
Lactobacillus salivarius SNK-6 Activates Intestinal Mucosal Immune System by Regulating Cecal Microbial Community Structure in Laying Hens

Abstract
:1. Introduction
2. Materials and Methods
2.1. Feed Preparation
2.2. Birds’ Diets and Management
2.3. Laying Performance Parameters
2.4. Quantitative Real-Time PCR for Measuring Immune-Related Gene Transcript Level in the Cecal Tonsil
2.5. DNA Extraction, Amplification, and Sequencing
2.6. Bioinformatic Analyses
2.7. Statistics Analysis
3. Results
3.1. Laying Performance
3.2. Cecal Tonsil Cytokines mRNA Levels
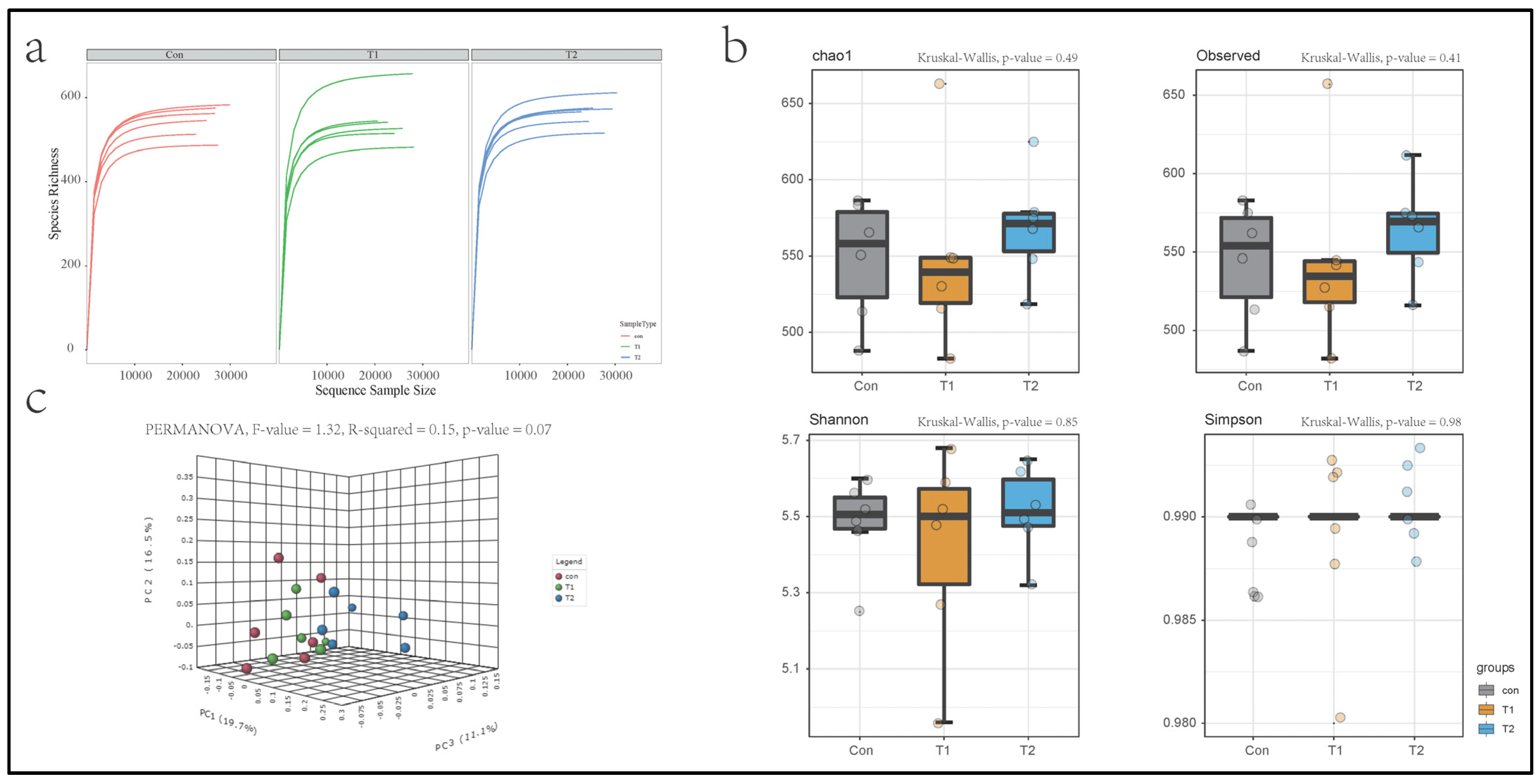
3.3. Intestinal Bacterial Richness, Diversity, and Similarity
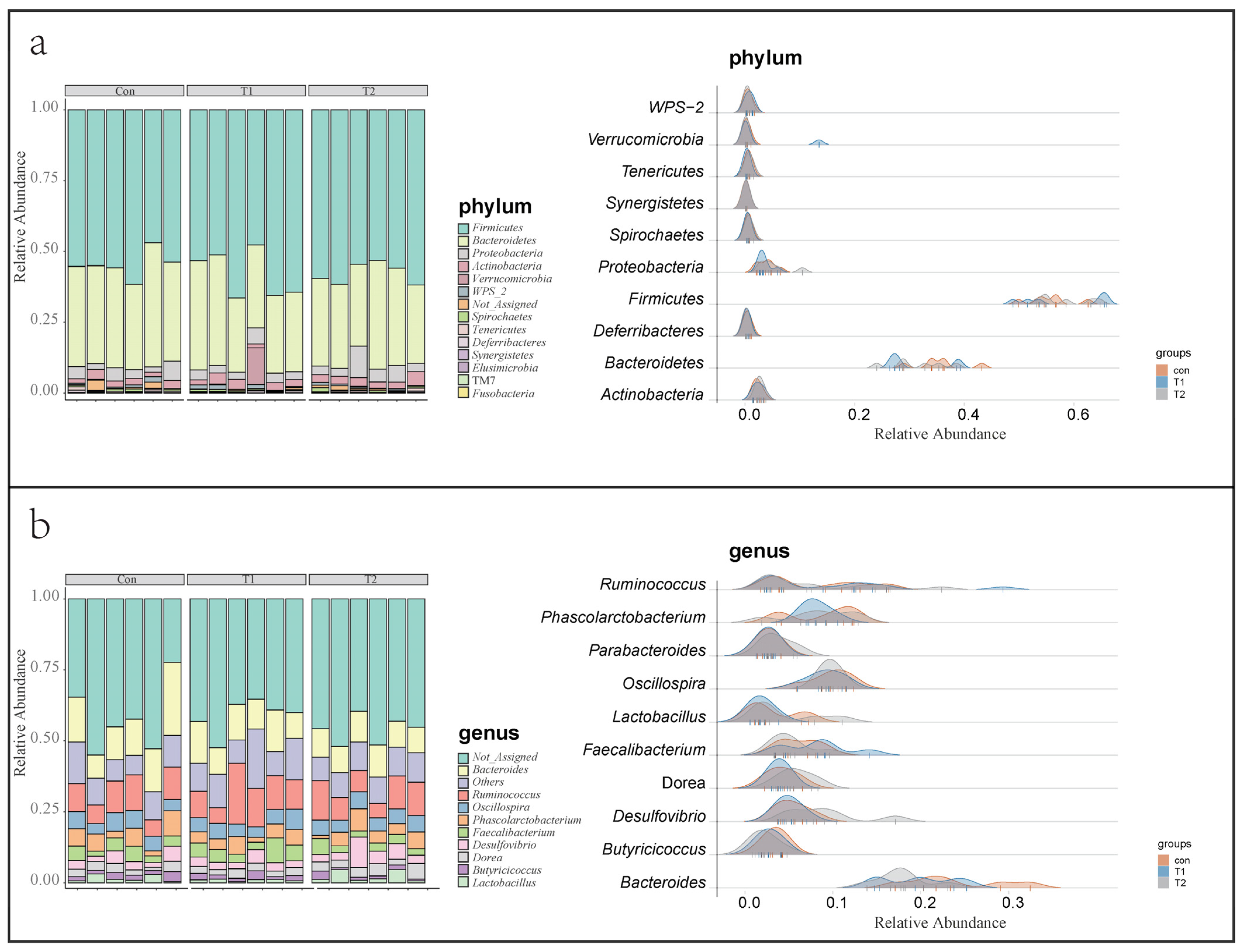
3.4. Ileal Microbial Community Structure
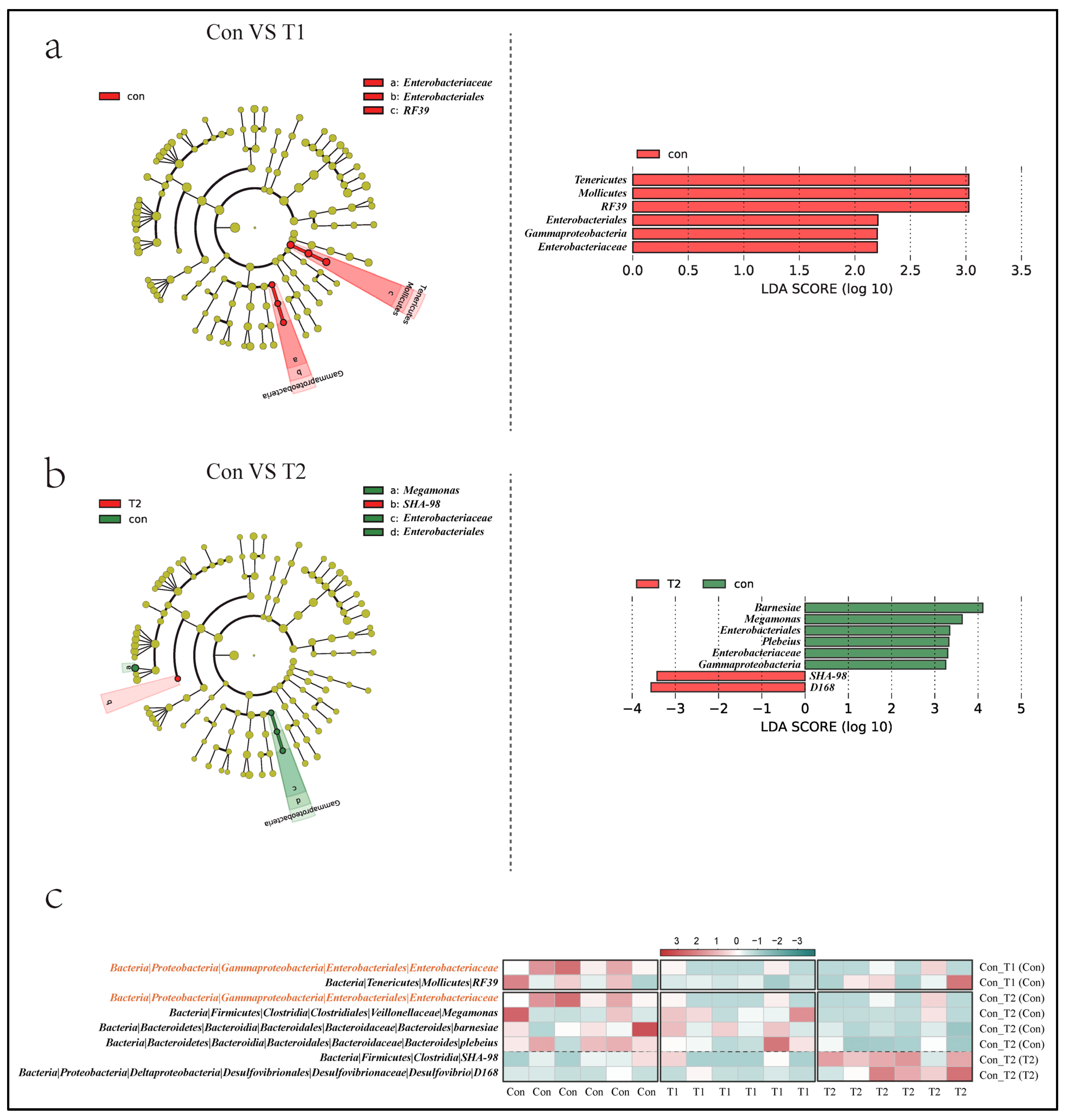
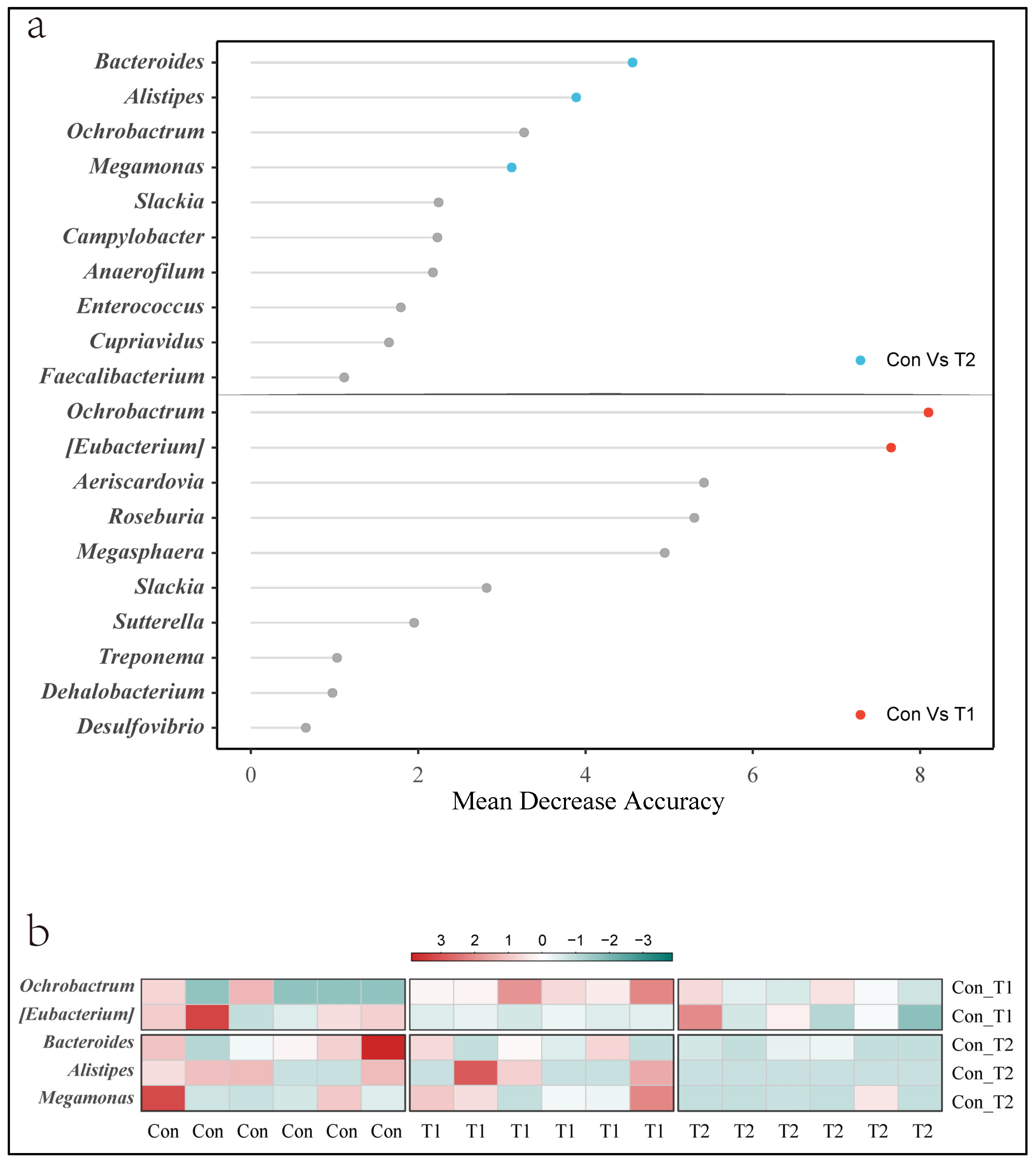
3.5. Key Microbial Identification
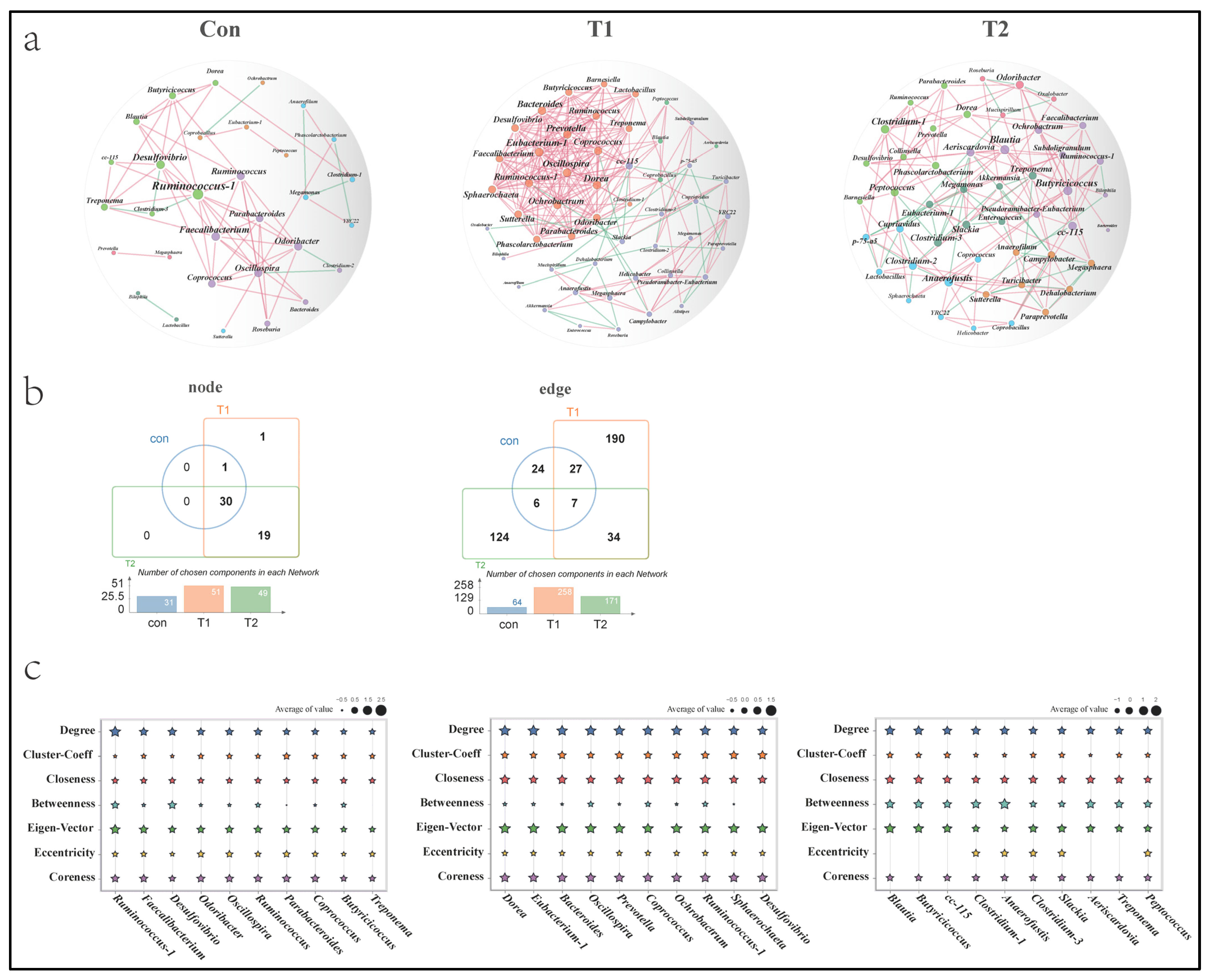
3.6. Co-Occurrence Patterns of Microbial Communities
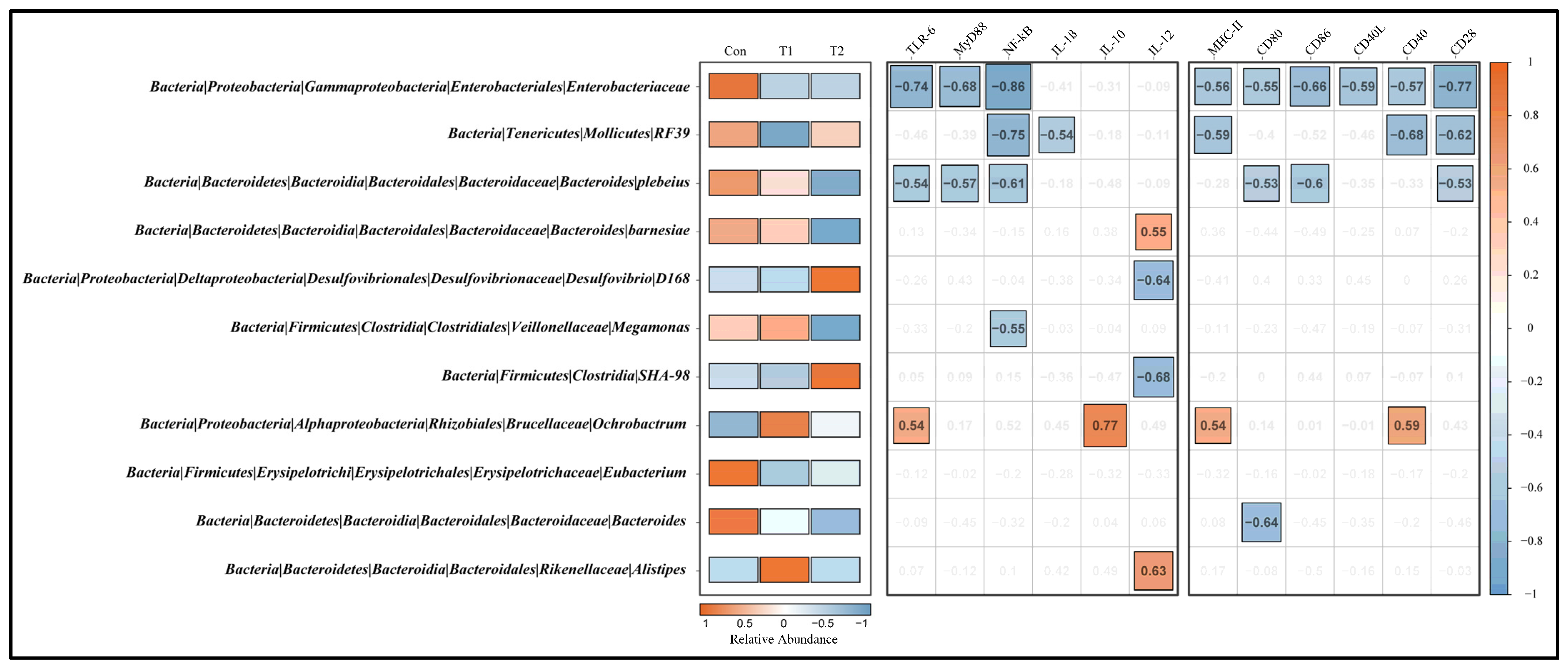
3.7. Correlation among Signature Taxa and Differential Genes in Cecal Tonsil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequences (5′–3′) | Genebank Accession |
|---|---|---|
| MHC-I | F: GCCAACACGGACCAGCAGTAC | NM_001044683.2 |
| R: CAGCATAACCTCCTTCCTCCCATTTC | ||
| MHC-II | F: AGGTGCTGGTGGTGCTGGAG | NM_001044679.2 |
| R: GCCGTCTGAGCGACTTCTTGG | ||
| CD80 | F: CTCGGGCTCGGGCTTGGG | NM_001079739.1 |
| R: GACTGGAGACTCTGAACGATACATTGG | ||
| CD86 | F: TCTTCCAGCGGAGGTTATCCAGAG | NM_001037839.1 |
| R: CTATGACTAGCGGCACTGAGACAAG | ||
| CD28 | F: CTAGTGGCTCAGCGTCCTTTGC | NM_205311.1 |
| R: TTCTTGTGTTTGGATGGGTGTCTCTC | ||
| CD40 | F: GGGCTTGTGGTGAAGGTGAAAGG | NM_204665.2 |
| R: TCGGCGATGCGGCTCTCC | ||
| CD40L | F: CAGTGAGAGTGCTGAAGTGGATGAC | NM_204733.1 |
| R: GGAGGTGCTGTGCGTGTCAAG | ||
| TLR-2 | F: CTGGGAAGTGGATTGTGGA | AB050005.2 |
| R: AAGGCGAAAGTGCGAGAAA | ||
| TLR-4 | F: GGATCTTTCAAGGTGCCACA | AY064697 |
| R: CAAGTGTCCGATGGGTAGGT | ||
| TLR-6 | F: CCAGAAGACTTGAGCGGAACACAG | NM_001081709.3 |
| R: AGGAAATGAAGGCGTGGAACTGC | ||
| MyD88 | F: TGCAAGACCATGAAGAACGA | NM_001030962.3 |
| R: TCACGGCAGCAAGAGAGATT | ||
| NF-κB | F: GTGTGAAGAAACGGGAACTG | NM_205129.1 |
| R: GGCACGGTTGTCATAGATGG | ||
| IL-1β | F: ACTGGGCATCAAGGGCTA | XM_015297469.1 |
| R: GGTAGAAGATGAAGCGGGTC | ||
| IL-2 | F: GCAGTGTTACCTGGGAGAAGTG | XM_015276098.2 |
| R: TCTTGCATTCACTTCCGGTGT | ||
| IL-4 | F: CCTCAACATGCGTCAGCTCCTG | NM_001007079.1 |
