X.-C.; Zhang, Z.; Lian, X.-Y. Novel antifungal janthinopolyenemycins A and B from a co-culture of marine-associated Janthinobacterium spp. ZZ145 and ZZ148. Tetrahedron Lett. 2018, 59, 3490–3494. [Google Scholar] [CrossRef]
- Sudek, S.; Lopanik, N.B.; Waggoner, L.E.; Hildebrand, M.; Anderson, C.; Liu, H.; Patel, A.; Sherman, D.H.; Haygood, M.G. Identification of the putative bryostatin polyketide synthase gene cluster from “Candidatus Endobugula sertula”, the uncultivated microbial symbiont of the marine bryozoan. Bugula Neritina J. Nat. Prod. 2007, 70, 67–74. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine compounds and cancer: The first two decades of XXI century. Mar. Drugs 2019, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Whicher, J.R.; Hansen, D.A.; Hale, W.A.; Chemler, J.A.; Congdon, G.R.; Narayan, A.R.; Hakansson, K.; Sherman, D.H.; Smith, J.L.; et al. Structure of a modular polyketide synthase. Nature 2014, 510, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Strieker, M.; Tanovic, A.; Marahiel, M.A. Nonribosomal peptide synthetases: Structures and dynamics. Curr. Opin. Struct. Biol. 2010, 20, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Lacombe-Harvey, M.-È.; Brzezinski, R.; Beaulieu, C. Chitinolytic functions in actinobacteria: Ecology, enzymes, and evolution. Appl. Microbiol. Biotechnol. 2018, 102, 7219–7230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, P.R.; Lauro, F.M. An assessment of actinobacterial diversity in the marine environment. Antonie Van Leeuwenhoek 2008, 94, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Williams, S.T. Ecology of actinomycetes. Annu. Rev. Microbiol. 1983, 37, 189–216. [Google Scholar] [CrossRef]
- Maldonado, L.A.; Fenical, W.; Jensen, P.R.; Kauffman, C.A.; Mincer, T.J.; Ward, A.C.; Bull, A.T.; Goodfellow, M. Salinispora arenicola gen. nov., sp. nov. and Salinispora tropica sp. nov., obligate marine actinomycetes belonging to the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 2005, 55, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Marinomycins A-D, antitumor-antibiotics of a new structure class from a marine actinomycete of the recently discovered genus “Marinispora”. J. Am. Chem. Soc. 2006, 128, 1622–1632. [Google Scholar] [CrossRef]
- Azman, A.S.; Othman, I.; Velu, S.S.; Chan, K.G.; Lee, L.H. Mangrove rare actinobacteria: Taxonomy, natural compound, and discovery of bioactivity. Front. Microbiol. 2015, 6, 856. [Google Scholar] [CrossRef]
- Girão, M.; Ribeiro, I.; Ribeiro, T.; Azevedo, I.C.; Pereira, F.; Urbatzka, R.; Leão, P.N.; Carvalho, M.F. Actinobacteria isolated from Laminaria ochroleuca: A source of new bioactive compounds. Front. Microbiol. 2019, 10, 683. [Google Scholar] [CrossRef] [Green Version]
- Kamjam, M.; Sivalingam, P.; Deng, Z.; Hong, K. Deep sea actinomycetes and their secondary metabolites. Front. Microbiol. 2017, 8, 760. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Zhang, L.; Guo, X.; Dai, X.; Liu, L.; Xi, L.; Wang, J.; Song, L.; Wang, Y.; Zhu, Y.; et al. Diversity, biogeography, and biodegradation potential of Actinobacteria in the deep-sea sediments along the southwest Indian ridge. Front. Microbiol. 2016, 7, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stach, E.M.; Bull, A.T. Estimating and comparing the diversity of marine Actinobacteria. Antonie Van Leeuwenhoek 2005, 87, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Mincer, T.J.; Fenical, W.; Jensen, P.R. Culture-dependent and culture-independent diversity within the obligate marine actinomycete genus Salinispora. Appl. Environ. Microbiol. 2005, 71, 7019–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manivasagan, P.; Venkatesan, J.; Kim, S.-K. Introduction to marine Actinobacteria. In Marine Microbiology; Kim, S.-K., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2013; pp. 1–19. [Google Scholar] [CrossRef]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.M.; Pham, L.; Mehta, A.; Belanger, A.; Kanigan, T.; Lewis, K.; Epstein, S.S. Use of ichip for high-throughput in situ cultivation of “uncultivable” microbial species. Appl. Environ. Microbiol. 2010, 76, 2445–2450. [Google Scholar] [CrossRef] [Green Version]
- Berdy, B.; Spoering, A.L.; Ling, L.L.; Epstein, S.S. In situ cultivation of previously uncultivable microorganisms using the ichip. Nat. Protoc. 2017, 12, 2232–2242. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J. Planctomycetes diversity associated with macroalgae. FEMS Microbiol. Ecol. 2011, 78, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Bondoso, J.; Albuquerque, L.; Nobre, M.F.; Lobo-da-Cunha, A.; da Costa, M.S.; Lage, O.M. Aquisphaera giovannonii gen. nov., sp. nov., a planctomycete isolated from a freshwater aquarium. Int. J. Syst. Evol. Microbiol. 2011, 61, 2844–2850. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J.; Viana, F. Isolation and characterization of Planctomycetes from the sediments of a fish farm wastewater treatment tank. Arch. Microbiol. 2012, 194, 879–885. [Google Scholar] [CrossRef]
