
Soil pH, Calcium Content and Bacteria as Major Factors Responsible for the Distribution of the Known Fraction of the DNA Bacteriophage Populations in Soils of Luxembourg

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
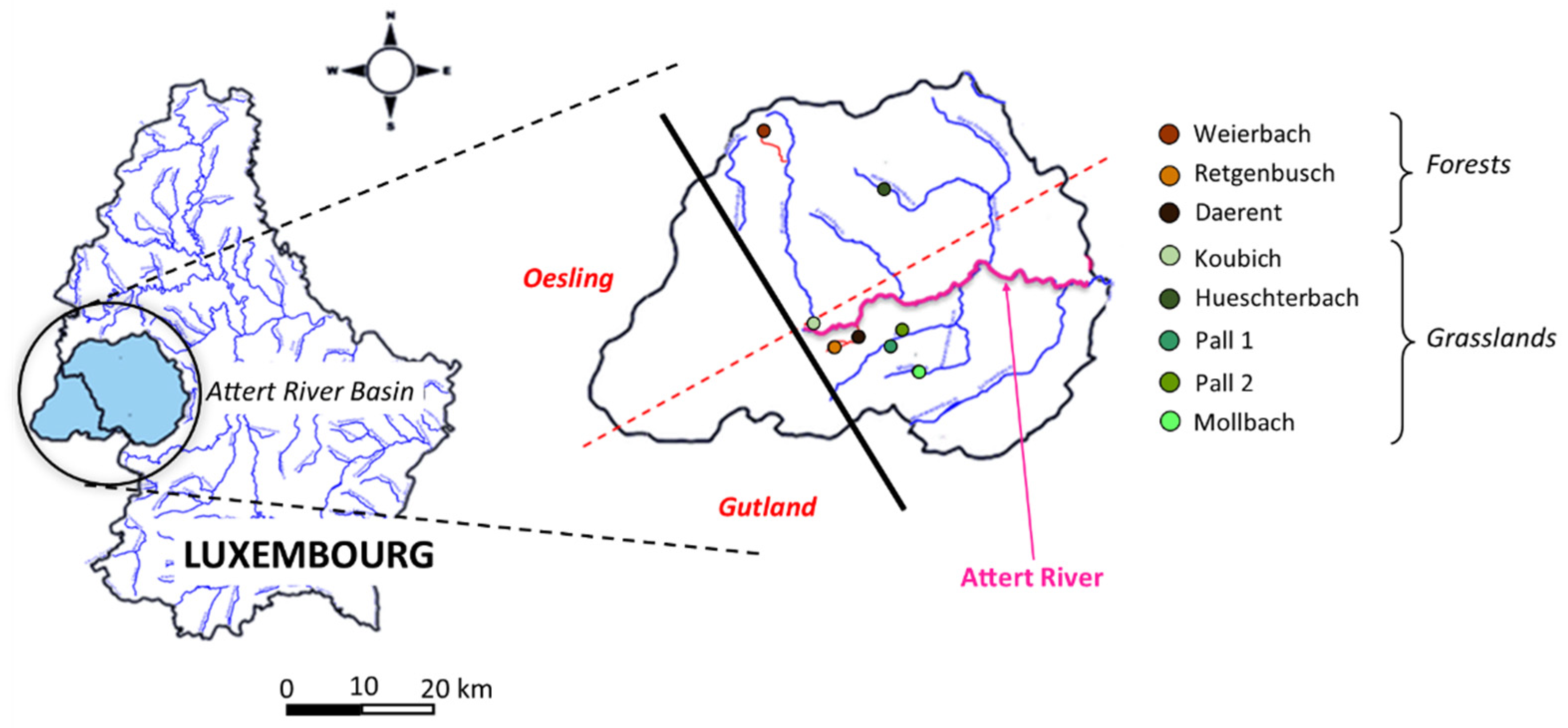
2.1. Studied Area
2.2. Sample Collection
2.3. Soil Characterisation
2.4. Viral Quantification
2.4.1. Viral Elution and Concentration
2.4.2. Viral-like Particles (VLPs) Counting
2.5. Identification of the Microbial Populations
2.5.1. Total DNA Extraction and Shotgun Metagenomics Sequencing
2.5.2. Reference-Based Bioinformatics
2.6. Statistical Analyses
3. Results
3.1. Soil Description
3.2. Description of the DNA-Bacteriophage Populations
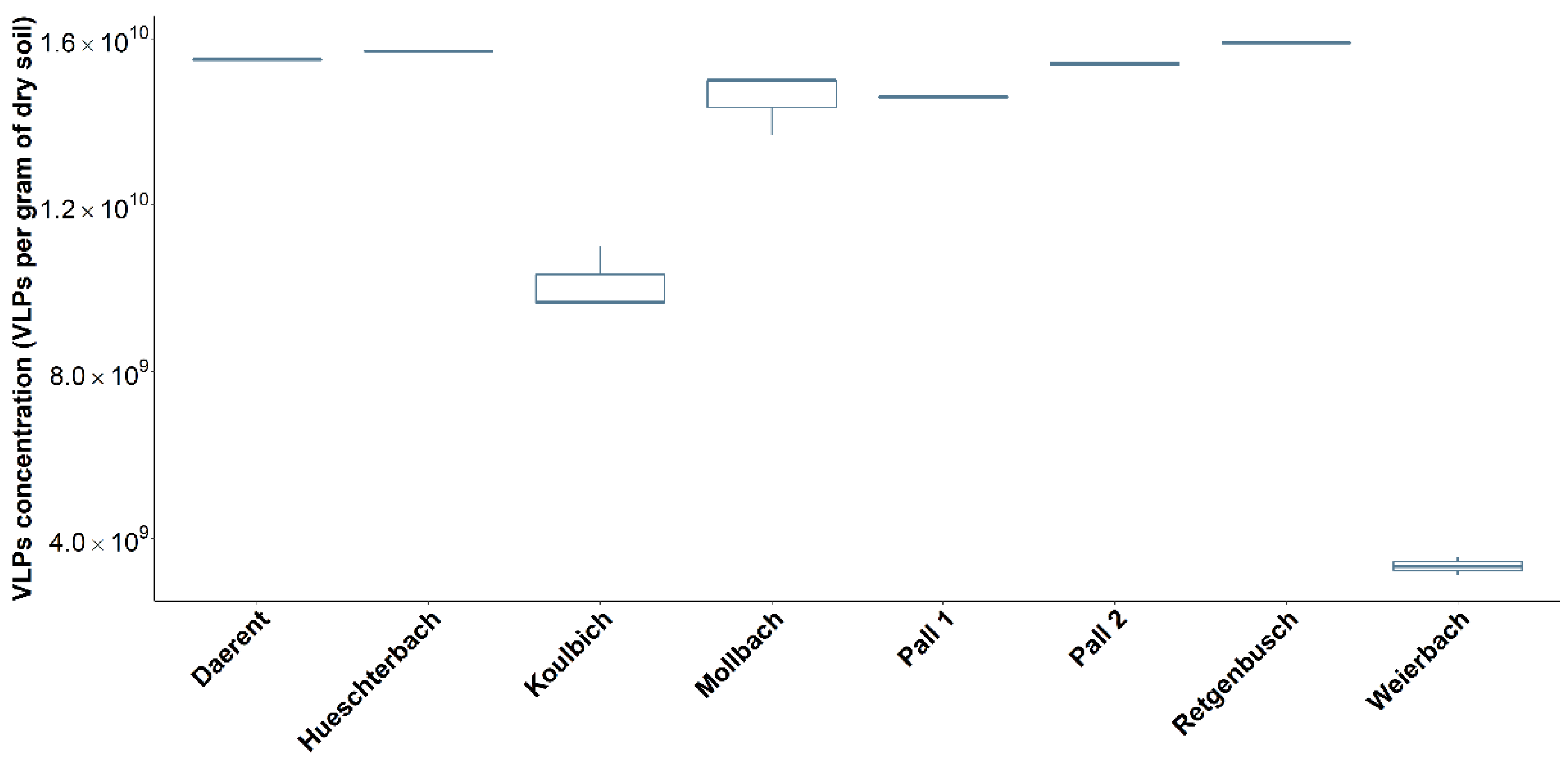
3.2.1. VLPs Enumeration
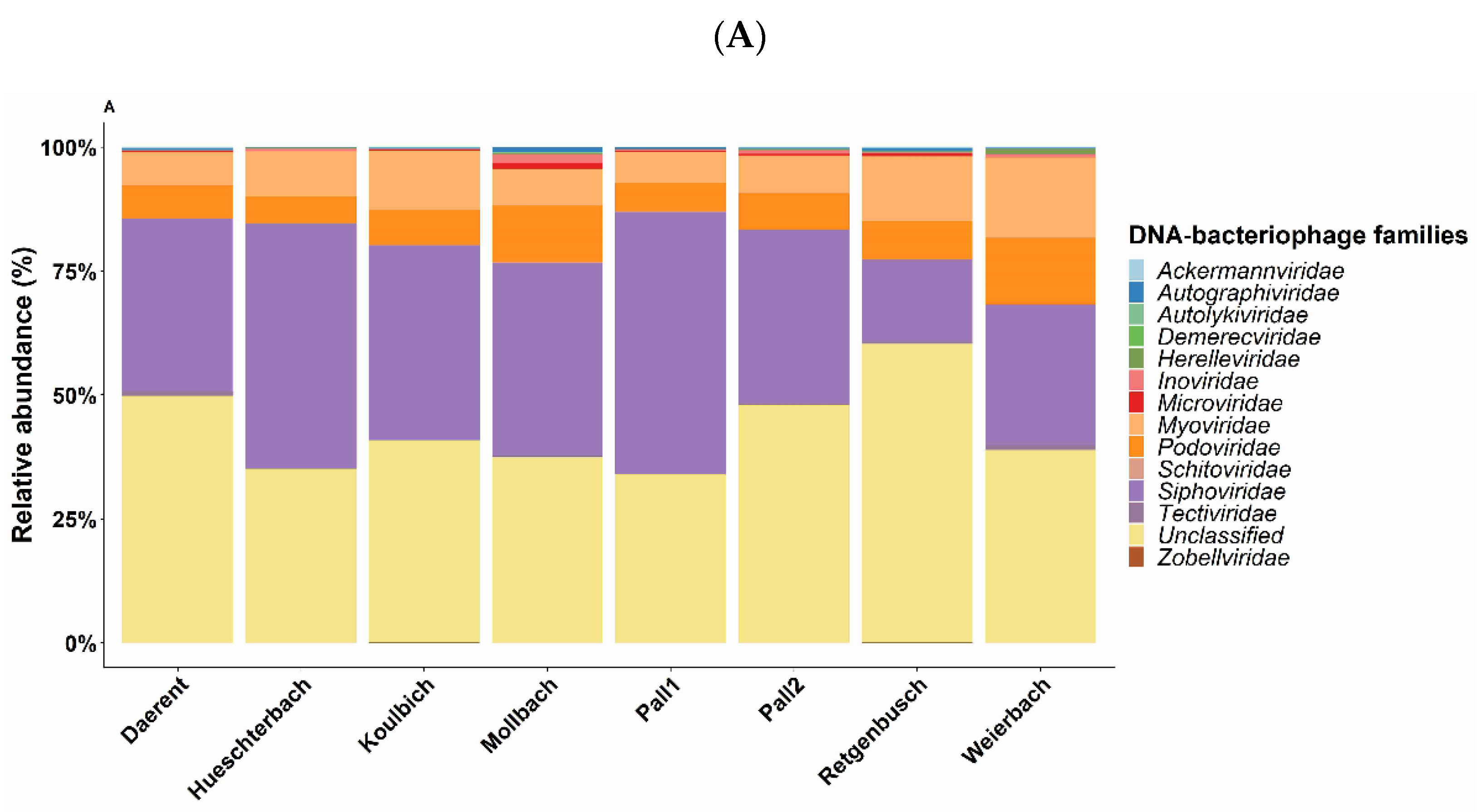
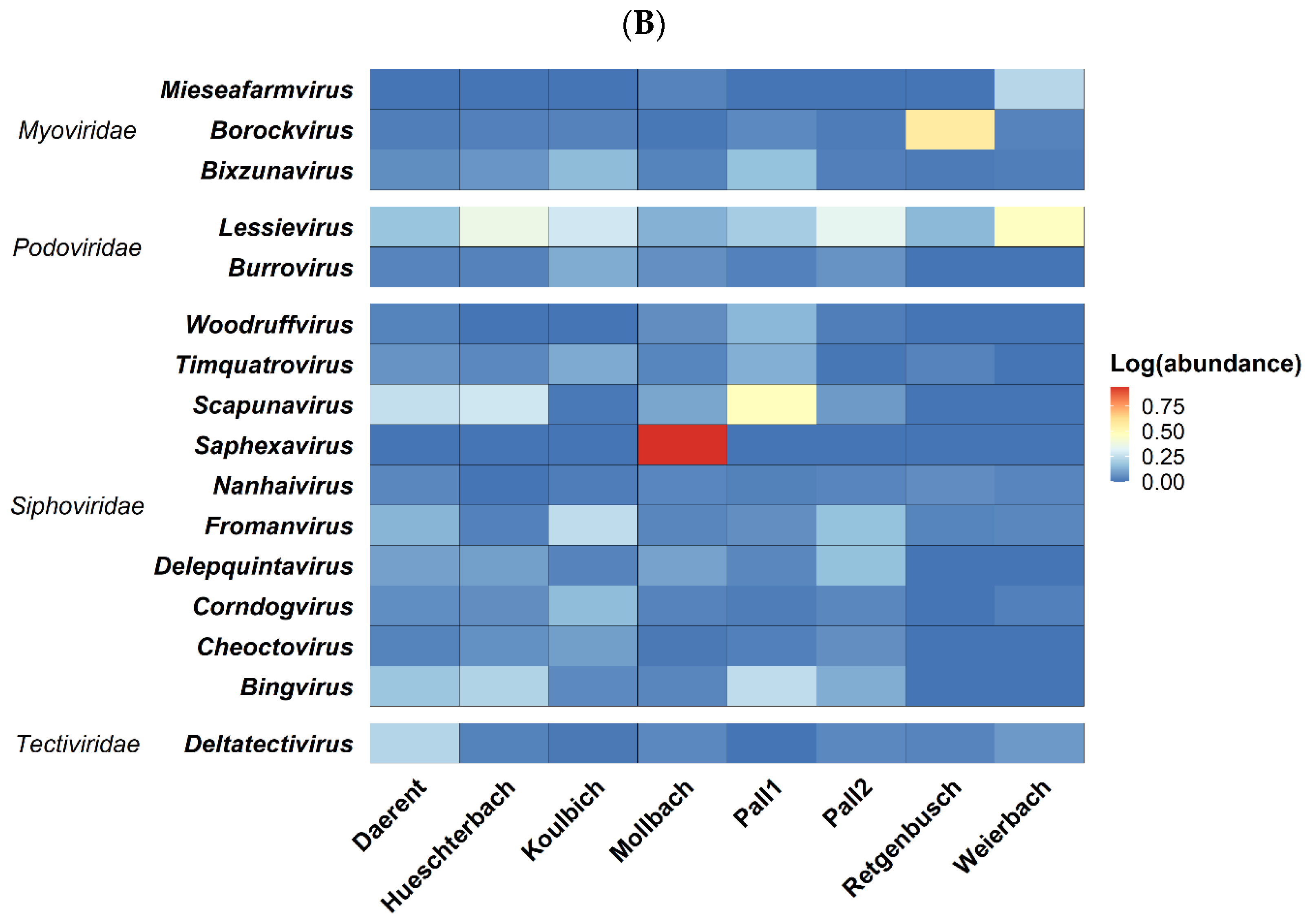
3.2.2. Taxonomic Classification
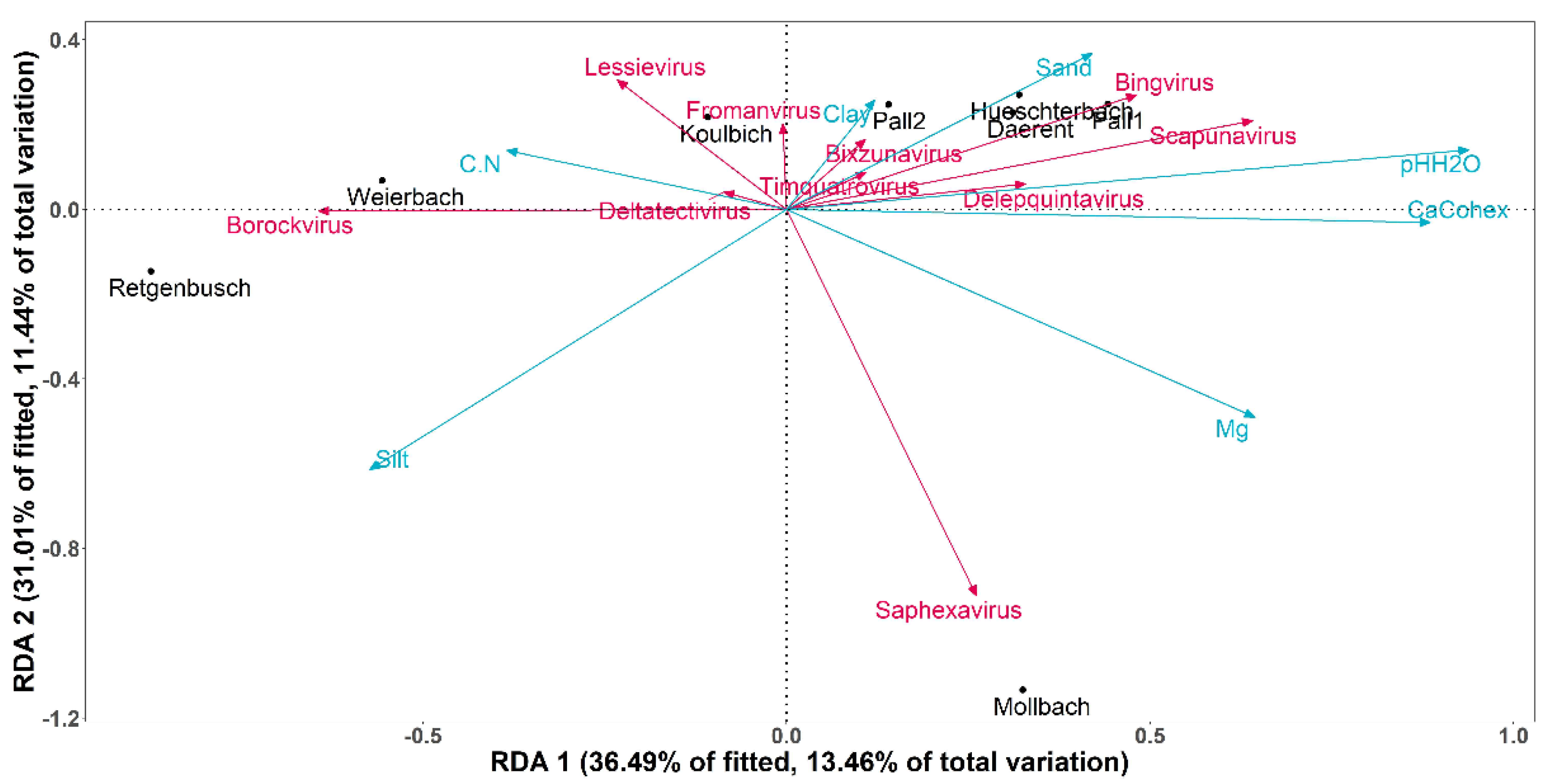
3.3. Impact of the Soil Properties on Known DNA-Bacteriophage Distribution
3.3.1. α-Diversity: Known DNA-Bacteriophage Diversity within Sites
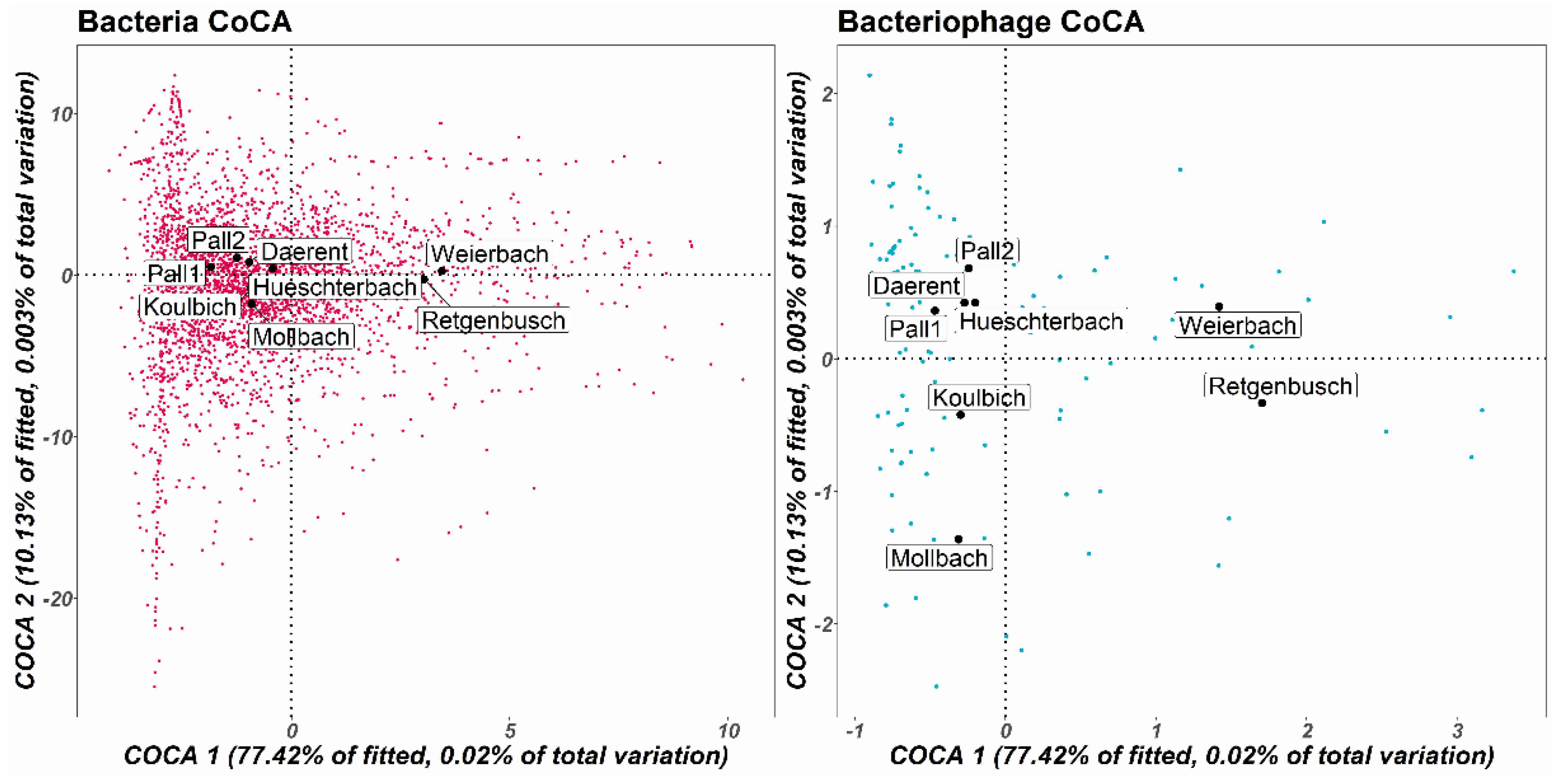
3.3.2. β-Diversity: Known DNA-Bacteriophage Diversity between Sites
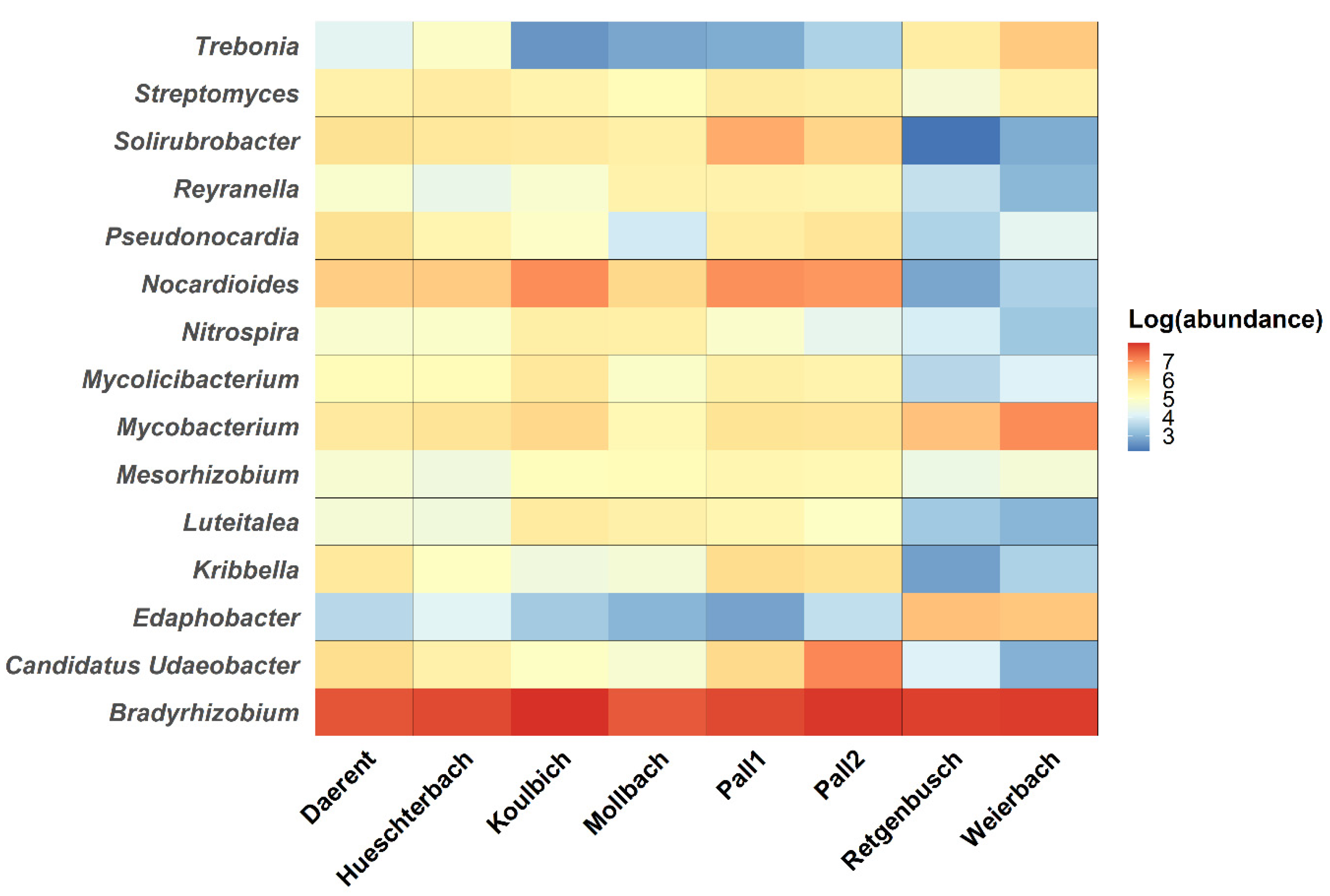
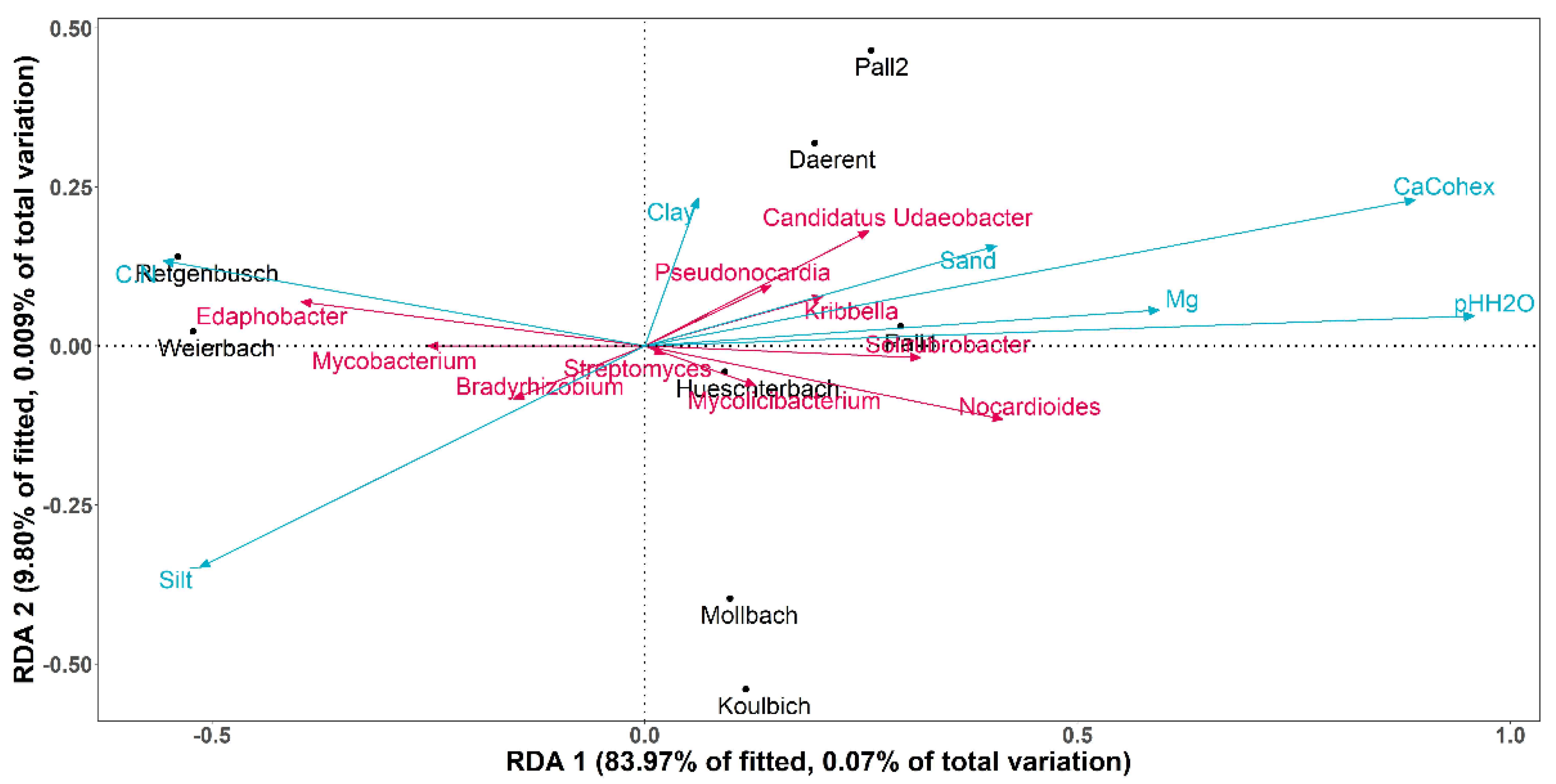
3.4. Known Bacterial Populations and Connections with the Known DNA-Bacteriophage Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant Diversity Increases Soil Microbial Activity and Soil Carbon Storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef]
- Vezzani, F.M.; Anderson, C.; Meenken, E.; Gillespie, R.; Peterson, M.; Beare, M.H. The Importance of Plants to Development and Maintenance of Soil Structure, Microbial Communities and Ecosystem Functions. Soil Tillage Res. 2018, 175, 139–149. [Google Scholar] [CrossRef]
- Eldridge, D.; James, A. Soil-disturbance by Native Animals Plays a Critical Role in Maintaining Healthy Australian Landscapes. Ecol. Manag. Restor. 2009, 10, S27–S34. [Google Scholar] [CrossRef]
- Lou, X.; Zhao, J.; Lou, X.; Xia, X.; Feng, Y.; Li, H. The Biodegradation of Soil Organic Matter in Soil-Dwelling Humivorous Fauna. Front. Bioeng. Biotechnol. 2021, 9, 1406. [Google Scholar] [CrossRef] [PubMed]