| R: GTGGAAGAAGGTACGTAGGTCTGC | ||
| IL-10 | F: CAGCACCAGTCATCAGCAGAGC | NM_001004414.2 |
| R: TCACTTCCTCCTCCTCATCAGCAG | ||
| IL-12 | F: ACTTTCCTTTGCTGCCCTTCTGG | NM_213571.1 |
| R: GAACTCCTTCACTTCGGTGGTCAG | ||
| IFN-γ | F: ACAACCTTCCTGATGGCGTG | NM_205427.1 |
| R: GAGTTCATTCGCGGCTTTGC | ||
| TNF-α | F: CTCAGGACAGCCTATGCCAACAAG | NM_204267.1 |
| R: GGCGGTCATAGAACAGCACTACG | ||
| GAPDH | F: GGCACGCCATCACTATCTTCCAG | NM_204305.1 |
| R: ACTCCACAACATACTCAGCACCTG |
Appendix B
| Item | Egg Production, % | FCR 1, g Feed/g Egg | Egg Weight, g | Feed Intake, g/d/hen | Egg Mass 4, g/d/hen | Broken Eggs, % | Mortality, % |
|---|---|---|---|---|---|---|---|
| 1–4 weeks | |||||||
| Con | 67.23 | 2.63 | 52.11 | 90.88 b | 35 | 0.17 b | 1.56 |
| T1 | 70.74 | 2.74 | 51.12 | 98.31 a | 36.21 | 0.61 a | 1.56 |
| T2 | 69.41 | 2.69 | 51.16 | 94.77 ab | 35.52 | 0.37 ab | 3.13 |
| SEM | 1.57 | 0.05 | 0.22 | 1.06 | 0.83 | 0.07 | 0.61 |
| p-value | 0.672 | 0.742 | 0.117 | 0.01 | 0.849 | 0.041 | 0.49 |
| 5–8 weeks | |||||||
| Con | 67.33 | 2.39 | 52.55 | 83.48 | 35.36 | 0.75 | 2.34 |
| T1 | 67.21 | 2.6 | 51.63 | 89.34 | 34.74 | 0.5 | 3.13 |
| T2 | 70.42 | 2.25 | 51.44 | 80.8 | 36.25 | 0.46 | 4.69 |
| SEM | 1.57 | 0.07 | 0.24 | 1.72 | 0.85 | 0.11 | 1.06 |
| p-value | 0.66 | 0.05 | 0.141 | 0.115 | 0.783 | 0.491 | 0.25 |
| 1–8 weeks | |||||||
| Con | 67.26 | 2.53 | 52.28 | 87.81 b | 35.15 | 0.42 | 3.91 |
| T1 2 | 69.3 | 2.68 | 51.32 | 94.60 a | 35.61 | 0.72 | 4.69 |
| T2 3 | 69.83 | 2.5 | 51.28 | 89.07 b | 35.82 | 0.43 | 7.81 |
| SEM | 1.5 | 0.06 | 0.23 | 1.19 | 0.8 | 0.06 | 1.32 |
| p-value | 0.778 | 0.254 | 0.116 | 0.039 | 0.945 | 0.097 | 0.31 |
Appendix C
| Item | Con | T1 | T2 | SEM | p-Value |
|---|---|---|---|---|---|
| IL-1β | 0.91 b | 1.18 a | 0.85 b | 0.044 | 0.003 |
| IL-2 | 1.06 | 1.54 | 0.92 | 0.147 | 0.236 |
| IL-4 | 1.03 | 0.7 | 0.92 | 0.062 | 0.077 |
| IL-10 | 1.06 b | 1.99 a | 0.97 b | 0.139 | <0.001 |
| IL-12 | 1.17 b | 1.49 a | 1.01 b | 0.066 | 0.003 |
| IFN-γ | 1.06 | 1.16 | 1.3 | 0.086 | 0.547 |
| TNF-α | 1.01 | 1.04 | 1.03 | 0.113 | 0.894 |
| TLR-2 | 1.3 | 1.58 | 1.47 | 0.054 | 0.096 |
| TLR-4 | 1.31 | 1.64 | 1.59 | 0.063 | 0.069 |
| TLR-6 | 1.01 b | 2.11 a | 1.48 ab | 0.157 | 0.024 |
| NF-κB | 1.00 b | 1.70 a | 1.44 a | 0.096 | 0.004 |
| MyD88 | 1.01 b | 1.78 a | 1.77 a | 0.125 | 0.005 |
| MHC-II | 1.01 b | 1.95 a | 1.24 b | 0.169 | 0.05 |
| MHC-I | 1.27 | 2.25 | 2.3 | 0.217 | 0.087 |