- Pavel, A.B.; Vasile, C.I. PyElph—A software tool for gel images analysis and phylogenetics. BMC Bioinform. 2012, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.K.; Garson, M.J.; Fuerst, J.A. Marine actinomycetes related to the ‘Salinospora’ group from the Great Barrier Reef sponge Pseudoceratina clavata. Environ. Microbiol. 2005, 7, 509–518. [Google Scholar] [CrossRef]
- Neilan, B.A.; Dittmann, E.; Rouhiainen, L.; Bass, R.A.; Schaub, V.; Sivonen, K.; Börner, T. Nonribosomal peptide synthesis and toxigenicity of Cyanobacteria. J. Bacteriol. 1999, 181, 4089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graça, A.P.; Calisto, R.; Lage, O.M. Planctomycetes as novel source of bioactive molecules. Front. Microbiol. 2016, 7, 1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, J.L.; Martín, J.; Tormo, J.R.; Vicente, F.; Brunati, M.; Ciciliato, I.; Losi, D.; Van Trappen, S.; Mergaert, J.; Swings, J.; et al. Bacterial diversity from benthic mats of Antarctic lakes as a source of new bioactive metabolites. Mar. Genom. 2009, 2, 33–41. [Google Scholar] [CrossRef]
- Santos, J.D.; Vitorino, I.; De la Cruz, M.; Díaz, C.; Cautain, B.; Annang, F.; Pérez-Moreno, G.; Gonzalez Martinez, I.; Tormo, J.R.; Martín, J.M.; et al. Bioactivities and extract dereplication of actinomycetales isolated from marine sponges. Front. Microbiol. 2019, 10, 727. [Google Scholar] [CrossRef]
- Perez-Victoria, I.; Martin, J.; Reyes, F. Combined LC/UV/MS and NMR strategies for the dereplication of marine natural products. Planta Med. 2016, 82, 857–871. [Google Scholar] [CrossRef] [Green Version]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glockner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzeby, J.; Amann, R.; Rossello-Mora, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Wang, D.S.; Xue, Q.H.; Ma, Y.Y.; Wei, X.L.; Chen, J.; He, F. Oligotrophy is helpful for the isolation of bioactive actinomycetes. Indian J. Microbiol. 2014, 54, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- Fdhila, F.; Vazquez, V.; Sanchez, J.L.; Riguera, R. dd-diketopiperazines: Antibiotics active against Vibrio anguillarum isolated from marine bacteria associated with cultures of Pecten maximus. J. Nat. Prod. 2003, 66, 1299–1301. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Puopolo, G.; Perazzolli, M.; Andolfi, A.; Melck, D.; Pertot, I.; Evidente, A. Cyclo(L-PRO-L-TYR), The fungicide isolated from Lysobacter capsici AZ78: A structure–activity relationship study. Chem. Heterocycl. Compd. 2014, 50, 290–295. [Google Scholar] [CrossRef]
- Plouffe, D.; Brinker, A.; McNamara, C.; Henson, K.; Kato, N.; Kuhen, K.; Nagle, A.; Adrian, F.; Matzen, J.T.; Anderson, P.; et al. In silico activity profiling reveals the mechanism of action of antimalarials discovered in a high-throughput screen. Proc. Natl. Acad Sci. USA 2008, 105, 9059–9064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, F.; Zahner, H.; Metzger, J.W.; Freund, S.; Hummel, R.P. Germicidin, an autoregulative germination inhibitor of Streptomyces viridochromogenes NRRL B-1551. J. Antibiot 1993, 46, 1126–1138. [Google Scholar] [CrossRef] [Green Version]
- Omura, S.; Nakagawa, A.; Imamura, N.; Kushida, K.; Liu, C.M.; Sello, L.H.; Westley, J.W. Structure of a new macrolide antibiotic, X-14952B. J. Antibiot 1985, 38, 674–676. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Chen, S.; Yan, Z.; Han, J.; Ta, Y.; Pu, T.; Wang, Y. Antimicrobial activity and identification of the biosynthetic gene cluster of X-14952B from Streptomyces sp. 135. Front. Microbiol. 2021, 12, 703093. [Google Scholar] [CrossRef]
- Scopel, M.; Abraham, W.R.; Henriques, A.T.; Macedo, A.J. Dipeptide cis-cyclo(Leucyl-Tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorg. Med. Chem. Lett. 2013, 23, 624–626. [Google Scholar] [CrossRef]
- Kumar, N.; Gorantla, J.N.; Mohandas, C.; Nambisan, B.; Lankalapalli, R.S. Isolation and antifungal properties of cyclo(D-Tyr-L-Leu) diketopiperazine isolated from Bacillus sp. associated with rhabditid entomopathogenic nematode. Nat. Prod. Res. 2013, 27, 2168–2172. [Google Scholar] [CrossRef]
- Zhao, T.; Chang, Y.; Zhu, T.; Li, J.; Gu, Q.; Li, D.; Che, Q.; Zhang, G. alpha-Pyrone derivatives with cyto-protective activity from two Takla Makan desert soil derived actinomycete Nocardiopsis strains recovered in seawater based medium. Nat. Prod. Res. 2019, 33, 2498–2506. [Google Scholar] [CrossRef]
