
Effects of Inhibitors Generated by Dilute Phosphoric Acid Plus Steam-Exploded Poplar on Saccharomyces cerevisiae Growth

Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Microorganism and Cultivation
2.3. PASE Spent Liquor Composition
2.4. S. cerevisiae 1517RM Biomass Measurement
2.5. Zeta Potential and Surface Morphology Characterization
2.6. Intracellular Ergosterol Determination
3. Results and Discussion
3.1. PASE Spent Liquor Composition
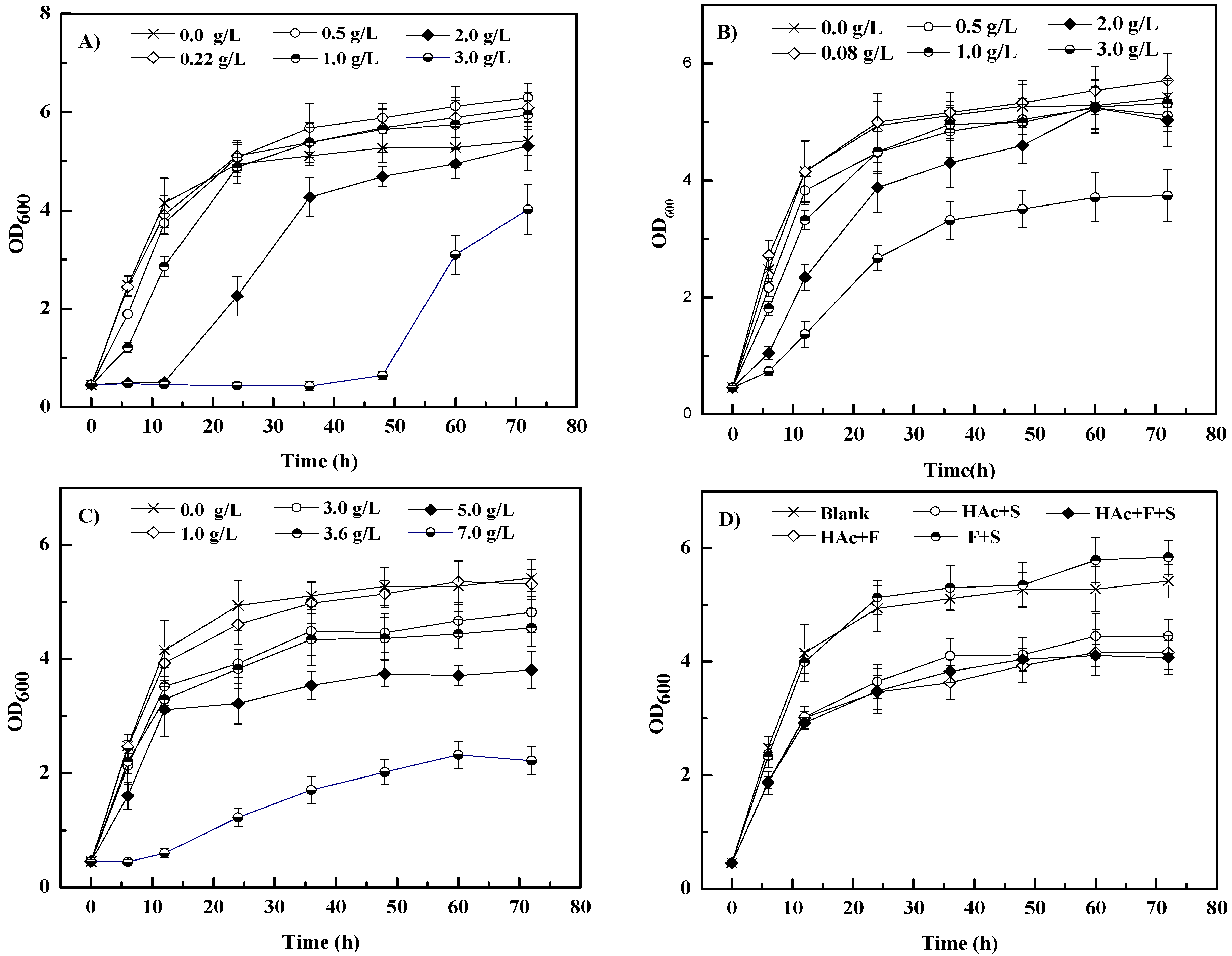
3.2. Effects of Inhibitors on S. cerevisiae 1517RM Growth
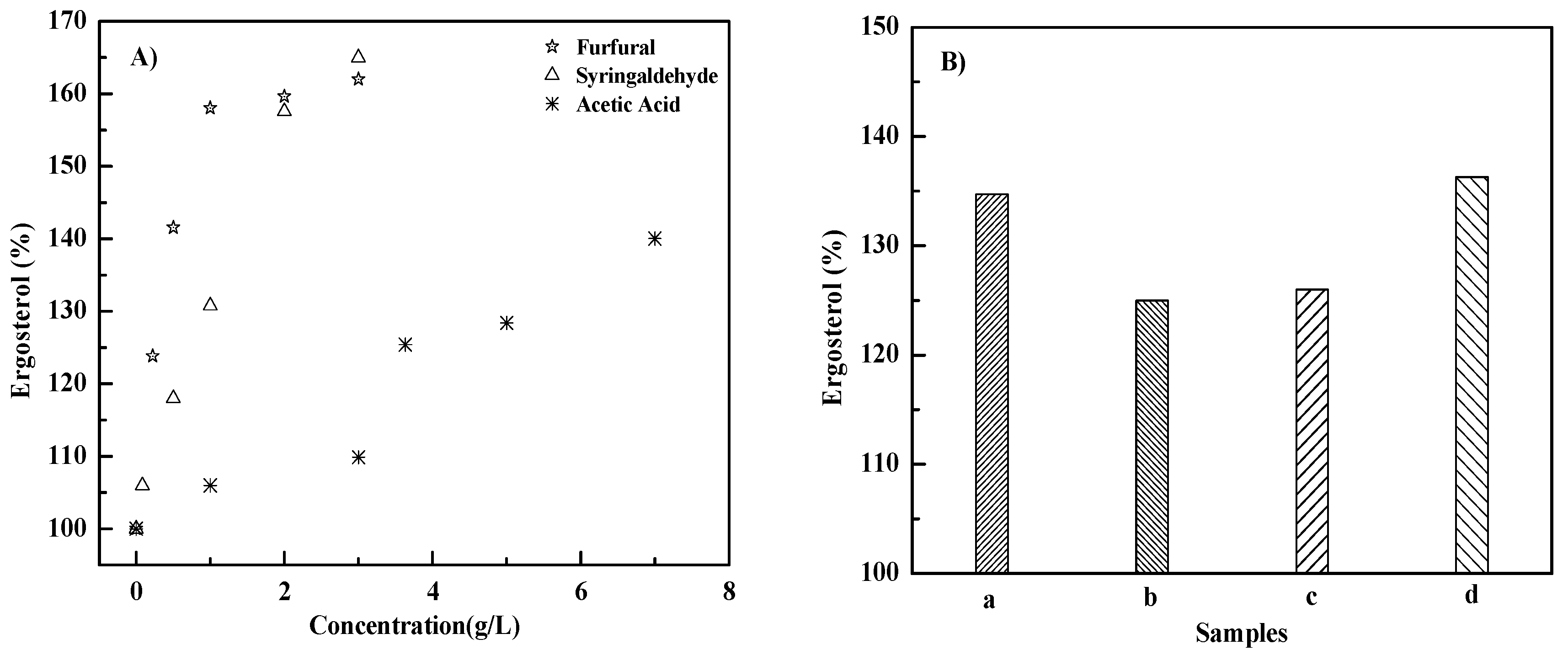
3.3. Effects of Inhibitors on Ergosterol Content
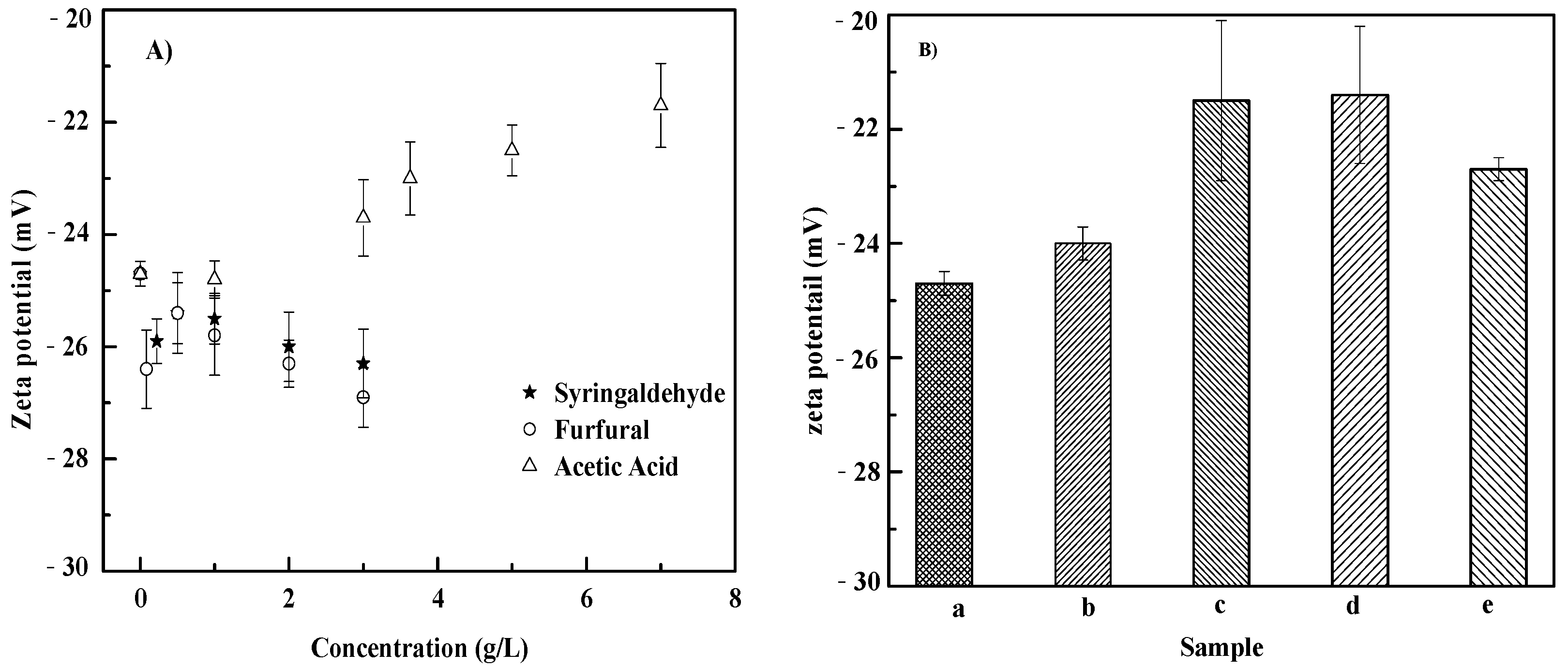
3.4. Zeta Potential on of S. cerevisiae Cells
3.5. Surface Morphologies of S. cerevisiae Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banu, J.R.; Kavitha, P.S.; Tyagi, V.K.; Gunasekaran, M.; Karthikeyan, O.P.; Kumar, G. Lignocellulosic biomass based biorefinery: A successful platform towards circular bioeconomy. Fuel 2021, 302, 121086. [Google Scholar] [CrossRef]
- Haldar, D.; Purkait, M.K. A review on the environment-friendly emerging techniques for pretreatment of lignocellulosic biomass Mechanistic insight and advancements. Chemosphere 2021, 264, 128523. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, N.; Maniet, G.; Vanderghem, C.; Delvigne, F.; Richel, A. Application of Steam Explosion as Pretreatment on Lignocellulosic Material—A Review. Ind. Eng. Chem. Res. 2015, 54, 2593–2598. [Google Scholar] [CrossRef]