| CD86 | 1.02 c | 1.78 b | 2.81 a | 0.205 | <0.001 |
| CD80 | 1.02 b | 2.23 a | 2.59 a | 0.214 | 0.002 |
| CD40L | 1.03 b | 2.07 a | 1.77 a | 0.168 | 0.022 |
| CD40 | 1.02 b | 1.80 a | 1.53 a | 0.119 | 0.012 |
| CD28 | 1.01 b | 2.08 a | 2.00 a | 0.179 | 0.013 |
References
- Chen, F.; Zhang, H.; Du, E.; Jin, F.; Zheng, C.; Fan, Q.; Zhao, N.; Guo, W.; Zhang, W.; Huang, S.; et al. Effects of magnolol on egg production, egg quality, antioxidant capacity, and intestinal health of laying hens in the late phase of the laying cycle. Poult. Sci. 2021, 100, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.P.; Yan, S.J.; Li, G.; Liu, D.; Guo, Y.M. Genistein improves the reproductive performance and bone status of breeder hens during the late egg-laying period. Poult. Sci. 2019, 98, 7022–7029. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, X.; Zhen, W.; Zeng, D.; Qu, L.; Wang, Z.; Ning, Z. Yeast culture improves egg quality and reproductive performance of aged breeder layers by regulating gut microbes. Front. Microbiol. 2021, 12, 633276. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; Abo El-Maaty, H.M. Calcium and cholecalciferol levels in late-phase laying hens: Effects on productive traits, egg quality, blood biochemistry, and immune responses. Front. Vet. Sci. 2020, 7, 389. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Bai, S.P.; Zhang, K.Y.; Ding, X.M.; Wang, J.P.; Zeng, Q.F.; Peng, H.W.; Lu, H.Y.; Bai, J.; Xuan, Y.; et al. Effects of Lonicera confusa and Astragali Radix extracts supplementation on egg production performance, egg quality, sensory evaluation, and antioxidative parameters of laying hens during the late laying period. Poult. Sci. 2019, 98, 4838–4847. [Google Scholar] [CrossRef]
- Wang, X.; Ryu, D.; Houtkooper, R.H.; Auwerx, J. Antibiotic use and abuse: A threat to mitochondria and chloroplasts with impact on research, health, and environment. BioEssays News Rev. Mol. Cell. Dev. Biol. 2015, 37, 1045–1053. [Google Scholar] [CrossRef]
- Hu, R.; He, Y.; Arowolo, M.A.; Wu, S.; He, J. Polyphenols as potential attenuators of heat stress in poultry production. Antioxidants 2019, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Saint-Cyr, M.J.; Haddad, N.; Taminiau, B.; Poezevara, T.; Quesne, S.; Amelot, M.; Daube, G.; Chemaly, M.; Dousset, X.; Guyard-Nicodème, M. Use of the potential probiotic strain Lactobacillus salivarius SMXD51 to control Campylobacter jejuni in broilers. Int. J. Food Microbiol. 2017, 247, 9–17. [Google Scholar] [CrossRef]