- Xu, Z.; Ding, L.; Hertweck, C. A branched extender unit shared between two orthogonal polyketide pathways in an endophyte. Angew. Chem. Int. Ed. Engl. 2011, 50, 4667–4670. [Google Scholar] [CrossRef]
- Takada, K.; Ninomiya, A.; Naruse, M.; Sun, Y.; Miyazaki, M.; Nogi, Y.; Okada, S.; Matsunaga, S. Surugamides A-E, cyclic octapeptides with four D-amino acid residues, from a marine Streptomyces sp.: LC-MS-aided inspection of partial hydrolysates for the distinction of D- and L-amino acid residues in the sequence. J. Org. Chem. 2013, 78, 6746–6750. [Google Scholar] [CrossRef] [PubMed]
- Le, T.C.; Yang, I.; Yoon, Y.J.; Nam, S.J.; Fenical, W. Ansalactams B-D illustrate further biosynthetic plasticity within the ansamycin pathway. Org. Lett. 2016, 18, 2256–2259. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.C.; Nam, S.J.; Gulder, T.A.; Kauffman, C.A.; Jensen, P.R.; Fenical, W.; Moore, B.S. Structure and biosynthesis of the marine streptomycete ansamycin ansalactam A and its distinctive branched chain polyketide extender unit. J. Am. Chem. Soc. 2011, 133, 1971–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Chai, W.; Lian, X.Y.; Zhang, Z. Novel propanamide analogue and antiproliferative diketopiperazines from mangrove Streptomyces sp. Q24. Nat. Prod. Res. 2017, 31, 1390–1396. [Google Scholar] [CrossRef]
- Cho, K.W.; Lee, H.S.; Rho, J.R.; Kim, T.S.; Mo, S.J.; Shin, J. New lactone-containing metabolites from a marine-derived bacterium of the genus Streptomyces. J. Nat. Prod. 2001, 64, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Lacret, R.; Oves-Costales, D.; Gomez, C.; Diaz, C.; de la Cruz, M.; Perez-Victoria, I.; Vicente, F.; Genilloud, O.; Reyes, F. New ikarugamycin derivatives with antifungal and antibacterial properties from Streptomyces zhaozhouensis. Mar. Drugs 2014, 13, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Tanaka, T.; Fukuhara, K.; Miyairi, N.; Yonehara, H.; Umezawa, H. Blastmycin, a new antibiotic from Streptomyces sp. J. Antibiot 1957, 10, 39–45. [Google Scholar]
- Hosotani, N.; Kumagai, K.; Nakagawa, H.; Shimatani, T.; Saji, I. Antimycins A10 approximately A16, seven new antimycin antibiotics produced by Streptomyces spp. SPA-10191 and SPA-8893. J. Antibiot. 2005, 58, 460–467. [Google Scholar] [CrossRef]
- Ambrose, P.J. Clinical pharmacokinetics of chloramphenicol and chloramphenicol succinate. Clin. Pharm. 1984, 9, 222–238. [Google Scholar] [CrossRef]
- Nakano, H.; Tomita, F.; Yamaguchi, K.; Nagashima, M.; Suzuki, T. Corynecin (chloramphenicol analogs) fermentation studies: Selective production of Corynecin I by Corynebacterium hydrocarboclastus grown on acetate. Biotechnol. Bioeng. 1977, 19, 1009–1018. [Google Scholar] [CrossRef]
- Undabarrena, A.; Valencia, R.; Cumsille, A.; Zamora-Leiva, L.; Castro-Nallar, E.; Barona-Gomez, F.; Camara, B. Rhodococcus comparative genomics reveals a phylogenomic-dependent non-ribosomal peptide synthetase distribution: Insights into biosynthetic gene cluster connection to an orphan metabolite. Microb. Genom. 2021, 7, 000621. [Google Scholar] [CrossRef]
- Naganuma, S.; Sakai, K.; Hasumi, K.; Endo, A. Acaterin, a novel inhibitor of acyl-CoA: Cholesterol acyltransferase produced by Pseudomonas sp. A92. J. Antibiot 1992, 45, 1216–1221. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.; Carlsson, M.; Uvell, H.; Islam, K.; Edlund, K.; Cullman, I.; Altermark, B.; Mei, Y.F.; Elofsson, M.; Willassen, N.P.; et al. Isolation and characterization of anti-adenoviral secondary metabolites from marine actinobacteria. Mar. Drugs 2014, 12, 799–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Pfoh, R.; Rühl, S.; Qin, S.; Laatsch, H. T-Muurolol sesquiterpenes from the marine Streptomyces sp. M491 and revision of the configuration of previously reported amorphanes. J. Nat. Prod. 2009, 72, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ogura, H.; Akasaka, K.; Oikawa, T.; Matsuura, N.; Imada, C.; Yasuda, H.; Igarashi, Y. Nocapyrones: Alpha- and gamma-pyrones from a marine-derived Nocardiopsis sp. Mar. Drugs 2014, 12, 4110–4125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raju, R.; Piggott, A.M.; Quezada, M.; Capon, R.J. Nocardiopsins C and D and nocardiopyrone A: New polyketides from an Australian marine-derived Nocardiopsis sp. Tetrahedron 2013, 69, 692–698. [Google Scholar] [CrossRef]
- Tang, X.; Eitel, K.; Kaysser, L.; Kulik, A.; Grond, S.; Gust, B. A two-step sulfation in antibiotic biosynthesis requires a type III polyketide synthase. Nat. Chem. Biol. 2013, 9, 610–615. [Google Scholar] [CrossRef]
- Tian, H.; Shafi, J.; Ji, M.; Bi, Y.; Yu, Z. Antimicrobial metabolites from Streptomyces sp. SN0280. J. Nat. Prod. 2017, 80, 1015–1019. [Google Scholar] [CrossRef]