- Pitarelo, A.P.; Fonseca, C.S.; Chiarello, L.M.; Gírio, F.M.; Ramos, L.P. Ethanol production from sugarcane bagasse using phosphoric acid-catalyzed steam explosion. J. Braz. Chem. Soc. 2016, 27, 1889–1898. [Google Scholar] [CrossRef]
- Fockink, D.H.; Sánchez, J.H.; Ramos, L.P. Comprehensive analysis of sugarcane bagasse steam explosion using autocatalysis and dilute acid hydrolysis (H3PO4 and H2SO4) at equivalent combined severity factors. Ind. Crops Prod. 2018, 123, 563–572. [Google Scholar] [CrossRef]
- Zeng, J.; Tong, Z.; Wang, L.; Zhu, J.; Ingram, L. Isolation and structural characterization of sugarcane bagasse lignin after dilute phosphoric acid plus steam explosion pretreatment and its effect on cellulose hydrolysis. Bioresour. Technol. 2014, 154, 274–281. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Van der Pol, E.C.; Bakker, R.R.; Baets, P.; Eggink, G. By-products resulting from lignocellulose pretreatment and their inhibitory effect on fermentations for (bio) chemicals and fuels. Appl. Microbiol. Biotechnol. 2014, 98, 9579–9593. [Google Scholar] [CrossRef]
- Klinke, H.B.; Thomsen, A.B.; Ahring, B.K. Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pretreatment of biomass. Appl. Microbiol. Biotechnol. 2004, 66, 10–26. [Google Scholar] [CrossRef]
- Sjulander, N.; Kikas, T. Origin, Impact and Control of Lignocellulosic Inhibitors in Bioethanol Production—A Review. Energies 2020, 13, 4751. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Sheng, J.Y.; Jiang, T.; Stevens, J.; Feng, X.Y.; Wei, N. Transcriptional profiling reveals molecular basis and novel genetic targets for improved resistance to multiple fermentation inhibitors in Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, P.; Zhang, Y.; Suo, Y.; Liao, Z.; Ma, Y.; Fu, H.; Wang, J. The global regulator IrrE from Deinococcus radiodurans enhances the furfural tolerance of Saccharomyces cerevisiae. Biochem. Eng. J. 2018, 136, 69–77. [Google Scholar] [CrossRef]