- Guerrero Sanchez, M.; Passot, S.; Campoy, S.; Olivares, M.; Fonseca, F. Ligilactobacillus salivarius functionalities, applications, and manufacturing challenges. Appl. Microbiol. Biotechnol. 2022, 106, 57–80. [Google Scholar] [CrossRef]
- Balamuralikrishnan, B.; Lee, S.I.; Kim, I.H. Dietary inclusion of different multi-strain complex probiotics; effects on performance in broilers. Br. Poult. Sci. 2017, 58, 83–86. [Google Scholar] [CrossRef]
- Zhao, P.Y.; Kim, I.H. Effect of direct-fed microbial on growth performance, nutrient digestibility, fecal noxious gas emission, fecal microbial flora and diarrhea score in weanling pigs. Anim. Feed Sci. Technol. 2015, 200, 86–92. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Faseleh Jahromi, M.; Liang, J.B.; Ramasamy, K.; Sieo, C.C.; Ho, Y.W. Effects of a Lactobacillus salivarius mixture on performance, intestinal health and serum lipids of broiler chickens. PLoS ONE 2017, 12, e0175959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Zhou, Z.; Yu, L.; Jiang, K.; Xia, J.; Mi, Y.; Zhang, C.; Li, J. Lactobacillus salivarius and Lactobacillus agilis feeding regulates intestinal stem cells activity by modulating crypt niche in hens. Appl. Microbiol. Biotechnol. 2021, 105, 8823–8835. [Google Scholar] [CrossRef] [PubMed]
- Blajman, J.E.; Olivero, C.A.; Fusari, M.L.; Zimmermann, J.A.; Rossler, E.; Berisvil, A.P.; Romero Scharpen, A.; Astesana, D.M.; Soto, L.P.; Signorini, M.L.; et al. Impact of lyophilized Lactobacillus salivarius DSPV 001P administration on growth performance, microbial translocation, and gastrointestinal microbiota of broilers reared under low ambient temperature. Res. Vet. Sci. 2017, 114, 388–394. [Google Scholar] [CrossRef]
- Messaoudi, S.; Manai, M.; Kergourlay, G.; Prévost, H.; Connil, N.; Chobert, J.M.; Dousset, X. Lactobacillus salivarius: Bacteriocin and probiotic activity. Food microbiology 2013, 36, 296–304. [Google Scholar] [CrossRef]
- Xu, C.; Wei, F.; Yang, X.; Feng, Y.; Liu, D.; Hu, Y. Lactobacillus salivarius CML352 isolated from Chinese local breed chicken modulates the gut microbiota and improves intestinal health and egg quality in late-phase laying hens. Microorganisms 2022, 10, 726. [Google Scholar] [CrossRef]
- Wang, J.; Ishfaq, M.; Guo, Y.; Chen, C.; Li, J. Assessment of Probiotic Properties of Lactobacillus salivarius Isolated from Chickens as Feed Additives. Front. Vet. Sci. 2020, 7, 415. [Google Scholar] [CrossRef]