- Ding, L.; Maier, A.; Fiebig, H.-H.; Lin, W.-H.; Peschel, G.; Hertweck, C. Kandenols A–E, eudesmenes from an endophytic Streptomyces sp. of the mangrove tree Kandelia candel. J. Nat. Prod. 2012, 75, 2223–2227. [Google Scholar] [CrossRef]
- Kim, M.C.; Kwon, O.-W.; Park, J.-S.; Kim, S.Y.; Kwon, H.C. Nocapyrones H–J, 3,6-disubstituted α-pyrones from the marine actinomycete Nocardiopsis sp. KMF-001. Chem. Pharm. Bull. 2013, 61, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Tian, X.; Huang, C.; Li, Q.; Zhang, S. Marinactinones A–C, new γ-pyrones from marine actinomycete Marinactinospora thermotolerans SCSIO 00606. J. Antibiot. 2011, 64, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brachmann, A.O.; Brameyer, S.; Kresovic, D.; Hitkova, I.; Kopp, Y.; Manske, C.; Schubert, K.; Bode, H.B.; Heermann, R. Pyrones as bacterial signaling molecules. Nat. Chem. Biol. 2013, 9, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Torres, J.P.; Ammon, M.A.; Marett, L.; Teichert, R.W.; Reilly, C.A.; Kwan, J.C.; Hughen, R.W.; Flores, M.; Tianero, M.D.; et al. A bacterial source for mollusk pyrone polyketides. Chem. Biol. 2013, 20, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneemann, I.; Ohlendorf, B.; Zinecker, H.; Nagel, K.; Wiese, J.; Imhoff, J.F. Nocapyrones A−D, γ-pyrones from a Nocardiopsis strain isolated from the marine sponge Halichondria panicea. J. Nat. Prod. 2010, 73, 1444–1447. [Google Scholar] [CrossRef] [Green Version]
- Neel, J.K. A Limnological investigation of the psammon in douglas lake, michigan, with especial reference to shoal and shoreline dynamics. Trans. Am. Microsc. Soc. 1948, 67, 1–53. [Google Scholar] [CrossRef]
- Whitman, R.; Harwood, V.J.; Edge, T.A.; Nevers, M.; Byappanahalli, M.; Vijayavel, K.; Brandao, J.; Sadowsky, M.J.; Alm, E.W.; Crowe, A.; et al. Microbes in beach sands: Integrating environment, ecology and public health. Rev. Environ. Sci. Biotechnol. 2014, 13, 329–368. [Google Scholar] [CrossRef] [Green Version]
- Probandt, D.; Eickhorst, T.; Ellrott, A.; Amann, R.; Knittel, K. Microbial life on a sand grain: From bulk sediment to single grains. ISME J. 2018, 12, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Gobet, A.; Böer, S.I.; Huse, S.M.; van Beusekom, J.E.E.; Quince, C.; Sogin, M.L.; Boetius, A.; Ramette, A. Diversity and dynamics of rare and of resident bacterial populations in coastal sands. ISME J. 2012, 6, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Musat, N.; Werner, U.; Knittel, K.; Kolb, S.; Dodenhof, T.; van Beusekom, J.E.; de Beer, D.; Dubilier, N.; Amann, R. Microbial community structure of sandy intertidal sediments in the North Sea, Sylt-Romo Basin, Wadden Sea. Syst. Appl. Microbiol. 2006, 29, 333–348. [Google Scholar] [CrossRef]
- Huang, Z.; Mo, S.; Yan, L.; Wei, X.; Huang, Y.; Zhang, L.; Zhang, S.; Liu, J.; Xiao, Q.; Lin, H.; et al. A simple culture method enhances the recovery of culturable actinobacteria from coastal sediments. Front. Microbiol. 2021, 12, 1451. [Google Scholar] [CrossRef]
- Cohen-Bazire, G.; Sistrom, W.R.; Stanier, R.Y. Kinetic studies of pigment synthesis by non-sulfur purple bacteria. J. Cell. Comp. Physiol. 1957, 49, 25–68. [Google Scholar] [CrossRef]
- Schlesner, H. Pirella marina sp. nov., a budding, peptidoglycan-less bacterium from brackish water. Syst. Appl. Microbiol. 1986, 8, 177–180. [Google Scholar] [CrossRef]
- Field, E.K.; Kato, S.; Findlay, A.J.; MacDonald, D.J.; Chiu, B.K.; Luther III, G.W.; Chan, C.S. Planktonic marine iron oxidizers drive iron mineralization under low-oxygen conditions. Geobiology 2016, 14, 499–508. [Google Scholar] [CrossRef]
- Ribeiro, I.; Girão, M.; Alexandrino, D.A.M.; Ribeiro, T.; Santos, C.; Pereira, F.; Mucha, A.P.; Urbatzka, R.; Leão, P.N.; Carvalho, M.F. Diversity and bioactive potential of actinobacteria isolated from a coastal marine sediment in northern Portugal. Microorganisms 2020, 8, 1691. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Jha, B. Intertidal marine sediment harbours Actinobacteria with promising bioactive and biosynthetic potential. Sci. Rep. 2017, 7, 10041. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Davo, A.; Dias, T.; Gomes, S.E.; Rodrigues, S.; Parera-Valadez, Y.; Borralho, P.M.; Pereira, F.; Rodrigues, C.M.; Santos-Sanches, I.; Gaudencio, S.P. The Madeira archipelago as a significant source of marine-derived actinomycete diversity with anticancer and antimicrobial potential. Front. Microbiol. 2016, 7, 1594. [Google Scholar] [CrossRef]
- Jensen, P.R.; Mafnas, C. Biogeography of the marine actinomycete Salinispora. Environ. Microbiol. 2006, 8, 1881–1888. [Google Scholar] [CrossRef]