- Odelade, K.A.; Babalola, O.O. Bacteria, Fungi and Archaea Domains in Rhizospheric Soil and Their Effects in Enhancing Agricultural Productivity. Int. J. Environ. Res. Public Health 2019, 16, 3873. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and Fungi Can Contribute to Nutrients Bioavailability and Aggregate Formation in Degraded Soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Buerkert, A.; Joergensen, R.G.; Ludwig, B.; Schlecht, E. Chapter 18—Nutrient and Carbon Fluxes in Terrestrial Agro-Ecosystems. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012; pp. 473–482. ISBN 978-0-12-384905-2. [Google Scholar]
- Ortiz, A.; Sansinenea, E. The Role of Beneficial Microorganisms in Soil Quality and Plant Health. Sustainability 2022, 14, 5358. [Google Scholar] [CrossRef]
- Beule, L.; Vaupel, A.; Moran-Rodas, V.E. Abundance, Diversity, and Function of Soil Microorganisms in Temperate Alley-Cropping Agroforestry Systems: A Review. Microorganisms 2022, 10, 616. [Google Scholar] [CrossRef]
- Armon, R. Soil Bacteria and Bacteriophages. In Biocommunication in Soil Microorganisms; Witzany, G., Ed.; Springer: Heidelberg, Germany, 2011; Volume 23, pp. 67–112. ISBN 978-3-642-14511-7. [Google Scholar]
- Kimura, M.; Jia, Z.J.; Nakayama, N.; Asakawa, S. Ecology of Viruses in Soils: Past, Present and Future Perspectives. Soil Sci. Plant Nutr. 2008, 54, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Coyne, M.S. A Cartoon History of Soil Microbiology. J. Nat. Resour. Life Sci. Educ. 1996, 25, 30–36. [Google Scholar] [CrossRef]
- Williamson, K.E.; Fuhrmann, J.J.; Wommack, K.E.; Radosevich, M. Viruses in Soil Ecosystems: An Unknown Quantity Within an Unexplored Territory. Annu. Rev. Virol. 2017, 4, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.E.; Corzo, K.A.; Drissi, C.L.; Buckingham, J.M.; Thompson, C.P.; Helton, R.R. Estimates of Viral Abundance in Soils Are Strongly Influenced by Extraction and Enumeration Methods. Biol. Fertil. Soils 2013, 49, 857–869. [Google Scholar] [CrossRef]
- Bertrand, I.; Schijven, J.F.; Sánchez, G.; Wyn-Jones, P.; Ottoson, J.; Morin, T.; Muscillo, M.; Verani, M.; Nasser, A.; de Roda Husman, A.M.; et al. The Impact of Temperature on the Inactivation of Enteric Viruses in Food and Water: A Review. J. Appl. Microbiol. 2012, 112, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Corapcioglu, M.Y.; Vogel, J.R.; Munster, C.L.; Pillai, S.D.; Dowd, S.; Wang, S. Virus Transport Experiments in a Sandy Aquifer. Water Air Soil Pollut. 2006, 169, 47–65. [Google Scholar] [CrossRef]