- Gorsich, S.W.; Dien, B.S.; Nichols, N.N.; Slininger, P.J.; Liu, Z.L.; Skory, C.D. Tolerance to furfural-induced stress is associated with pentose phosphate pathway genes ZWF1, GND1, RPE1, and TKL1 in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 71, 339–349. [Google Scholar] [CrossRef]
- Liu, Z.L.; Moon, J. A novel NADPH-dependent aldehyde reductase gene from Saccharomyces cerevisiae NRRL Y-12632 involved in the detoxi- fication of aldehyde inhibitors derived from lignocellulosic biomass conversion. Gene 2009, 446, 1–10. [Google Scholar] [CrossRef]
- Allen, S.A.; Clark, W.; Mccaffery, J.M.; Zhen, C.; Gorsich, S.W. Furfural induces reactive oxygen species accumulation and cellular damage in Saccharomyces cerevisiae. Biotechnol. Biofuels 2010, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Mollapour, M.; Piper, P.W. Hog1 mitogen-activated protein kinase phosphorylation targets the yeast Fps1 aquaglyceroporin for endocytosis, thereby rendering cells resistant to acetic acid. Mol. Cell Biol. 2007, 27, 6446–6456. [Google Scholar] [CrossRef] [Green Version]
- Li, B.Z.; Yuan, Y.J. Transcriptome shifts in response to furfural and acetic acid in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2010, 86, 1915–1924. [Google Scholar] [CrossRef]
