
Conidial Emulsion Formulation and Thermal Storability of Metarhizium anisopliae against Red Palm Weevil, Rhynchophorusferrugineus Olivier (Coleoptera: Dryophthoridae)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Active and Inert Ingredient Compatibility Screening
2.3. Emulsion Formulation Preparation
2.4. Incorporation of Conidia in Emulsion System
2.5. Characterization of Emulsion Formulations
2.6. Effect of Storage Temperature on Conidial Viability
2.7. Effect of Heat Stress on Conidial Viability
2.8. Statistical Analyses
3. Results
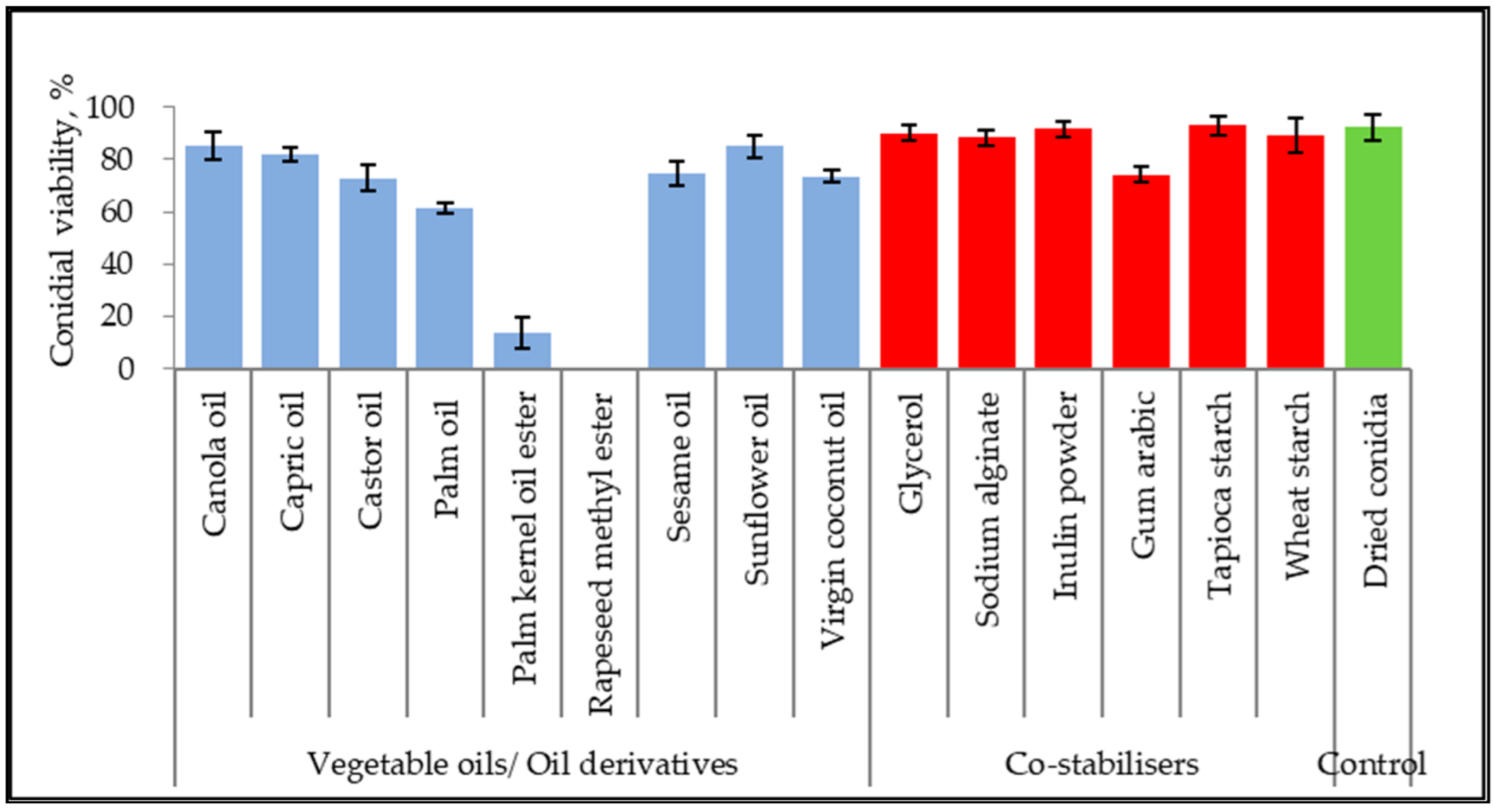
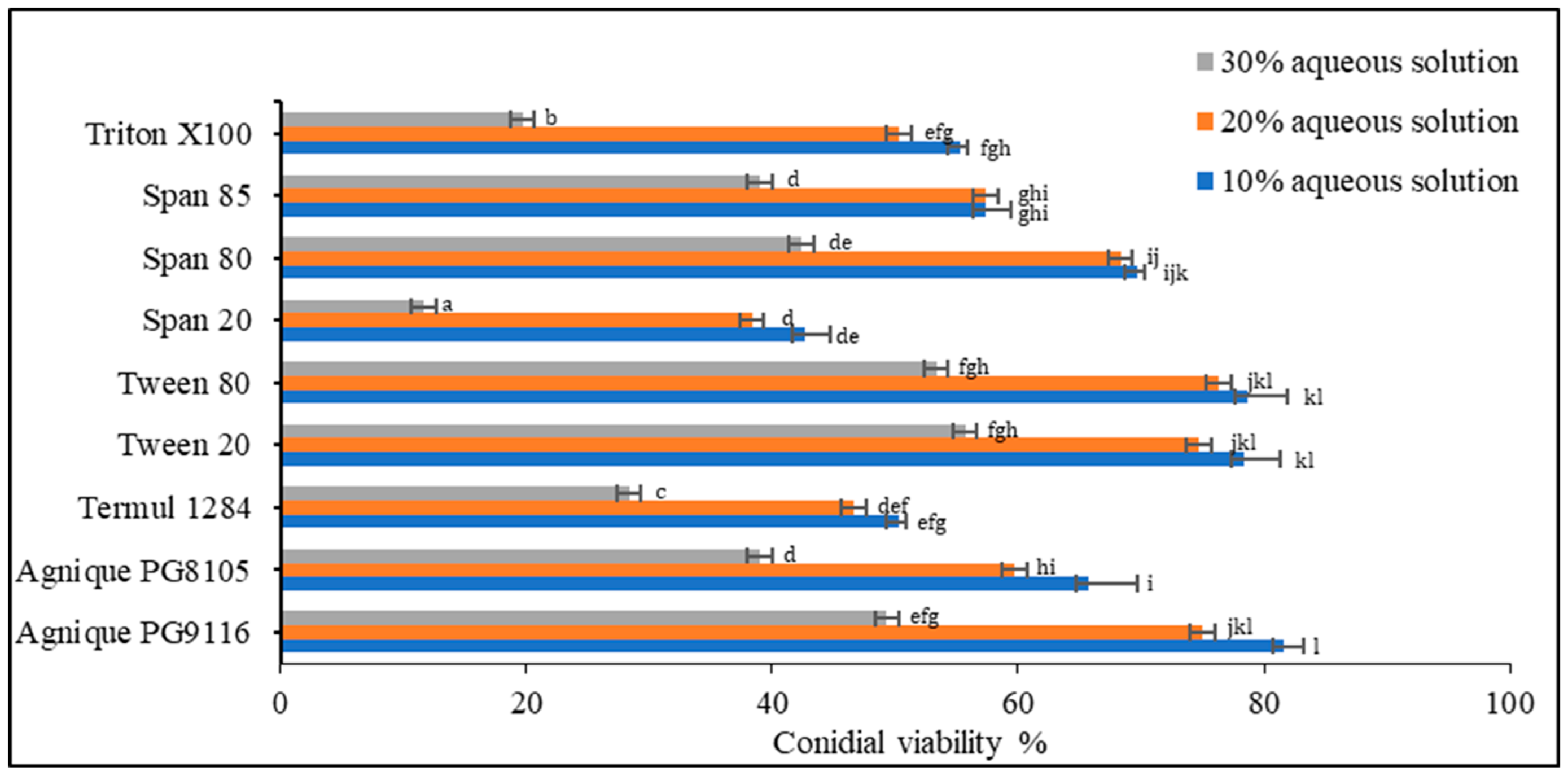
3.1. Inert Ingredient Screening
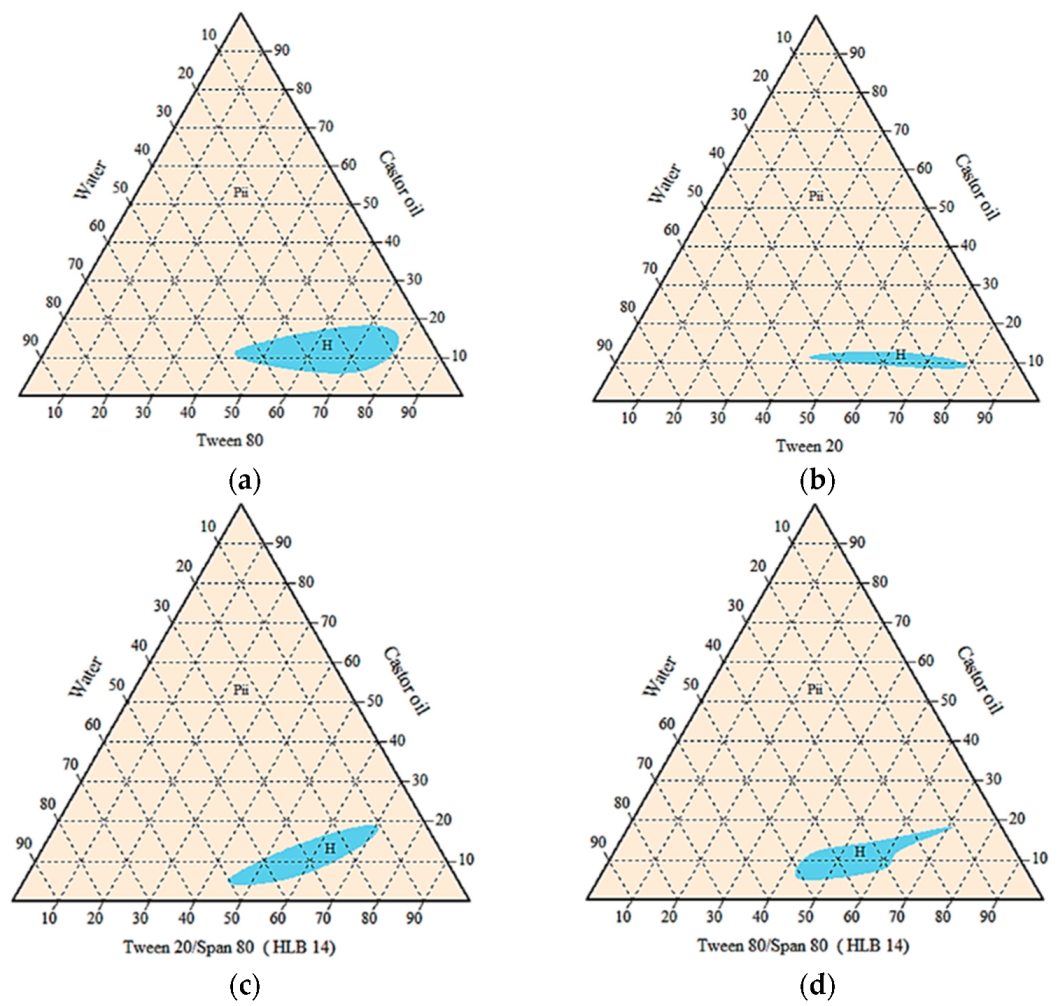
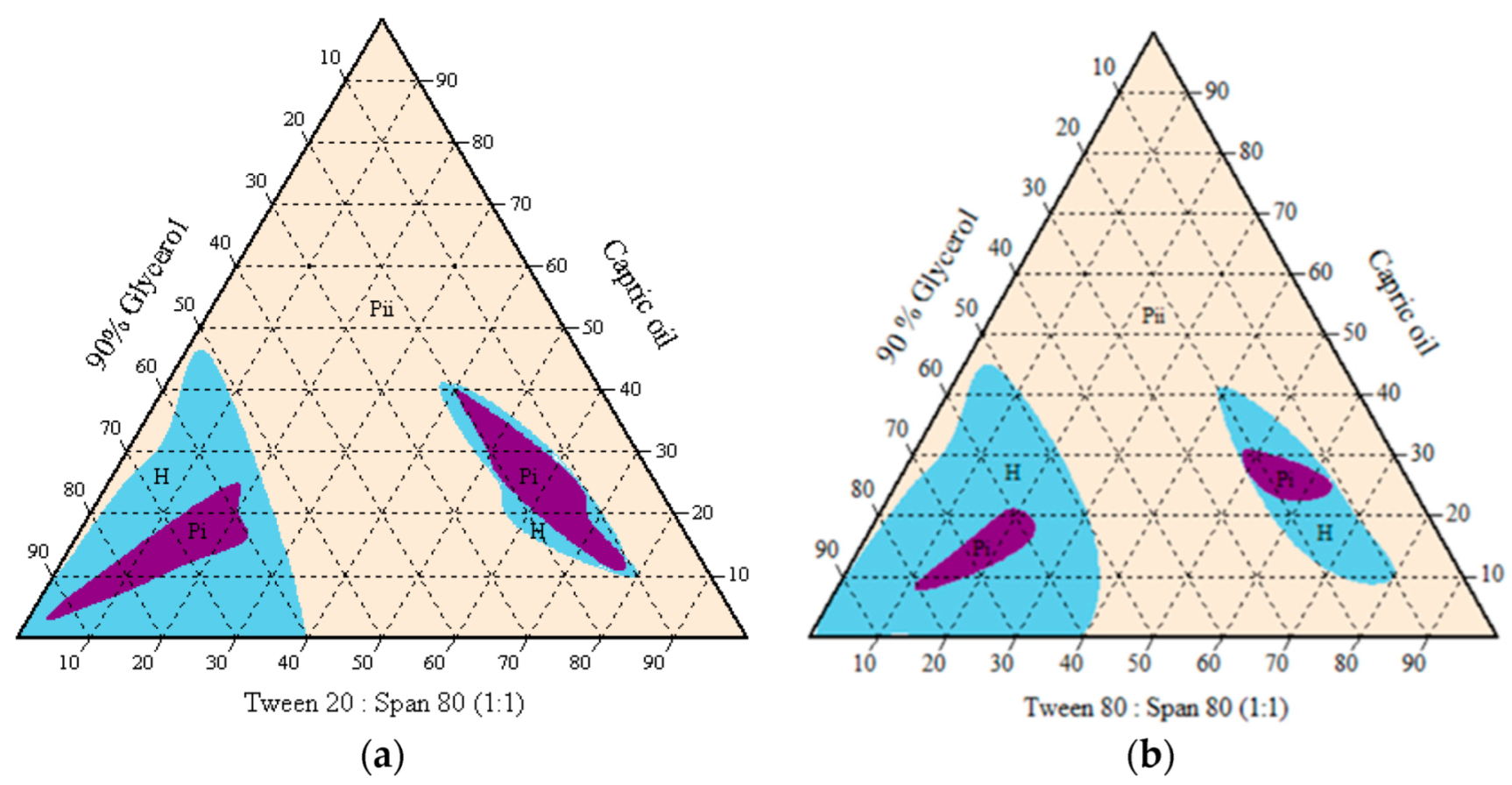
3.2. Phase Behaviour Study of the Mixtures
3.3. Characterization of Emulsion Formulations
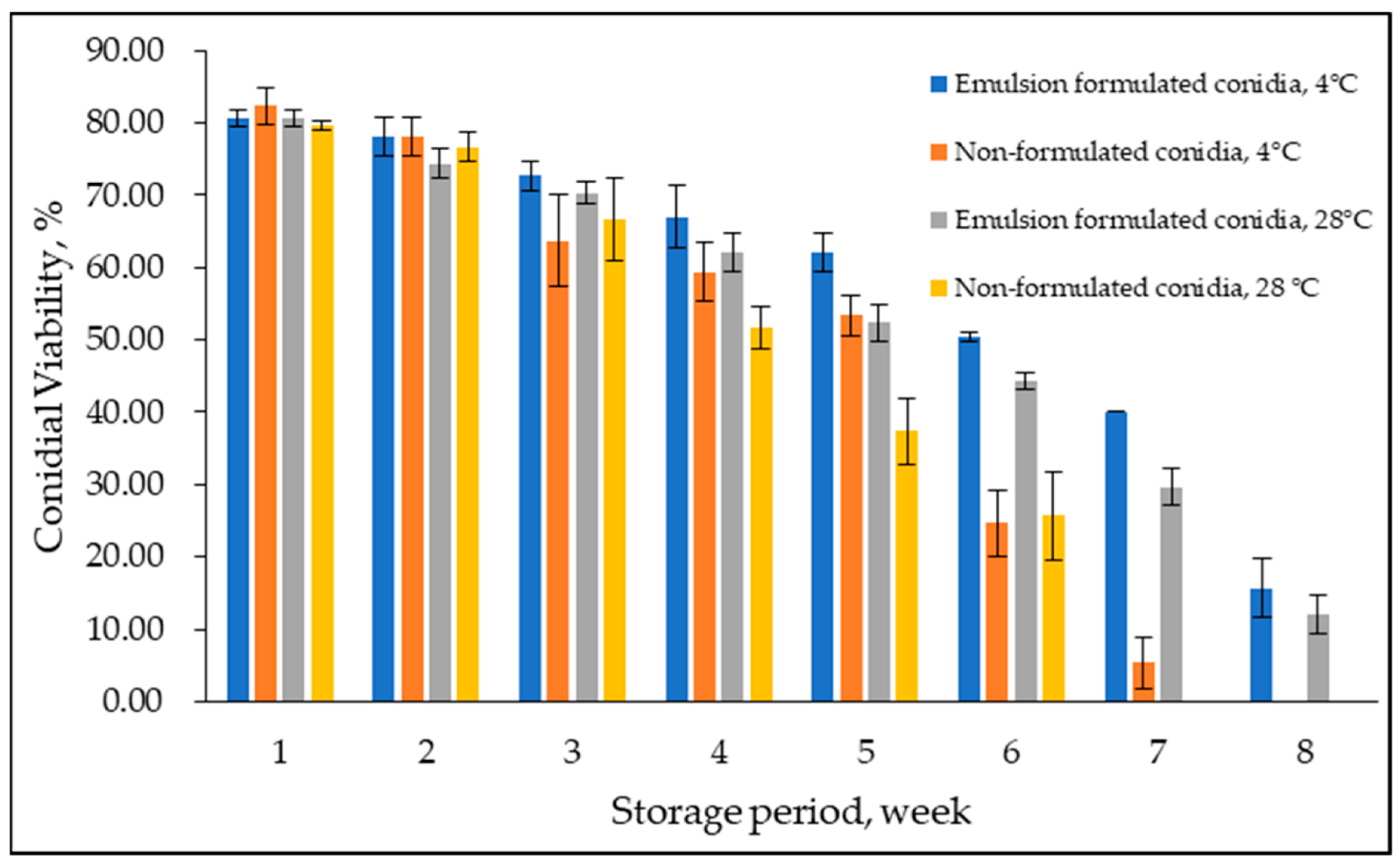
3.4. Conidial Viability at Storage Temperature
3.5. Conidial Viability under Heat Stress
4. Discussion
4.1. Development of Emulsion Formulation
4.2. Thermal Storability of Conidial Emulsion Formulation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wahizatul, A.A.; Chong, J.L.; Hazlina, A.Z.; Norhayati, Y.; Wan Bayani, W.O.; Yong, K.W.; Ainatun, N.Z.; Mohd Haris, H. The red palm weevil, Rhynchophorus ferrugineus: Current issues and challenges in Malaysia. Oil Palm Bull. 2017, 74, 17–24. [Google Scholar]
- Rahman, A.A. Pesticides residue analysis of fruits for farm accreditation schemes in Sabah, Malaysia. TFNet 2020, 123, 28–31. [Google Scholar]
- Ehler, L.E. Integrated pest management (IPM): Definition, historical development and implementation, and the other IPM. Pest Manag. Sci. 2006, 62, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Omukoko, C.A.; Wesonga, J.M.; Maniania, K.N.; Wamocho, L.S.; Kahangi, E.M. Screening of Beauveria bassiana isolates to the banana weevil and horizontal transmission under laboratory conditions. J. Agric. Sci. Technol. 2014, 16, 1–12. [Google Scholar]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Use of hyphomycetous fungi for managing insect pests. In Fungi as Biocontrol Agents; Butt, T.M., Jackson, C., Magan, N., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 23–69. [Google Scholar]