- Jensen, P.R.; Gontang, E.; Mafnas, C.; Mincer, T.J.; Fenical, W. Culturable marine actinomycete diversity from tropical Pacific Ocean sediments. Environ. Microbiol. 2005, 7, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.R.; Mincer, T.J.; Williams, P.G.; Fenical, W. Marine actinomycete diversity and natural product discovery. Antonie Van Leeuwenhoek 2005, 87, 43–48. [Google Scholar] [CrossRef]
- Tamura, S.; Suzuki, A.; Aoki, Y.; Otake, N. Isolation of Several Diketopiperazines from Peptone. Agric. Biol. Chem. 1964, 28, 650–652. [Google Scholar] [CrossRef]
- Khare, E.; Tyagi, S.; Patil, K.S. Chapter 5—Language of plant-microbe-microbe interactions in rhizospheric ecosystems. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 59–76. [Google Scholar] [CrossRef]
- Jones, D.; Keddie, R.M. The Genus Arthrobacter. In The Prokaryotes: Volume 3: Archaea. Bacteria: Firmicutes, Actinomycetes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 945–960. [Google Scholar] [CrossRef]
- Dhakal, D.; Rayamajhi, V.; Mishra, R.; Sohng, J.K. Bioactive molecules from Nocardia: Diversity, bioactivities and biosynthesis. J. Ind. Microbiol. Biotechnol. 2019, 46, 385–407. [Google Scholar] [CrossRef]
- Bennur, T.; Ravi Kumar, A.; Zinjarde, S.S.; Javdekar, V. Nocardiopsis species: A potential source of bioactive compounds. J. Appl. Microbiol. 2016, 120, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.H.; Desoukey, S.Y.; Fouad, M.A.; Kamel, M.S.; Gulder, T.A.M.; Abdelmohsen, U.R. Natural product potential of the genus Nocardiopsis. Mar. Drugs 2018, 16, 147. [Google Scholar] [CrossRef] [PubMed]
- Tsujibo, H.; Sato, T.; Inui, M.; Yamamoto, H.; Inamori, Y. Intracellular accumulation of phenazine antibiotics produced by an alkalophilic actinomycete. I. Taxonomy, isolation and identification of the phenazine antibiotics. Agric. Biol. Chem. 1988, 52, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, K.; Degnes, K.F.; Kemmler, M.; Bredholt, H.; Fjaervik, E.; Klinkenberg, G.; Sletta, H.; Ellingsen, T.E.; Zotchev, S.B. Production of a new thiopeptide antibiotic, TP-1161, by a marine Nocardiopsis species. Appl. Environ. Microbiol. 2010, 76, 4969–4976. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Chen, X.; Li, W.; Lu, C.; Shen, Y. New diketopiperazine derivatives with cytotoxicity from Nocardiopsis sp. YIM M13066. J. Antibiot. 2017, 70, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wan, C.; Zhang, L. A new diketopiperazine isolated from a Nocardiopsis strain TRM20105 guided by bioassay against Candida albicans. Nat. Prod. Res. 2019, 33, 3421–3425. [Google Scholar] [CrossRef]
- Li, Y.; Lai, Y.-M.; Lu, Y.; Yang, Y.-L.; Chen, S. Analysis of the biosynthesis of antibacterial cyclic dipeptides in Nocardiopsis alba. Arch. Microbiol. 2014, 196, 765–774. [Google Scholar] [CrossRef]
- Sunish, K.S.; Sreedharan, P.; Daniel, S.; Biji, M.; Rosamma, P.; Sukumaran, V.; Mohandas, A.; Singh, I.S.B. A novel substituted derivative of sterol from marine actinomycetes Nocardiopsis alba MCCB 110 antagonistic to the aquaculture pathogen Vibrio harveyi. Microb. Pathog. 2021, 157, 104967. [Google Scholar] [CrossRef]
- Iwatsuki, M.; Tomoda, H.; Uchida, R.; Gouda, H.; Hirono, S.; Ōmura, S. Lariatins, antimycobacterial peptides produced by Rhodococcus sp. K01−B0171, have a lasso structure. J. Am. Chem. Soc. 2006, 128, 7486–7491. [Google Scholar] [CrossRef]
- Kurosawa, K.; Ghiviriga, I.; Sambandan, T.G.; Lessard, P.A.; Barbara, J.E.; Rha, C.; Sinskey, A.J. Rhodostreptomycins, antibiotics biosynthesized following horizontal gene transfer from Streptomyces padanus to Rhodococcus fascians. J. Am. Chem. Soc. 2008, 130, 1126–1127. [Google Scholar] [CrossRef]
- Moumbock, A.F.A.; Gao, M.; Qaseem, A.; Li, J.; Kirchner, P.A.; Ndingkokhar, B.; Bekono, B.D.; Simoben, C.V.; Babiaka, S.B.; Malange, Y.I.; et al. StreptomeDB 3.0: An updated compendium of streptomycetes natural products. Nucleic Acids Res. 2021, 49, D600–D604. [Google Scholar] [CrossRef]
- Yan, L.-L.; Han, N.-N.; Zhang, Y.-Q.; Yu, L.-Y.; Chen, J.; Wei, Y.-Z.; Li, Q.-P.; Tao, L.; Zheng, G.-H.; Yang, S.-E.; et al. Antimycin A18 produced by an endophytic Streptomyces albidoflavus isolated from a mangrove plant. J. Antibiot. 2010, 63, 259–261. [Google Scholar] [CrossRef]
- Wang, L.; Yun, B.-S.; George, N.P.; Wendt-Pienkowski, E.; Galm, U.; Oh, T.-J.; Coughlin, J.M.; Zhang, G.; Tao, M.; Shen, B. Glycopeptide antitumor antibiotic zorbamycin from Streptomyces flavoviridis ATCC 21892: Strain improvement and structure elucidation. J. Nat. Prod. 2007, 70, 402–406. [Google Scholar] [CrossRef] [Green Version]
- Reusser, F. Mode of Action of Zorbamycin. J. Bacteriol. 1971, 108, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Ochi, K.; Tsurumi, Y.; Shigematsu, N.; Iwami, M.; Umehara, K.; Okuhara, M. Physiological analysis of bicozamycin high-producing Streptomyces griseoflavus used at industrial level. J. Antibiot 1988, 41, 1106–1115. [Google Scholar] [CrossRef]