- Mira, N.P.; Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Genome-wide identifcation of Saccharomyces cerevisiae genes required for tolerance to acetic acid. Microb. Cell Fact. 2010, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Martıín, C.; Jönsson, L.J. Comparison of the resistance of industrial and laboratory strains of Saccharomyces and Zygosaccharomyces to lignocellulose-derived fermentation inhibitors. Enzyme Microb. Technol. 2003, 32, 386–395. [Google Scholar] [CrossRef]
- Almeida, J.R.; Modig, T.; Petersson, A.; Hähn-Hägerdal, B.; Lidén, G.; Gorwa-Grauslund, M.F. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2007, 82, 340–349. [Google Scholar] [CrossRef]
- Wang, S.; Sun, X.; Yuan, Q. Strategies for enhancing microbial tolerance to inhibitors for biofuel production: A review. Bioresour. Technol. 2018, 258, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Nakamura, T.; Ando, A.; Tokuyasu, K.; Shima, J. Genome-wide screening of the genes required for tolerance to vanillin, which is a potential inhibitor of bioethanol fermentation, in Saccharomyces cerevisiae. Biotechnol. Biofuels 2008, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.J.; Yi, C.F.; Li, H. Changes of Saccharomyces cerevisiae cell membrane components and promotion to ethanol tolerance during the bioethanol fermentation. Int. J. Biochem. Cell B 2015, 69, 196–203. [Google Scholar] [CrossRef]
- Endo, A.; Nakamura, T.; Shima, J. Involvement of ergosterol in tolerance to vanillin, a potential inhibitor of bioethanol fermentation, in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2009, 299, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Chen, J.; Zhan, P.; Zhang, L.; Zhou, X.; Zeng, B.; Wu, Z.; Wang, H. Optimization of Mixed Enzymolysis of Acid-exploded Poplar Wood Residues for Directional Bioconversion. Bioresources 2020, 15, 1945–1958. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 2008, 1617, 1–16. [Google Scholar]