- Alves, R.T.; Bateman, R.P.; Gunn, J.; Prior, C.; Leather, S.R. Effects of different formulations on viability and medium-term storage of Metarhizium anisopliae conidia. Neotrop. Entomol. 2002, 31, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Athanassiou, C.G.; Kavallieratos, N.G.; Rumbos, C.I.; Kontodimas, D.C. Influence of temperature and relative humidity on the insecticidal efficacy of Metarhizium anisopliae against larvae of Ephestia kuehniella (Lepidoptera: Pyralidae) on wheat. J. Insect Sci. 2017, 17, 1–7. [Google Scholar] [CrossRef]
- Fernandes, É.K.; Bittencourt, V.R.; Roberts, D.W. Perspectives on the potential of entomopathogenic fungi in biological control of ticks. Exp. Parasitol. 2012, 130, 300–305. [Google Scholar] [CrossRef]
- de la Cruz Quiroz, R.; Maldonado, J.J.C.; Alanis, M.D.J.R.; Torres, J.A.; Saldívar, R.P. Fungi-based biopesticides: Shelf-life preservation technologies used in commercial products. J. Pest Sci. 2019, 92, 1003–1015. [Google Scholar] [CrossRef]
- Awan, U.A.; Meng, L.; Xia, S.; Raza, M.F.; Zhang, Z.; Zhang, H. Isolation, fermentation, and formulation of entomopathogenic fungi virulent against adults of Diaphorina citri. Pest Manag. Sci. 2021, 77, 4040–4053. [Google Scholar] [CrossRef]
- Olabiyi, D.O.; Duren, E.B.; Price, T.; Avery, P.B.; Hahn, P.G.; Stelinski, L.L.; Diepenbrock, L.M. Suitability of formulated entomopathogenic fungi against hibiscus mealybug, Nipaecoccus viridis (Hemiptera: Pseudococcidae), deployed within mesh covers intended to protect citrus from Huanglongbing. J. Econ. Entomol. 2021, 115, 212–223. [Google Scholar] [CrossRef]