- Grote, R.; Zeeck, A.; Drautz, H.; Zahner, H. Metabolic products of microorganisms. 244. Colabomycins, new antibiotics of the manumycin group from Streptomyces griseoflavus. I. Isolation, characterization and biological properties. J. Antibiot. 1988, 41, 1178–1185. [Google Scholar] [CrossRef] [Green Version]
- Palla, M.S.; Guntuku, G.S.; Muthyala, M.K.K.; Pingali, S.; Sahu, P.K. Isolation and molecular characterization of antifungal metabolite producing actinomycete from mangrove soil. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 250–256. [Google Scholar] [CrossRef]
- Lewis, R.A.; Devi, J.; Green, K.; Li, J.; Hopkins, A.; Hayles, J.; Nurse, P.; Errington, J.; Allenby, N.E.E. Screening and purification of natural products from actinomycetes that induce a “rounded” morphological phenotype in fission yeast. Nat. Prod. Bioprospect. 2021, 11, 431–445. [Google Scholar] [CrossRef]
- Kawabata, K.; Inamoto, Y.; Sakane, K.; Iwamoto, T.; Hashimoto, S. Synthesis and structure determination of FR109615, a new antifungal antibiotic. J. Antibiot 1990, 43, 513–518. [Google Scholar] [CrossRef]
- Iwamoto, T.; Tsujii, E.; Ezaki, M.; Fujie, A.; Hashimoto, S.; Okuhara, M.; Kohsaka, M.; Imanaka, H.; Kawabata, K.; Inamoto, Y.; et al. FR109615, a new antifungal antibiotic from Streptomyces setonii. Taxonomy, fermentation, isolation, physico-chemical properties and biological activity. J. Antibiot. 1990, 43, 1–7. [Google Scholar] [CrossRef]
- Neumann, C.S.; Fujimori, D.G.; Walsh, C.T. Halogenation strategies in natural product biosynthesis. Chem. Biol. 2008, 15, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Hai, Y.; Wei, M.-Y.; Wang, C.-Y.; Gu, Y.-C.; Shao, C.-L. The intriguing chemistry and biology of sulfur-containing natural products from marine microorganisms (1987–2020). Mar. Life Sci. Technol. 2021, 3, 488–518. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain ID | Affiliation | NRPS | PKS-I | % Inhibition | ||
|---|---|---|---|---|---|---|
| % | Taxonomic Unit | E. coli | S. aureus | |||
| PMIC_1E12B | 100.00 | Arthrobacter gandavensis R 5812 | N.A | A | 64.8 | 39.6 |
| PMIC_1F10C.1 | 100.00 | Arthrobacter gandavensis R 5812 | N.A | A | 1.7 | 2.9 |
| PMIC_1H7A | 100.00 | Kocuria polaris CMS 76or | A | N.A | 1.9 | −16.3 |
| PMIC_1A10B | 99.82 | Nocardia nova NBRC 15556 | N.A | A | 50.6 | −12.7 |
| PMIC_1A11B.2 | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 76.3 | 17.6 |
| PMIC_1F6A.3 | 99.76 | Nocardiopsis alba DSM 43377 | N.A | A | 79.2 | 22.4 |
| PMIC_2A11A.1 | 99.92 | Nocardiopsis alba DSM 43377 | N.A | A | 39.6 | 19.2 |
| PMIC_2A11B.1 | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 77.7 | −8.3 |
| PMIC_2A11B.2 | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 12.9 | 4.2 |
| PMIC_2A11B.3 | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 37.4 | 22.6 |
| PMIC_2F6A | 99.76 | Nocardiopsis alba DSM 43377 | N.A | A | 45.2 | 30.8 |
| PMIC_2F6B | 99.76 | Nocardiopsis alba DSM 43377 | N.A | A | 15.7 | 1.0 |
| PMIC_2F6C | 99.77 | Nocardiopsis alba DSM 43377 | N.A | A | 19.4 | 18.6 |
| PMIC_2H2A | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 26.7 | 26.8 |
| PMIC_2H2C.1 | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 38.7 | 48.5 |
| PMIC_2H2C.2 | 100.00 | Nocardiopsis alba DSM 43377 | N.A | A | 33.4 | 52.0 |
| PMIC_2B1A | 99.85 | Nocardiopsis prasina DSM 43845 | N.A | A | 14.1 | 3.4 |
| PMIC_2B1C | 99.82 | Nocardiopsis prasina DSM 43845 | N.A | A | 12.8 | 17.0 |
| PMIC_2B1D | 99.85 | Nocardiopsis prasina DSM 43845 | N.A | A | 38.4 | 49.4 |
| PMIC_2D10A | 99.84 | Nocardiopsis prasina DSM 43845 | N.A | A | 20.0 | 14.9 |
| PMIC_2D10B.1 | 99.75 | Nocardiopsis prasina DSM 43845 | N.A | A | 35.6 | 22.2 |
| PMIC_2D10B.2 | 99.84 | Nocardiopsis prasina DSM 43845 | N.A | A | 19.8 | 7.5 |
| PMIC_2D10C | 99.84 | Nocardiopsis prasina DSM 43845 | N.A | A | 44.1 | 36.1 |
| PMIC_2F12A | 99.82 | Nocardiopsis prasina DSM 43845 | N.A | A | −5.4 | 17.3 |
| PMIC_2C3B.2 | 99.11 | Nocardiopsis umidischolae 66/93 | N.A | A | 41.4 | 7.9 |
| PMIC_2C3B.3 | 99.19 | Nocardiopsis umidischolae 66/93 | N.A | A | 39.1 | 23.1 |
| PMIC_2C3B.4 | 99.20 | Nocardiopsis umidischolae 66/93 | N.A | A | 16.9 | 13.5 |
| PMIC_2H6 | 99.81 | Plantibacter flavus VKM Ac-2504 | N.A | A | −3.4 | −9.8 |
| PMIC_1E10C | 99.69 | Rhodococcus coprophilus NBRC 100603 | N.A | A | 86.7 | 16.6 |
| PMIC_2E10 | 99.69 | Rhodococcus coprophilus NBRC 100603 | A | A | 24 | −9.5 |
| PMIC_1E9B | 100.00 | Rhodococcus erythropolis NBRC 15567 | A | A | 8.9 | −0.9 |
| PMIC_2E9C | 99.76 | Rhodococcus qingshengii JCM 15477 | A | N.A | 3.2 | −11.5 |
| PMIC_1C12A | 99.67 | Streptomyces albidoflavus DSM 40455 | A | N.A | 1.9 | −18.2 |