- Fauvel, B.; Cauchie, H.M.; Gantzer, C.; Ogorzaly, L. Influence of Physico-Chemical Characteristics of Sediment on the in Situ Spatial Distribution of F-Specific RNA Phages in the Riverbed. FEMS Microbiol. Ecol. 2019, 95, fiy240. [Google Scholar] [CrossRef]
- Jurczak-Kurek, A.; Gasior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of Bacteriophages: Morphological and Biological Properties of a Large Group of Phages Isolated from Urban Sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Ong, S.L.; Hu, J.Y.; Tan, X.L.; Ng, W.J. Effects of PH and Temperature on the Survival of Coliphages MS2 and Qβ. J. Ind. Microbiol. Biotechnol. 2003, 30, 549–552. [Google Scholar] [CrossRef]
- Nishide, M.; Tsujimoto, K.; Uozaki, M.; Ikeda, K.; Yamasaki, H.; Koyama, A.H.; Arakawa, T. Effects of Electrolytes on Virus Inactivation by Acidic Solutions. Int. J. Mol. Med. 2011, 27, 803–809. [Google Scholar] [CrossRef]
- Taj, M.K.; Ling, J.X.; Bing, L.L.; Qi, Z.; Taj, I.; Hassani, T.M.; Samreen, Z.; Yunlin, W. Effect of Dilution, Temperature and PH on the Lysis Activity of T4 Phage against E. Coli BL21. J. Anim. plant Sci. 2014, 24, 1252–1255. [Google Scholar]
- Cuadros, J. Clay Minerals Interaction with Microorganisms: A Review. Clay Miner. 2017, 52, 235–261. [Google Scholar] [CrossRef] [Green Version]
- Park, J.A.; Kang, J.K.; Kim, J.H.; Kim, S.B.; Yu, S.; Kim, T.H. Bacteriophage Removal in Various Clay Minerals and Clay-Amended Soils. Environ. Eng. Res. 2015, 20, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.W.; Ryan, J.N. Use of PRD1 Bacteriophage in Groundwater Viral Transport, Inactivation, and Attachment Studies. FEMS Microbiol. Ecol. 2004, 49, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.E.; Radosevich, M.; Wommack, K.E. Abundance and Diversity of Viruses in Six Delaware Soils. Appl. Environ. Microbiol. 2005, 71, 3119–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, M.; Prigent, M.; Dutertre, M.; Confalonieri, F.; Dubow, M. Bacteriophage Morphotype and Genome Diversity in Seine River Sediment. Freshw. Biol. 2008, 53, 1176–1185. [Google Scholar] [CrossRef]
- Parmar, K.; Dafale, N.; Pal, R.; Tikariha, H.; Purohit, H. An Insight into Phage Diversity at Environmental Habitats Using Comparative Metagenomics Approach. Curr. Microbiol. 2018, 75, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.; Schellhorn, H.E. Spatial and Temporal Dynamics of Virus Occurrence in Two Freshwater Lakes Captured through Metagenomic Analysis. Front. Microbiol. 