- Corval, A.R.; Mesquita, E.; Corrêa, T.A.; Silva, C.D.S.; Bitencourt, R.D.O.; Fernandes, É.K.; Gôlo, P.S. UV-B tolerances of conidia, blastospores, and microsclerotia of Metarhizium spp. entomopathogenic fungi. J. Basic Microbiol. 2021, 61, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, M.; Pazhanisamy, M.; Sathyaseelan, V. Efficacy of oil and granular based formulations of entomopathogenic fungi, Zoophthora radicans against the biology of rice leaf folder, Cnaphalocrocis medinalis. J. Biopestic. 2019, 12, 139–144. [Google Scholar]
- Pathak, N.; Bhaduri, A.; Rai, A.K. Sesame: Bioactive compounds and health benefits. In Bioactive Molecules in Food; Mérillon, J.M., Ramawat, K.G., Eds.; Springer International Publishing: New York, NY, USA, 2019; pp. 1–20. [Google Scholar]
- Grace, L.E.L.; Jamilah, M.S.H.; Mohd, F.A.; Wahizatul, A.A. Entomopathogenic fungi isolated from the soil of Terengganu, Malaysia as potential bio-pesticides against the red palm weevil, Rhynchophorus ferrugineus. J. Sustain. Sci. Manag. 2017, 12, 71–79. [Google Scholar]
- Yang, J.; Gu, Z.; Cheng, L.; Li, Z.; Li, C.; Ban, X.; Hong, Y. Preparation and stability mechanisms of double emulsions stabilized by gelatinized native starch. Carbohydr. Polym. 2021, 262, 117926. [Google Scholar] [CrossRef]
- Chouhan, P.; Saini, T.R. D-optimal design and development of microemulsion based transungual drug delivery formulation of ciclopirox olamine for treatment of onychomycosis. Indian J. Pharm. Sci. 2016, 78, 498–511. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, D.G.P.; Lopes, R.B.; Rezende, J.M.; Delalibera, I., Jr. Increased tolerance of Beauveria bassiana and Metarhizium anisopliae conidia to high temperature provided by oil-based formulations. J. Invertebr. Pathol. 2018, 151, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soberón, M.; Bravo, A.; Blanco, C.A. Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Gurpreet, K.; Singh, S.K. Review of nanoemulsion formulation and characterization techniques. Indian J. Pharm. Sci. 2018, 80, 781–789. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoemulsions versus microemulsions: Terminology, differences, and similarities. Soft Matter 2012, 8, 1719–1729. [Google Scholar] [CrossRef]