- Wang, J.; Ishfaq, M.; Li, J. Lactobacillus salivarius ameliorated Mycoplasma gallisepticum-induced inflammatory injury and secondary Escherichia coli infection in chickens: Involvement of intestinal microbiota. Vet. Immunol. Immunopathol. 2021, 233, 110192. [Google Scholar] [CrossRef]
- Deng, J.; Li, Y.; Zhang, J.; Yang, Q. Co-administration of Bacillus subtilis RJGP16 and Lactobacillus salivarius B1 strongly enhances the intestinal mucosal immunity of piglets. Res. Vet. Sci. 2013, 94, 62–68. [Google Scholar] [CrossRef]
- Zhang, J.; Deng, J.; Wang, Z.; Che, C.; Li, Y.F.; Yang, Q. Modulatory effects of Lactobacillus salivarius on intestinal mucosal immunity of piglets. Curr. Microbiol. 2011, 62, 1623–1631. [Google Scholar] [CrossRef]
- Ren, D.; Wang, D.; Liu, H.; Shen, M.; Yu, H. Two strains of probiotic Lactobacillus enhance immune response and promote naive T cell polarization to Th1. Food Agric. Immunol. 2019, 30, 281–295. [Google Scholar] [CrossRef] [Green Version]
- China National Standard. Feeding Standard of Chicken (NY/T 33-2004); Ministry of Agriculture: Beijing, China, 2004.
- Zhu, Z.; Yang, L.; Yu, P.; Wang, Y.; Peng, X.; Chen, L. Comparative Proteomics and Secretomics Revealed Virulence and Antibiotic Resistance-Associated Factors in Vibrio parahaemolyticus Recovered From Commonly Consumed Aquatic Products. Front. Microbiol. 2020, 11, 1453. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, S.; Singh, R.; Yadav, D.; Mande, S.S. MetagenoNets: Comprehensive inference and meta-insights for microbial correlation networks. Nucleic Acids Res. 2020, 48, W572–W579. [Google Scholar] [CrossRef]
- Archer, E. Package ‘rfPermute’; R Project: Indianapolis, IN, USA, 2016. [Google Scholar]
- Kursa, M.B.; Rudnicki, W.R. Feature selection with the Boruta package. J. Stat. Softw. 2010, 36, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.Y.; Chen, J.F.; He, C.Q.; Chi, F.; Johnston, S.L. Effects of modified montmorillonite adsorbent on performance, egg quality, serum biochemistry, oxidation status, and immune response of laying hens in late production. Livest. Sci. 2018, 210, 15–20. [Google Scholar] [CrossRef]
- Tiedje, J.M.; Wang, F.; Manaia, C.M.; Virta, M.; Sheng, H.; Ma, L.; Zhang, T.; Topp, E. Antibiotic resistance genes in the human-impacted environment: A one health perspective. Pedosphere 2019, 29, 273–282. [Google Scholar] [CrossRef]