| PMIC_2C12 | 99.69 | Streptomyces albidoflavus DSM 40455 | N.A | A | 64.8 | 13.1 |
| PMIC_1A8C | 99.66 | Streptomyces albogriseolus NRRL B-1305 | N.A | A | 2.0 | 39.9 |
| PMIC_2G8C | 99.68 | Streptomyces ambofaciens ATCC 23877 | N.A | A | 30.9 | 12.0 |
| PMIC_1C8A | 99.69 | Streptomyces ardesiacus NRRL B-1773 | N.A | A | 34.0 | 12.3 |
| PMIC_2C8A | 99.69 | Streptomyces ardesiacus NRRL B-1773 | N.A | A | 33.1 | 31.9 |
| PMIC_2C8B | 99.70 | Streptomyces ardesiacus NRRL B-1773 | A | A | 10.7 | −2.0 |
| PMIC_2D8A | 100.00 | Streptomyces ardesiacus NRRL B-1773 | N.A | A | 20.8 | 10.4 |
| PMIC_2D8B | 100.00 | Streptomyces ardesiacus NRRL B-1773 | N.A | A | 27.6 | 28.4 |
| PMIC_1A8B | 99.43 | Streptomyces flavoviridis NBRC 12772 | N.A | A | 75.9 | 100 |
| PMIC_1D9A | 99.80 | Streptomyces griseoflavus LMG 19344 | N.A | A | 30.1 | 4.8 |
| PMIC_1D9B | 99.75 | Streptomyces griseoflavus LMG 19344 | N.A | A | 64.6 | 9.8 |
| PMIC_1I1A | 100.00 | Streptomyces hydrogenans NBRC 13475 | N.A | A | 80.2 | 11.9 |
| PMIC_1I1B | 100.00 | Streptomyces hydrogenans NBRC 13475 | N.A | A | 30.1 | 26.6 |
| PMIC_2C8C | 99.57 | Streptomyces hydrogenans NBRC 13475 | N.A | A | 36.5 | 15.6 |
| PMIC_2D11A.2 | 99.69 | Streptomyces hydrogenans NBRC 13475 | N.A | A | 46.1 | 31.7 |
| PMIC_2D11C | 99.70 | Streptomyces hydrogenans NBRC 13475 | N.A | A | 21.4 | 26.1 |
| PMIC_1F12B | 100.00 | Streptomyces setonii NRRL ISP-5322 | N.A | A | 62.1 | 21.6 |
| PMIC_2F12B | 100.00 | Streptomyces setonii NRRL ISP-5322 | N.A | A | 28.5 | 40.8 |
| PMIC_1B3A.1 | 99.52 | Streptomyces xiamenensis MCCC 1A01550 | N.A | A | 29.4 | 15.5 |
| PMIC_2C2B | 99.81 | Streptomyces xiamenensis MCCC 1A01550 | A | N.A | 35.8 | 18.8 |
| Culture medium extract | 25.8 | 9.9 | ||||
| Strain ID | Affiliation | Inhibition (%) | Inhibition (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E. coli | S. aureus | |||||||||||||
| % | Taxonomic Unit | M607 | OSMAC 1:10 M607 | OSMAC M607 | OSMAC M600 | OSMAC MA | OSMAC CGY | M607 | OSMAC 1:10 M607 | OSMAC M607 | OSMAC M600 | OSMAC MA | OSMAC CGY | |
| PMIC_1E12B | 100% | Arthrobacter gandavensis R 5812 | 64.8 | 24.6 | 17.4 | 16.3 | 29.8 | 29.9 | 39.6 | −6.5 | −9.8 | 3.7 | 2.8 | −10.4 |
| PMIC_1A10B | 99.82% | Nocardia nova NBRC 15556 | 50.6 | 100.0 | 62.0 | 79.9 | 54.3 | 87.0 | −12.7 | 28.8 | 27.0 | 22.6 | −25.4 | 6.2 |
| PMIC_1A11B.2 | 100% | Nocardiopsis alba DSM 43377 | 76.3 | 23.6 | 17.4 | 16.3 | 29.8 | 29.9 | 17.6 | −10.1 | −9.8 | 3.7 | 2.8 | −10.4 |
| PMIC_2A11B.1 | 100% | Nocardiopsis alba DSM 43377 | 77.7 | 40.9 | 43.1 | 45.7 | 41.7 | 45.0 | −8.3 | 19.2 | 29.0 | 19.0 | 16.7 | 36.5 |
| PMIC_1F6A.3 | 99.76% | Nocardiopsis alba DSM 43377 | 79.2 | 14.7 | 21.4 | 15.4 | 26.1 | 15.9 | 22.4 | −12.2 | −13.5 | −16.8 | 9.5 | −3.2 |
| PMIC_2H2C.2 | 100% | Nocardiopsis alba DSM 43377 | 33.4 | 91.7 | 42.9 | 54.7 | 44.1 | 68.1 | 52.0 | 26.5 | 30.9 | 40.3 | 31.7 | 41.1 |
| PMIC_1E10C | 99.69% | Rhodococcus coprophilus NBRC 100603 | 86.7 | 31.2 | 44.9 | 48.0 | 42.7 | 33.1 | 16.6 | 23.9 | 24.8 | 7.6 | 19.3 | 21.0 |
| PMIC_2C12 | 99.69% | Streptomyces albidoflavus DSM 40455 | 64.8 | 22.0 | 35.4 | 31.0 | 32.2 | 0.0 | 13.1 | 19.8 | −14.6 | −9.2 | −3.7 | −15.3 |
| PMIC_1A8B | 99.43% | Streptomyces flavoviridis NBRC 12772 | 75.9 | 39.1 | 41.8 | 40.5 | 42.4 | 46.7 | 100.0 | 100.0 | 77.2 | 100.0 | 37.0 | 40.3 |
| PMIC_1D9B | 99.75% | Streptomyces griseoflavus LMG 19344 | 64.6 | 57.2 | 54.3 | 54.2 | 43.2 | 98.3 | 9.8 | 27.8 | 34.3 | 27.3 | 5.0 | 91.1 |
| PMIC_1I1A | 100% | Streptomyces hydrogenans NBRC 13475 | 80.2 | 53.7 | 44.0 | 49.7 | 44.9 | 52.0 | 11.9 | 73.2 | 14.8 | 14.3 | 19.1 | 17.0 |
| PMIC_1F12B | 100% | Streptomyces setonii NRRL ISP-5322 | 62.1 | 93.1 | 70.4 | 52.9 | 48.7 | 42.1 | 21.6 | 31.5 | 33.5 | 21.9 | 23.3 | −15.0 |
| Culture medium extract | 25.8 | 9.0 | 11.7 | 15.4 | 9.3 | −0.2 | 9.9 | 14.7 | 12.0 | 10.2 | 5.5 | 5.9 | ||
| Strain ID | Taxonomic Unit | Medium | Putatively Detected Bioactive Molecules | Non-Identified Molecules |
|---|---|---|---|---|
| PMIC_1E12B | Arthrobacter gandavensis | M607 | cyclo(Pro-Tyr), cyclo(L-Leu-L-Pro) | − |
| OSMAC 1:10 M607 | − | − | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | − | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | − | − | ||
| PMIC_1A10B | Nocardia nova | M607 | N-acetyltyramine, cyclo(L-Leu-L-Pro) | − |
| OSMAC 1:10 M607 | N-acetyltyramine, cyclo(L-Leu-L-Pro) | C9H10N2O; C15H24O3 | ||
| OSMAC M607 | cyclo(Pro-Tyr), cyclo(L-Leu-L-Pro) | C27H54N10O10 | ||
| OSMAC M600 | cyclo(pro-tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro) | C22H44O12 | ||
| OSMAC MA | cyclo(pro-tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro) | − | ||
| OSMAC CGY | N-acetyltyramine, cyclo(L-Leu-L-Pro) | C22H44O12; C24H48O13 | ||