- Rao, J.; McClements, D.J. Lemon oil solubilization in mixed surfactant solutions: Rationalizing microemulsion & nanoemulsion formation. Food Hydrocoll. 2012, 26, 268–276. [Google Scholar]
- Pavoni, L.; Perinelli, D.R.; Bonacucina, G.; Cespi, M.; Palmieri, G.F. An overview of micro-and nanoemulsions as vehicles for essential oils: Formulation, preparation and stability. Nanomaterials 2020, 10, 135. [Google Scholar] [CrossRef] [Green Version]
- Lv, G.; Wang, F.; Cai, W.; Zhang, X. Characterization of the addition of lipophilic Span 80 to the hydrophilic Tween 80-stabilized emulsions. Colloids Surf. A Physicochem. Eng. Asp. 2014, 447, 8–13. [Google Scholar] [CrossRef]
- Stang, M.; Schuchmann, H.; Schubert, H. Emulsification in high-pressure homogenizers. Eng. Life Sci. 2001, 1, 151–157. [Google Scholar] [CrossRef]
- Wooster, T.J.; Golding, M.; Sanguansri, P. Impact of oil type on nanoemulsion formation and Ostwald ripening stability. Langmuir 2008, 24, 12758–12765. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Li, Y.; McClements, D.J.; Xiao, H. Nanoemulsion-and emulsion-based delivery systems for curcumin: Encapsulation and release properties. Food Chem. 2012, 132, 799–807. [Google Scholar] [CrossRef]
- Taha, A.; Hu, T.; Zhang, Z.; Bakry, A.M.; Khalifa, I.; Pan, S.; Hu, H. Effect of different oils and ultrasound emulsification conditions on the physicochemical properties of emulsions stabilized by soy protein isolate. Ultrason. Sonochem. 2018, 49, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Han, S.W.; Song, H.Y.; Moon, T.W.; Choi, S.J. Influence of emulsion interfacial membrane characteristics on Ostwald ripening in a model emulsion. Food Chem. 2018, 242, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Meroni, E.; Raikos, V. Physicochemical stability, antioxidant properties and bioaccessibility of β-carotene in orange oil-in-water beverage emulsions: Influence of carrier oil types. Food Funct. 2018, 9, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Van Westen, T.; Groot, R.D. Effect of temperature cycling on Ostwald ripening. Cryst. Growth Des. 2018, 18, 4952–4962. [Google Scholar] [CrossRef] [Green Version]