- Befus, A.D.; Johnston, N.; Leslie, G.A.; Bienenstock, J. Gut-associated lymphoid tissue in the chicken. I. Morphology, ontogeny, and some functional characteristics of Peyer’s patches. J. Immunol. 1980, 125, 2626–2632. [Google Scholar]
- Setta, A.M.; Barrow, P.A.; Kaiser, P.; Jones, M.A. Early immune dynamics following infection with Salmonella enterica serovars Enteritidis, Infantis, Pullorum and Gallinarum: Cytokine and chemokine gene expression profile and cellular changes of chicken cecal tonsils. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 397–410. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.; Edidin, M.; Almo, S.C.; Nathenson, S.G. B7-1 and B7-2: Similar costimulatory ligands with different biochemical, oligomeric and signaling properties. Immunol. Lett. 2006, 104, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Khalique, A.; Zeng, D.; Wang, H.; Qing, X.; Zhou, Y.; Xin, J.; Zeng, Y.; Pan, K.; Shu, G.; Jing, B.; et al. Transcriptome analysis revealed ameliorative effect of probiotic Lactobacillus johnsonii BS15 against subclinical necrotic enteritis induced hepatic inflammation in broilers. Microb. Pathog. 2019, 132, 201–207. [Google Scholar] [CrossRef]
- Satpathy, A.T.; Wu, X.; Albring, J.C.; Murphy, K.M. Re(de)fining the dendritic cell lineage. Nat. Immunol. 2012, 13, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Taha-Abdelaziz, K.; Astill, J.; Kulkarni, R.R.; Read, L.R.; Najarian, A.; Farber, J.M.; Sharif, S. In vitro assessment of immunomodulatory and anti-Campylobacter activities of probiotic lactobacilli. Sci. Rep. 2019, 9, 17903. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Brisbin, J.T.; Hodgins, D.C.; Sharif, S. Lactobacillus and Lactobacillus cell-free culture supernatants modulate chicken macrophage activities. Res. Vet. Sci. 2015, 103, 170–175. [Google Scholar] [CrossRef]
- Oláh, I.; Nagy, N.; Vervelde, L. Structure of the avian lymphoid system. In Avian Immunology, 2nd ed.; Schat, K.A., Kaspers, B., Kaiser, P., Eds.; Academic Press: Boston, NY, USA, 2014; Chapter 2; pp. 11–44. [Google Scholar]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.P.; Pembroke, J.T. The Genus Ochrobactrum as major opportunistic pathogens. Microorganisms 2020, 8, 1797. [Google Scholar] [CrossRef] [PubMed]