2015, 6, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.; Liang, Y.; Wang, M.; Jiang, Y.; Yang, Q.; Xia, J.; Zhou, X.; You, S.; Gao, C.; Wang, J.; et al. Viral Diversity and Its Relationship With Environmental Factors at the Surface and Deep Sea of Prydz Bay, Antarctica. Front. Microbiol. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Narr, A.; Nawaz, A.; Wick, L.Y.; Harms, H.; Chatzinotas, A. Soil Viral Communities Vary Temporally and along a Land Use Transect as Revealed by Virus-like Particle Counting and a Modified Community Fingerprinting Approach (FRAPD). Front. Microbiol. 2017, 8, 1975. [Google Scholar] [CrossRef]
- Trubl, G.; Jang, H.B.; Roux, S.; Emerson, J.B.; Solonenko, N.; Vik, D.R.; Solden, L.; Ellenbogen, J.; Runyon, A.T.; Bolduc, B.; et al. Soil Viruses Are Underexplored Players in Ecosystem Carbon Processing. mSystems 2018, 3, e00076-18. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.-M.; Liu, S.-Y.; Bi, L.; Chen, S.-J.; Wu, H.-Y.; Ge, Y.; Han, B.; Zhang, L.-M.; He, J.-Z.; Han, L.-L. Distribution Characteristics of Soil Viruses Under Different Precipitation Gradients on the Qinghai-Tibet Plateau. Front. Microbiol. 2022, 13, 848305. [Google Scholar] [CrossRef]
- Han, L.L.; Yu, D.T.; Bi, L.; Du, S.; Silveira, C.; Cobián Güemes, A.G.; Zhang, L.M.; He, J.Z.; Rohwer, F. Distribution of Soil Viruses across China and Their Potential Role in Phosphorous Metabolism. Environ. Microbiome 2022, 17, 6. [Google Scholar] [CrossRef]
- Koskella, B.; Brockhurst, M.A. Bacteria–Phage Coevolution as a Driver of Ecological and Evolutionary Processes in Microbial Communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Fazzino, L.; Anisman, J.; Chacón, J.M.; Heineman, R.H.; Harcombe, W.R. Lytic Bacteriophage Have Diverse Indirect Effects in a Synthetic Cross-Feeding Community. ISME J. 2020, 14, 123–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.B.; Weitz, J.S.; Wilhelm, S. Viral Ecology Comes of Age. Environ. Microbiol. Rep. 2017, 9, 33–35. [Google Scholar] [CrossRef] [PubMed]
- Klimenko, A.I.; Matushkin, Y.G.; Kolchanov, N.A.; Lashin, S.A. Bacteriophages Affect Evolution of Bacterial Communities in Spatially Distributed Habitats: A Simulation Study. BMC Microbiol. 2016, 16, S10. [Google Scholar] [CrossRef] [Green Version]