- Patel, H.; Raval, G.; Nazari, M.; Heerklotz, H. Effects of glycerol and urea on micellization, membrane partitioning and solubilization by a non-ionic surfactant. Biophys. Chem. 2010, 150, 119–128. [Google Scholar] [CrossRef]
- Mustafa, I.F.; Hussein, M.Z. Synthesis and technology of nanoemulsion-based pesticide formulation. Nanomaterials 2020, 10, 1608. [Google Scholar] [CrossRef]
- Kang, S.W.; Kim, Y.B.; Shin, J.D.; Kim, E.K. Enhanced biodegradation of hydrocarbons in soil by microbial biosurfactant, sophorolipid. Appl. Biochem. Biotechnol. 2010, 160, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Mehta, S.K. Developments of Polysorbate (Tween) based microemulsions: Preclinical drug delivery, toxicity and antimicrobial applications. Int. J. Pharm. 2017, 529, 134–160. [Google Scholar] [CrossRef] [PubMed]
- Rahate, A.R.; Nagarkar, J.M. Emulsification of vegetable oils using a blend of non-ionic surfactants for cosmetic applications. J. Dispers. Sci. Technol. 2007, 28, 1077–1080. [Google Scholar] [CrossRef]
- Porras, M.; Solans, C.; Gonzalez, C.; Gutierrez, J.M. Properties of water-in-oil (W/O) nano-emulsions prepared by a low-energy emulsification method. Colloids Surf. A Physicochem. Eng. Asp. 2008, 324, 181–188. [Google Scholar] [CrossRef]
- Pan, H.; Yu, L.; Xu, J.; Sun, D. Preparation of highly stable concentrated W/O nanoemulsions by PIC method at elevated temperature. Colloids Surf. A Physicochem. Eng. Asp. 2014, 447, 97–102. [Google Scholar] [CrossRef]
- Yi, J.; Li, Y.; Zhong, F.; Yokoyama, W. The physicochemical stability and in vitro bioaccessibility of beta-carotene in oil-in-water sodium caseinate emulsions. Food Hydrocoll. 2014, 35, 19–27. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Sun, P.; Yang, C. Pickering emulsions stabilized by native starch granules. Colloids Surf. A Physicochem. Eng. Asp. 2013, 431, 142–149. [Google Scholar] [CrossRef]