- Barquero-Calvo, E.; Conde-Alvarez, R.; Chacón-Díaz, C.; Quesada-Lobo, L.; Martirosyan, A.; Guzmán-Verri, C.; Iriarte, M.; Mancek-Keber, M.; Jerala, R.; Gorvel, J.P.; et al. The differential interaction of Brucella and ochrobactrum with innate immunity reveals traits related to the evolution of stealthy pathogens. PLoS ONE 2009, 4, e5893. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef]
- Vieira, A.T.; Vinolo, M.A.R. Regulation of immune cell function by short chain fatty acids and their impact on arthritis. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases, 2nd ed.; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; Chapter 9; pp. 175–188. [Google Scholar]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Nguyen, S.M.; Yang, Y.; Xu, W.; Cai, H.; Wu, J.; Cai, Q.; Long, J.; Zheng, W.; Shu, X.O. Long-term diet quality is associated with gut microbiome diversity and composition among urban Chinese adults. Am. J. Clin. Nutr. 2021, 113, 684–694. [Google Scholar] [CrossRef]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and disease markers correlate with gut microbiome composition across thousands of people. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef]
- Ericson, U.; Brunkwall, L.; Hellstrand, S.; Nilsson, P.M.; Orho-Melander, M. A health-conscious food pattern is associated with prediabetes and gut microbiota in the Malmö Offspring Study. J. Nutr. 2020, 150, 861–872. [Google Scholar] [CrossRef]
- Zumla, A. Mandell, Douglas, and Bennett’s principles and practice of infectious diseases. Lancet Infect. Dis. 2010, 10, 303–304. [Google Scholar] [CrossRef]
- Lippert, K.; Kedenko, L.; Antonielli, L.; Kedenko, I.; Gemeier, C.; Leitner, M.; Kautzky-Willer, A.; Paulweber, B.; Hackl, E. Gut microbiota dysbiosis associated with glucose metabolism disorders and the metabolic syndrome in older adults. Benef. Microbes 2017, 8, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Zhang, K.; Ma, X.; He, P. Clostridium species as probiotics: Potentials and challenges. J. Anim. Sci. Biotechnol. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Chevrot, R.; Carlotti, A.; Sopena, V.; Marchand, P.; Rosenfeld, E. Megamonas rupellensis sp. nov., an anaerobe isolated from the caecum of a duck. Int. J. Syst. Evol. Microbiol. 2008, 58, 2921–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakon, H.; Nagai, F.; Morotomi, M.; Tanaka, R. Sutterella parvirubra sp. nov. and Megamonas funiformis sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jiang, Q.; Liu, Z.; Shen, S.; Ai, J.; Zhu, Y.; Zhou, L. Alteration of Gut Microbiota Relates to Metabolic Disorders in Primary Aldosteronism Patients. Front. Endocrinol. 2021, 12, 667951. [Google Scholar] [CrossRef]