- Ashelford, K.E.; Day, M.J.; Fry, J.C. Elevated Abundance of Bacteriophage Infecting Bacteria in Soil. Appl. Environ. Microbiol. 2003, 69, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, L.P.P.; Spor, A.; Kot, W.; Breuil, M.C.; Hansen, L.H.; Setubal, J.C.; Philippot, L. Impact of Phages on Soil Bacterial Communities and Nitrogen Availability under Different Assembly Scenarios. Microbiome 2020, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Marx, S.; Flammang, F. La Cartographie Des Sols Au Grand-Duché de Luxembourg Version Provisoire_V4; Ministère de L’agriculture, de la Viticulture et du Développement Rural: Luxembourg, 2015. [Google Scholar]
- Heffernan, A.L.; Aylward, L.L.; Toms, L.M.L.; Sly, P.D.; Macleod, M.; Mueller, J.F. Pooled Biological Specimens for Human Biomonitoring of Environmental Chemicals: Opportunities and Limitations. J. Expo. Sci. Environ. Epidemiol. 2014, 24, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Caudill, S.P. Characterizing Populations of Individuals Using Pooled Samples. J. Expo. Sci. Environ. Epidemiol. 2010, 20, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Roy, K.; Ghosh, D.; DeBruyn, J.M.; Dasgupta, T.; Wommack, K.E.; Liang, X.; Wagner, R.E.; Radosevich, M. Temporal Dynamics of Soil Virus and Bacterial Populations in Agricultural and Early Plant Successional Soils. Front. Microbiol. 2020, 11, 1494. [Google Scholar] [CrossRef]
- AFNOR. ISO 10390: Sols, Biodéchets Traités et Boues—Détermination Du PH, 3rd ed.; Association Française de Normalisation: Geneva, Switzerland, 2021. [Google Scholar]
- VDLUFA. Methodenbuch A5.1.1: Bestimmung des PH-Wertes; Verlag: Darmstadt, Germany, 2016. [Google Scholar]
- VDLUFA. Methodenbuch A6.2.1.1: Bestimmung von Phosphor und Kalium im Calcium-Acetat-Lactat-Auszug; Verlag: Darmstadt Germany, 2012. [Google Scholar]
- VDLUFA. Methodenbuch A6.2.1.7: Bestimmung von Pflanzenverfügbarem Kalium und Natrium im Calciumchloridauszug auf Gewichtsbasis; Verlag: Darmstadt, Germany, 1997. [Google Scholar]
- AFNOR. ISO 10694: Qualité du Sol–Dosage du Carbone Organique et du Carbon Total Après Combustion Sèche (Analyse Élémentaire), 1st ed.; Association Française de Normalisation: Geneva, Switzerland, 1995. [Google Scholar]
- AFNOR. ISO 13878: Qualité du Sol–Détermination de la Teneur Totale en Azote par Combustion Sèche (“Analyse Élémentaire”), 1st ed.; Association Française de Normalisation: Geneva, Switzerland, 1998. [Google Scholar]
- AFNOR. ISO 23470: Qualité Du Sol–Détermination de La Capacité D’échange Cationique (CEC) Effective et des Cations Échangeables à L’aide d’une Solution de Trichlorure de Cobaltihexammine, 1st ed.; Association Française de Normalisation: Geneva, Switzerland, 2007. [Google Scholar]