- Leal-Calderon, F.; Schmitt, V. Solid-stabilized emulsions. Curr. Opin. Colloid Interface Sci. 2008, 13, 217–227. [Google Scholar] [CrossRef]
- Lazzarini, G.M.J.; Rocha, L.F.N.; Luz, C. Impact of moisture on in vitro germination of Metarhizium anisopliae and Beauveria bassiana and their activity on Triatoma infestans. Mycol. Res. 2006, 110, 485–492. [Google Scholar] [CrossRef]
- Birnbaum, N.; Reingold, V.; Matveev, S.; Kottakota, C.; Davidovitz, M.; Mani, K.A.; Ment, D. Not Only a Formulation: The Effects of Pickering Emulsion on the Entomopathogenic Action of Metarhizium brunneum. J. Fungi 2021, 7, 499. [Google Scholar] [CrossRef]
- Xavier-Santos, S.; Lopes, R.B.; Faria, M. Emulsifiable oils protect Metarhizium robertsii and Metarhizium pingshaense conidia from imbibitional damage. Biol. Control 2011, 5, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.J.; Smith, D.; Bridge, P.D.; Jeffries, P. The relationship between fungal preservation method and secondary metabolite production in Metarhizium anisopliae and Fusarium oxysporum. World J. Microbiol. Biotechnol. 2003, 19, 839–844. [Google Scholar] [CrossRef]
- Stevenson, A.; Hamill, P.G.; Medina, Á.; Kminek, G.; Rummel, J.D.; Dijksterhuis, J.; Hallsworth, J.E. Glycerol enhances fungal germination at the water-activity limit for life. Environ. Microbiol. 2017, 19, 947–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boruah, S.; Dutta, P.; Puzari, K.C.; Hazarika, G.N. Liquid bioformulation of Metarhizium anisopliae is effective for the management of cowpea mosaic disease. Int. J. Appl. Biol. Pharm. 2015, 6, 178–185. [Google Scholar]
- Batta, Y.A. Production and testing of novel formulations of the entomopathogenic fungus Metarhizium anisopliae (Metschinkoff) Sorokin (Deuteromycotina: Hyphomycetes). Crop Prot. 2003, 22, 415–422. [Google Scholar] [CrossRef]