| PMIC_1A11B.2 | Nocardiopsis alba | M607 | cyclo(pro-tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), germicidin A | C12H25NO3, C14H29NO3 |
| OSMAC 1:10 M607 | − | − | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | − | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | − | − | ||
| PMIC_2A11B.1 | Nocardiopsis alba | M607 | cyclo(Pro-Trp), N-acetyltyramine, cyclo(L-Leu-L-Pro), germicidin A | − |
| OSMAC 1:10 M607 | − | − | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | N-acetyltyramine, cyclo(L-Leu-L-Pro), cyclo(Pro-Trp), germicidin A | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | N-acetyltyramine, cyclo(L-Leu-L-Pro), germicidin A, antibiotic X-14952B | − | ||
| PMIC_1F6A.3 | Nocardiopsis alba | M607 | cyclo(Pro-Tyr), cyclo(Tyr-Leu), cyclo(L-Leu-L-Pro) | C20H31NO4S |
| OSMAC 1:10 M607 | − | − | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | − | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | − | − | ||
| PMIC_2H2C.2 | Nocardiopsis alba | M607 | cyclo(L-Leu-L-Pro) | − |
| OSMAC 1:10 M607 | cyclo(L-Leu-L-Pro) | C27H52N10O10 | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | N-acetyltyramine, cyclo(L-Leu-L-Pro), antibiotic X-14952B | C15H24O4 | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | cyclo(Pro-Tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), antibiotic X-14952B | C15H24O4, C16H24O3 | ||
| PMIC_1E10C | Rhodococcus coprophilus | M607 | cyclo(L-Leu-L-Pro) | C14H29NO3 |
| OSMAC 1:10 M607 | − | − | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | − | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | − | − | ||
| PMIC_2C12 | Streptomyces albidoflavus | M607 | cyclo(Pro-Tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), germicidin G, surugamide A, ansalactam A | − |
| OSMAC 1:10 M607 | − | − | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | − | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | − | − | ||
| PMIC_1A8B | Streptomyces flavoviridis | M607 | N-acetyltyramine, cyclo(L-Leu-L-Pro), | C20H13N3O6, C23H13ClO4S2 |
| OSMAC 1:10 M607 | cyclo(L-Leu-L-Pro) | C23H13ClO4S2 | ||
| OSMAC M607 | cyclo(pro-tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro) | − | ||
| OSMAC M600 | N-acetyltyramine, cyclo(L-Leu-L-Pro) | C20H13N3O6, C23H13ClO4S2 | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | − | − | ||
| PMIC_1D9B | Streptomyces griseoflavus | M607 | N-acetyltyramine, cyclo(L-Leu-L-Pro), ansalactam A | − |
| OSMAC 1:10 M607 | N-acetyltyramine, ansalactam A | − | ||
| OSMAC M607 | cyclo(Pro-Tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), 3-acetylamino-N-2-thienylpropanamide, ansalactam A | − | ||
| OSMAC M600 | cyclo(Pro-Tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), 3-acetylamino-N-2-thienylpropanamide, ansalactam A | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | N-acetyltyramine, 3-acetylamino-N-2-thienylpropanamide, ansalactam A | − | ||
| PMIC_1I1A | Streptomyces hydrogenans | M607 | cyclo(Pro-Tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), antibiotic MKN-003B, germicidin G, surugamide A | C13H22O3 |
| OSMAC 1:10 M607 | cyclo(L-Leu-L-Pro), antibiotic MKN-003B, germicidin G, surugamide E, surugamide A | C13H22O3 | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | − | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | cyclo(Pro-Tyr), N-acetyltyramine, cyclo(L-Leu-L-Pro), antibiotic MKN-003B, germicidin G, surugamide E, surugamide A, blastmycin, antimycin A13, antimycin A11 | C13H22O3 | ||
| PMIC_1F12B | Streptomyces setonii | M607 | N-acetyltyramine, corynecin I, chloramphenicol | − |
| OSMAC 1:10 M607 | N-acetyltyramine, cyclo(L-Leu-L-Pro) | C27H53N5O10 | ||
| OSMAC M607 | − | − | ||
| OSMAC M600 | N-acetyltyramine, cyclo(L-Leu-L-Pro) | − | ||
| OSMAC MA | − | − | ||
| OSMAC CGY | N-acetyltyramine, cyclo(L-Leu-L-Pro) | C22H44O12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, J.D.N.; João, S.A.; Martín, J.; Vicente, F.; Reyes, F.; Lage, O.M. iChip-Inspired Isolation, Bioactivities and Dereplication of Actinomycetota from Portuguese Beach Sediments. Microorganisms 2022, 10, 1471. https://doi.org/10.3390/microorganisms10071471
dos Santos JDN, João SA, Martín J, Vicente F, Reyes F, Lage OM. iChip-Inspired Isolation, Bioactivities and Dereplication of Actinomycetota from Portuguese Beach Sediments. Microorganisms. 2022; 10(7):1471. https://doi.org/10.3390/microorganisms10071471
Chicago/Turabian Styledos Santos, José Diogo Neves, Susana Afonso João, Jesús Martín, Francisca Vicente, Fernando Reyes, and Olga Maria Lage. 2022. "iChip-Inspired Isolation, Bioactivities and Dereplication of Actinomycetota from Portuguese Beach Sediments" Microorganisms 10, no. 7: 1471. https://doi.org/10.3390/microorganisms10071471