- Marx, S.; Flammang, F. Cartographie des Sols au Grand-Duché de Luxembourg; Ministère de L’agriculture, de la Viticulture et du Développement Rural: Luxembourg, 2018. [Google Scholar]
- Jacquet, S.; Dorigo, U.; Personnic, S. A Few Tests Prior to Flow Cytometry and Epifluorescence Analyses of Freshwater Bacterioand Virioplankton Communities. Flow Cytom. Princ. Methodol. Appl. 2013, 1, 1–30. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. MetaSPAdes: A New Versatile Metagenomic Assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinegger, M.; Söding, J. MMseqs2 Enables Sensitive Protein Sequence Searching for the Analysis of Massive Data Sets. Nat. Biotechnol. 2017, 35, 1026–1028. [Google Scholar] [CrossRef] [Green Version]
- Mirdita, M.; Steinegger, M.; Breitwieser, F.; Sö Ding, J.; Levy Karin, E. Fast and Sensitive Taxonomic Assignment to Metagenomic Contigs. Bioinformatics 2021, 37, 3029–3031. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Palermo, C.N.; Fulthorpe, R.R.; Saati, R.; Short, S.M. Metagenomic Analysis of Virus Diversity and Relative Abundance in a Eutrophic Freshwater Harbour. Viruses 2019, 11, 792. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, M.C.; Konaté, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts? A Comparative Study of Quantification Measures for the Analysis of RNA-Seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. RStudio; PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 11 July 2022).
- Thukral, A.K. A Review on Measurement of Alpha Diversity in Biology. Agric. Res. J. 2017, 54, 10. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 11 July 2022).
- Mulder, C.P.H.; Bazeley-White, E.; Dimitrakopoulos, P.G.; Hector, A.; Scherer-Lorenzen, M.; Schmid, B. Species Evenness and Productivity in Experimental Plant Communities. Oikos 2004, 107, 50–63. [Google Scholar] [CrossRef]
- Moore, J.C. Diversity, Taxonomic versus Functional. In Encyclopedia of Biodiversity, 2nd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 648–656. ISBN 9780123847195. [Google Scholar]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA (United States Department of Agriculture, Natural resources conservation service, Soil Texture Calculator. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/survey/?cid=nrcs142p2_054167 (accessed on 18 July 2022).