- Buteler, M.; Stadler, T. A Review on the mode of action and current use of petroleum distilled spray oils. In Pesticides in the Modern World; Stoycheva, M., Ed.; InTech: London, UK, 2011; pp. 119–136. [Google Scholar]
- Lopes, R.B.; Faria, M. Influence of two formulation types and moisture levels on the storage stability and insecticidal activity of Beauveria bassiana. Biocontrol Sci. Technol. 2019, 29, 437–450. [Google Scholar] [CrossRef]
- Inyang, E.N.; McCartney, H.A.; Oyejola, B.; Ibrahim, L.; Archer, S.A. Effect of formulation, application and rain on the persistence of the entomogenous fungus Metarhizium anisopliae on oilseed rape. Mycol. Res. 2000, 104, 653–661. [Google Scholar] [CrossRef]
- Murphy, S.T.; Briscoe, B.R. The red palm weevil as an alien invasive: Biology and the prospects for biological control as a component of IPM. Biocontrol News Inform. 1999, 20, 35–46. [Google Scholar]
- McGuire, A.V.; Northfield, T.D. Tropical occurrence and agricultural importance of Beauveria bassiana and Metarhizium anisopliae. Front. Sustain. Food Syst. 2020, 4, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Bugeme, D.M.; Knapp, M.; Boga, H.I.; Wanjoya, A.K.; Maniania, N.K. Influence of temperature on virulence of fungal isolates of Metarhizium anisopliae and Beauveria bassiana to the two-spotted spider mite Tetranychus urticae. Mycopathologia 2009, 167, 221–227. [Google Scholar] [CrossRef]
- Seid, A.M.; Fredensborg, B.L.; Steinwender, B.M.; Meyling, N.V. Temperature-dependent germination, growth and co-infection of Beauveria spp. isolates from different climatic regions. Biocontrol Sci. Technol. 2019, 29, 411–426. [Google Scholar] [CrossRef]
- Alali, S.; Mereghetti, V.; Faoro, F.; Bocchi, S.; Al Azmeh, F.; Montagna, M. Thermotolerant isolates of Beauveria bassiana as potential control agent of insect pest in subtropical climates. PLoS ONE 2019, 14, e0211457. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
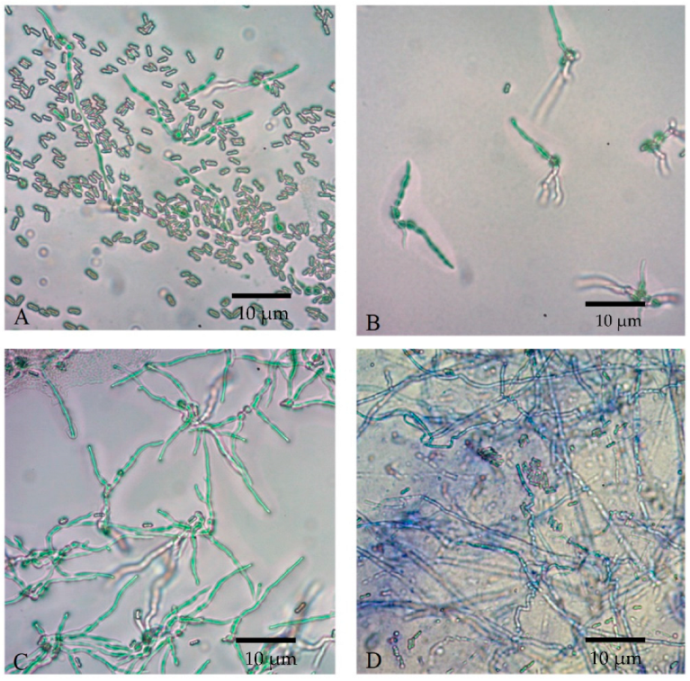{kind=link}
{kind=link}