- Kumar, S.; Chen, C.; Indugu, N.; Werlang, G.O.; Singh, M.; Kim, W.K.; Thippareddi, H. Effect of antibiotic withdrawal in feed on chicken gut microbial dynamics, immunity, growth performance and prevalence of foodborne pathogens. PLoS ONE 2018, 13, e0192450. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.L.; Chung, C.Y.; Kuo, C.H.; Kuo, T.F.; Yang, C.W.; Yang, W.C. Beneficial effect of Bidens pilosa on body weight gain, food conversion ratio, gut bacteria and coccidiosis in chickens. PLoS ONE 2016, 11, e0146141. [Google Scholar] [CrossRef]
- Wrigley, D.M. Inhibition of Clostridium perfringens sporulation by Bacteroides fragilis and short-chain fatty acids. Anaerobe 2004, 10, 295–300. [Google Scholar] [CrossRef]






| Ingredients | Percent (%) | Nutrient Levels | Content |
|---|---|---|---|
| Corn (CP 8.3%) | 64.00 | ME (MJ/Kg) | 11.16 |
| Soybean meal (CP 44.0%) | 19.80 | CP (%) | 15.00 |
| Soybean oil | 0.70 | CF (%) | 2.33 |
| Wheat bran (CP 14.3%) | 4.11 | Met (%) | 0.33 |
| Limestone | 9.50 | Lys (%) | 0.77 |
| Calcium hydrogen phosphate | 1.00 | Met + Cys (%) | 0.67 |
| Sodium chloride | 0.30 | Trp (%) | 0.17 |
| DL-Methionine (98%) | 0.10 | Thr (%) | 0.59 |
| L-Lysine HCL (78%) | 0.07 | Calcium (%) | 3.72 |
| Vitamin premix a | 0.03 | Total p (%) | 0.52 |
| Mineral premix b | 0.20 | Available p (%) | 0.34 |
| Choline chloride (50%) | 0.15 | ||
| Phytase | 0.02 | ||
| NSP enzymes | 0.02 | ||
| Total | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, L.; Yan, H.; Ning, Z.; Wang, Z. Lactobacillus salivarius SNK-6 Activates Intestinal Mucosal Immune System by Regulating Cecal Microbial Community Structure in Laying Hens. Microorganisms 2022, 10, 1469. https://doi.org/10.3390/microorganisms10071469
Liu Y, Li L, Yan H, Ning Z, Wang Z. Lactobacillus salivarius SNK-6 Activates Intestinal Mucosal Immune System by Regulating Cecal Microbial Community Structure in Laying Hens. Microorganisms. 2022; 10(7):1469. https://doi.org/10.3390/microorganisms10071469
Chicago/Turabian StyleLiu, Yuchen, Lianhong Li, Huaxiang Yan, Zhonghua Ning, and Zhong Wang. 2022. "Lactobacillus salivarius SNK-6 Activates Intestinal Mucosal Immune System by Regulating Cecal Microbial Community Structure in Laying Hens" Microorganisms 10, no. 7: 1469. https://doi.org/10.3390/microorganisms10071469