- Pratama, A.A.; van Elsas, J.D. The ‘Neglected’ Soil Virome—Potential Role and Impact. Trends Microbiol. 2018, 26, 649–662. [Google Scholar] [CrossRef]
- Williamson, K.E.; Wommack, K.E.; Radosevich, M. Sampling Natural Viral Communities from Soil for Culture-Independent Analyses. Appl. Environ. Microbiol. 2003, 69, 6628–6633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duhamel, S.; Jacquet, S. Flow Cytometric Analysis of Bacteria- and Virus-like Particles in Lake Sediments. J. Microbiol. Methods 2006, 64, 316–332. [Google Scholar] [CrossRef] [Green Version]
- Colombet, J.; Billard, H.; Viguès, B.; Balor, S.; Boulé, C.; Geay, L.; Benzerara, K.; Menguy, N.; Ilango, G.; Fuster, M.; et al. Discovery of High Abundances of Aster-Like Nanoparticles in Pelagic Environments: Characterization and Dynamics. Front. Microbiol. 2019, 10, 2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Kim, K.H.; Chang, H.W.; Nam, Y.D.; Roh, S.W.; Kim, M.S.; Sung, Y.; Jeon, C.O.; Oh, H.M.; Bae, J.W. Amplification of Uncultured Single-Stranded DNA Viruses from Rice Paddy Soil. Appl. Environ. Microbiol. 2008, 74, 5975–5985. [Google Scholar] [CrossRef] [Green Version]
- Reavy, B.; Swanson, M.M.; Cock, P.J.A.; Dawson, L.; Freitag, T.E.; Singh, B.K.; Torrance, L.; Mushegian, A.R.; Taliansky, M. Distinct Circular Single-Stranded DNA Viruses Exist in Different Soil Types. Appl. Environ. Microbiol. 2015, 81, 3934–3945. [Google Scholar] [CrossRef] [Green Version]
- Grujcic, V.; Nuy, J.K.; Salcher, M.M.; Shabarova, T.; Kasalicky, V.; Boenigk, J.; Jensen, M.; Simek, K. Cryptophyta as Major Bacterivores in Freshwater Summer Plankton. ISME J. 2018, 12, 1668–1681. [Google Scholar] [CrossRef] [Green Version]
- Cherwa, J.E.; Fane, B.A. Microviridae: Microviruses and Gokushoviruses. eLS 2011, 5, 1–10. [Google Scholar] [CrossRef]
- Székely, A.J.; Breitbart, M. Single-Stranded DNA Phages: From Early Molecular Biology Tools to Recent Revolutions in Environmental Microbiology. FEMS Microbiol. Lett. 2016, 363, 27. [Google Scholar] [CrossRef]
- Wuddivira, M.N.; Camps-Roach, G. Effects of Organic Matter and Calcium on Soil Structural Stability. Eur. J. Soil Sci. 2007, 58, 722–727. [Google Scholar] [CrossRef]
- Zavarzin, G.A. Microbial Geochemical Calcium Cycle. Microbiology 2002, 71, 1–17. [Google Scholar] [CrossRef]
- Whittinghill, K.A.; Hobbie, S.E. Effects of PH and Calcium on Soil Organic Matter Dynamics in Alaskan Tundra. Biogeochemistry 2011, 111, 569–581. [Google Scholar] [CrossRef]
- Ng, J.F.; Ahmed, O.H.; Jalloh, M.B.; Omar, L.; Kwan, Y.M.; Musah, A.A.; Poong, K.H. Soil Nutrient Retention and PH Buffering Capacity Are Enhanced by Calciprill and Sodium Silicate. Agronomy 2022, 12, 219. [Google Scholar] [CrossRef]
- Pham, M.; Mintz, E.A.; Nguyen, T.H. Deposition Kinetics of Bacteriophage MS2 to Natural Organic Matter: Role of Divalent Cations. J. Colloid Interface Sci. 2009, 338, 1–9. [Google Scholar] [CrossRef]
- Brydon, J.E.; Sowden, F.J. A Study o the Clay-Humus Complexes of a Chernozemic and a Podzol Soil. J. Soil Sci. 2011, 39, 136–143. [Google Scholar] [CrossRef]
- Xu, X.; Li, Y.; Hu, X.; Xie, G.; Xu, H.; Gao, M.; Zhang, X.; Zhang, R.; Tang, C.; Hu, X. Effect of Humic Acid on the Adsorption/Desorption Behaviors of Trivalent Chromium on Calcium Modified Montmorillonite and Kaolinite. ChemistrySelect 2022, 7, e202104302. [Google Scholar] [CrossRef]
- Sadeghi, G.; Behrends, T.; Schijven, J.F.; Hassanizadeh, S.M. Effect of Dissolved Calcium on the Removal of Bacteriophage PRD1 during Soil Passage: The Role of Double-Layer Interactions. J. Contam. Hydrol. 2013, 144, 78–87. [Google Scholar] [CrossRef]
- McLeod, M.; Aislabie, J.; Smith, J.; Fraser, R.; Roberts, A.; Taylor, M. Viral and Chemical Tracer Movement through Contrasting Soils. J. Environ. Qual. 2001, 30, 2134–2140. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cassi, X.; Rusinol, M.; Martinez-Puchol, S. Viral Concentration and Amplification from Human Serum Samples Prior to Application of Next-Generation Sequencing Analysis. In The Human Virome: Methods and Protocols; Springer: New York, NY, USA, 2018; Volume 1838, pp. 173–188. ISBN 9781493986828. [Google Scholar]
- Zhang, Y.Z.; Shi, M.; Holmes, E.C. Using Metagenomics to Characterize an Expanding Virosphere. Cell 2018, 172, 1168–1172. [Google Scholar] [CrossRef]
- Trubl, G.; Hyman, P.; Roux, S.; Abedon, S.T. Coming-of-Age Characterization of Soil Viruses: A User’s Guide to Virus Isolation, Detection within Metagenomes, and Viromics. Soil Syst. 2020, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Baer, A.; Kehn-Hall, K. Viral Concentration Determination Through Plaque Assays: Using Traditional and Novel Overlay Systems. J. Vis. Exp. 2014, 93, e52065. [Google Scholar] [CrossRef] [PubMed]
- Van Twest, R.; Kropinski, A.M. Bacteriophages Enrichment from Water and Soil; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 501, ISBN 1402037902. [Google Scholar]
- Walshe, G.E.; Pang, L.; Flury, M.; Close, M.E.; Flintoft, M. Effects of PH, Ionic Strength, Dissolved Organic Matter, and Flow Rate on the Co-Transport of MS2 Bacteriophages with Kaolinite in Gravel Aquifer Media. Water Res. 2010, 44, 1255–1269. [Google Scholar] [CrossRef]
- Furiga, A.; Pierre, G.; Glories, M.; Aimar, P.; Roques, C.; Causserand, C.; Berge, M. Effects of Ionic Strength on Bacteriophage MS2 Behavior and Their Implications for the Assessment of Virus Retention by Ultrafiltration Membranes. Appl. Environ. Microbiol. 2011, 77, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Breitbart, M.; Nulton, J.; Salamon, P.; Lozupone, C.; Jones, R.; Robeson, M.; Edwards, R.A.; Felts, B.; Rayhawk, S.; et al. Metagenomic and Small-Subunit RRNA Analyses Reveal the Genetic Diversity of Bacteria, Archaea, Fungi, and Viruses in Soil. Appl. Environ. Microbiol. 2007, 73, 7059–7066. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.P.; Warnow, T.; Pop, M.; White, B. A Perspective on 16S RRNA Operational Taxonomic Unit Clustering Using Sequence Similarity. NPJ Biofilms Microbiomes 2016, 2, 16004. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L. Assessing and Improving Methods Used in Operational Taxonomic Unit-Based Approaches for 16S RRNA Gene Sequence Analysis. Appl. Environ. Microbiol. 2011, 77, 3219–3226. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Xun, W.; Sun, L.; Zhang, N.; Shen, Q.; Zhang, R. Effect of Different Long-Term Fertilization Regimes on the Viral Community in an Agricultural Soil of Southern China. Eur. J. Soil Biol. 2014, 62, 121–126. [Google Scholar] [CrossRef]