| Oil | Emulsifier (% w/w) | Span 80: Tween 20 | Span 80: Tween 80 | HLB Number of Surfactant Blend | Required HLB of Oil |
|---|---|---|---|---|---|
| Canola oil | <20% | 8:2 | 7:3 | 6.78–7.51 | 7 |
| Capric oil | <20% | 1:1 | 1:1 | 9.65–10.5 | 11 |
| Sunflower oil | <20% | 8:2 | 7:3 | 6.78–7.51 | 7 |
| Sesame oil | <20% | 8:2 | 7:3 | 6.78–7.51 | 7 |
| Castor oil | <20% | 1:9 | 2:8 | 12.86–15.46 | 14 |
| Emulsion F25 | Storage Duration (Days) | Z-Average (d.nm ± SD) | Polydispersity Index (pdi ± sd) | Zeta Potential (mv ± sd) | Surface Tension (mN/m ± SD) |
|---|---|---|---|---|---|
| Blank | 0 | 154.3 ± 1.4 | 0.257 ± 0.012 | 36.6 ± 0.6 | 29.83 ± 0.24 |
| 30 | 213.2 ± 4.5 | 0.297 ± 0.000 | 36.6 ± 0.5 | ||
| 60 | 162.2 ± 3.4 | 0.272 ± 0.000 | 36.8 ± 1.2 | ||
| 90 | 236.4 ± 1.3 | 0.281 ± 0.017 | 36.2 ± 1.4 | ||
| Conidia-loaded | 0 | 220.9 ± 2.2 | 0.348 ± 0.000 | 37.2 ± 0.8 | 30.72 ± 0.11 |
| 30 | 805.2 ± 135.4 | 0.873 ± 0.016 | 15.3 ± 1.7 | ||
| 60 | 804.5 ± 49.4 | 0.759 ± 0.002 | 16.0 ± 0.7 | ||
| 90 | 803.3 ± 48.4 | 0.800 ± 0.040 | 15.5 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, C.J.; Halim, N.A.; Asib, N.; Zakaria, A.; Azmi, W.A. Conidial Emulsion Formulation and Thermal Storability of Metarhizium anisopliae against Red Palm Weevil, Rhynchophorusferrugineus Olivier (Coleoptera: Dryophthoridae). Microorganisms 2022, 10, 1460. https://doi.org/10.3390/microorganisms10071460
Lei CJ, Halim NA, Asib N, Zakaria A, Azmi WA. Conidial Emulsion Formulation and Thermal Storability of Metarhizium anisopliae against Red Palm Weevil, Rhynchophorusferrugineus Olivier (Coleoptera: Dryophthoridae). Microorganisms. 2022; 10(7):1460. https://doi.org/10.3390/microorganisms10071460
Chicago/Turabian StyleLei, Cheong Jia, Najihah Abdul Halim, Norhayu Asib, Azlina Zakaria, and Wahizatul Afzan Azmi. 2022. "Conidial Emulsion Formulation and Thermal Storability of Metarhizium anisopliae against Red Palm Weevil, Rhynchophorusferrugineus Olivier (Coleoptera: Dryophthoridae)" Microorganisms 10, no. 7: 1460. https://doi.org/10.3390/microorganisms10071460