- Doan, T.T.; Bouvier, C.; Bettarel, Y.; Bouvier, T.; Henry-des-Tureaux, T.; Janeau, J.L.; Lamballe, P.; Van Nguyen, B.; Jouquet, P. Influence of Buffalo Manure, Compost, Vermicompost and Biochar Amendments on Bacterial and Viral Communities in Soil and Adjacent Aquatic Systems. Appl. Soil Ecol. 2014, 73, 78–86. [Google Scholar] [CrossRef]
- Swanson, M.M.; Fraser, G.; Daniell, T.J.; Torrance, L.; Gregory, P.J.; Taliansky, M. Viruses in Soils: Morphological Diversity and Abundance in the Rhizosphere. Ann. Appl. Biol. 2009, 155, 51–60. [Google Scholar] [CrossRef]
- Ateba, C.N.; Akindolire, M.A. Isolation and Characterisation of Bacteriophages with Lytic Activity Against Virulent Escherichia Coli O157:H7: Potential Bio-Control Agents. Preprints 2019, 2019010132. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Saxena, H.M. Isolation and Characterization of BpL1, a Broad Acting Lytic Bacteriophage against Brucella. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2486–2496. [Google Scholar] [CrossRef]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Sites | Sample ID | Vegetation | GPS Coordinates |
|---|---|---|---|
| Weierbach | WPF12092019 | Forest | (N) 49.831937 (E) 5.80341 |
| Retgenbusch | RPF12092019 | Forest | (N) 49.743436 (E) 5.834341 |
| Daerent | DPF12092019 | Forest | (N) 49.748226 (E) 5.856064 |
| Koulbich | KPF12092019 | Grassland | (N) 49.751942 (E) 5.826879 |
| Hueschterbach | HPF12092019 | Grassland | (N) 49.809914 (E) 5.863031 |
| Pall 1 | P1PF12092019 | Grassland | (N) 49.74617 (E) 5.871577 |
| Pall 2 | P2PF12092019 | Grassland | (N) 49.748935 (E) 5.881184 |
| Mollbach | MPF12092019 | Grassland | (N) 49.7347405 (E) 5.888961 |
| Site Names | GWC | pH | C/N | Corg | Ntot | Mg | Na | P2O5 | K2O | CEC | Ca2+ | S/T | Textural Class | Clay | Silt | Sand |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | - | - | % | % | mg per 100 g ds | mg per 100 g ds | mg per 100 g ds | mg per 100 g ds | Meq kg−1 | Meq kg−1 | % | % | % | % | ||
| Weierbach | 24.6 | 4.1 | 18 | 9.7 | 0.54 | 3 | 2 | 5 | 13 | 85 | 21 | 42 | CL | 23.9 | 42.2 | 33.9 |
| Retgenbusch | 13.7 | 4.5 | 8.6 | 1.9 | 0.22 | 3 | 1 | 3 | 9 | 26 | 16 | 101 | A | 27.2 | 58.5 | 14.3 |
| Daerent | 15.8 | 6.3 | 10 | 3.9 | 0.39 | 46 | 2 | 4 | 21 | 171 | 126 | 107 | SL | 53.9 | 28.0 | 18.1 |
| Koulbich | 10.8 | 5.7 | 8.3 | 1.9 | 0.23 | 12 | 3 | 9 | 11 | 74 | 58 | 105 | L | 37.7 | 43.6 | 18.6 |
| Hueschterbach | 14.7 | 6.4 | 8.8 | 2.2 | 0.25 | 11 | 3 | 20 | 16 | 85 | 87 | 126 | L | 52.0 | 29.6 | 18.4 |
| Pall 1 | 20.9 | 6.8 | 9.8 | 5.1 | 0.52 | 17 | 2 | 6 | 17 | 237 | 166 | 100 | CL | 24.2 | 42.6 | 33.2 |
| Pall 2 | 16.3 | 6.2 | 8.8 | 3.6 | 0.41 | 31 | 3 | 7 | 15 | 172 | 151 | 115 | CL | 33.9 | 38.1 | 28.1 |
| Mollbach | 18.3 | 6.0 | 8.3 | 3.1 | 0.23 | 52 | 5 | 13 | 21 | 143 | 131 | 112 | SL | 28.1 | 52.5 | 19.4 |
| Sites | Number of Read Pairs | Yield in Mbp | Average Quality (Q-Score) | Number of Total Mapped Read Pairs | Percentage of Mapped Read Pairs | Number of Total Contigs | GC Content | N50 |
|---|---|---|---|---|---|---|---|---|
| Daerent | 45,506,199 | 12,929 | 35.6 | 36,248,481 | 79.66% | 5,469,310 | 63.70% | 271 |
| Hueschterbach | 42,222,600 | 13,046 | 35.88 | 36,987,700 | 87.60% | 5,727,441 | 63.95% | 269 |
| Koulbich | 64,930,676 | 18,587 | 35.53 | 49,069,306 | 75.57% | 7,792,136 | 63.18% | 268 |
| Mollbach | 50,983,302 | 14,522 | 35.63 | 35,064,164 | 68.77% | 6,306,081 | 63.40% | 264 |
| Pall 1 | 46,905,743 | 13,397 | 35.62 | 40,072,879 | 85.43% | 5,980,767 | 64.72% | 273 |
| Pall 2 | 56,510,613 | 16,136 | 35.6 | 48,535,078 | 85.89% | 6,883,923 | 63.40% | 272 |
| Retgenbusch | 49,316,386 | 13,878 | 35.59 | 46,692,284 | 94.68% | 5,677,103 | 60.01% | 295 |
| Weierbach | 57,936,000 | 16,650 | 35.22 | 57,518,714 | 99.28% | 5,898,678 | 60.95% | 323 |
| Sites | Shannon Index (H) | Evenness (E) | Species Richness (S) |
|---|---|---|---|
| Daerent | 2.39 | 0.48 | 147 |
| Hueschterbach | 2.54 | 0.51 | 140 |
| Koulbich | 2.58 | 0.51 | 149 |
| Mollbach | 2.79 | 0.55 | 155 |
| Pall 1 | 2.65 | 0.52 | 160 |
| Pall 2 | 2.62 | 0.51 | 174 |
| Retgenbusch | 2.31 | 0.49 | 117 |
| Weierbach | 2.83 | 0.63 | 91 |
| Environmental Variables | p-Value |
|---|---|
| Vegetation | 0.075 |
| Ca2+ | 0.008 ** |
| Carbon-to-nitrogen ratio | 0.349 |
| pH | 0.003 ** |
| Silt content | 0.125 |
| Sand content | 0.764 |
| Clay content | 0.901 |
| Mg content | 0.083 |
| GWC | 0.846 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Florent, P.; Cauchie, H.-M.; Herold, M.; Jacquet, S.; Ogorzaly, L. Soil pH, Calcium Content and Bacteria as Major Factors Responsible for the Distribution of the Known Fraction of the DNA Bacteriophage Populations in Soils of Luxembourg. Microorganisms 2022, 10, 1458. https://doi.org/10.3390/microorganisms10071458
Florent P, Cauchie H-M, Herold M, Jacquet S, Ogorzaly L. Soil pH, Calcium Content and Bacteria as Major Factors Responsible for the Distribution of the Known Fraction of the DNA Bacteriophage Populations in Soils of Luxembourg. Microorganisms. 2022; 10(7):1458. https://doi.org/10.3390/microorganisms10071458
Chicago/Turabian StyleFlorent, Perrine, Henry-Michel Cauchie, Malte Herold, Stéphan Jacquet, and Leslie Ogorzaly. 2022. "Soil pH, Calcium Content and Bacteria as Major Factors Responsible for the Distribution of the Known Fraction of the DNA Bacteriophage Populations in Soils of Luxembourg" Microorganisms 10, no. 7: 1458. https://doi.org/10.3390/microorganisms10071458