- Gu, H.; Zhu, Y.; Peng, Y.; Liang, X.; Liu, X.; Shao, L.; Xu, Y.; Xu, Z.; Liu, R.; Li, J. Physiological mechanism of improved tolerance of Saccharomyces cerevisiae to lignin-derived phenolic acids in lignocellulosic ethanol fermentation by short-term adaptation. Biotechnol. Biofuels 2019, 14, 268. [Google Scholar] [CrossRef] [Green Version]
- Arthington-Skaggs, B.A.; Jradi, H.; Desai, T.; Morrison, C.J. Quantitation of Ergosterol Content: Novel Method for Determination of Fluconazole Susceptibility of Candida albicans. J. Clin. Microbiol. 1999, 37, 3332–3337. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Chen, X.; Du, X.; Zhou, G.; Deng, Y.; Huang, Z. Effects of synthetic defects of very long-chain fatty acids on cell membrane stability and sensitivity of polyenes in yeast. Microbiol. China 2019, 46, 1645–1653. [Google Scholar]
- Koo, B.; Park, N.; Jeong, H.; Choi, J.; Yeo, H.; Choi, I. Characterization of by-products from organosolv pretreatments of yellow poplar wood (Liriodendron tulipifera) in the presence of acid and alkali catalysts. J. Ind. Eng. Chem. 2011, 17, 18–24. [Google Scholar] [CrossRef]
- Oliva, J.M.; Sáez, F.; Ballesteros, I.; González, A.; Negro, M.J.; Manzanares, P.; Ballesteros, M. Effect of lignocellulosic degradation compounds from steam explosion pretreatment on ethanol fermentation by thermotolerant yeast Kluyveromyces marxianus. In Biotechnology for Fuels and Chemicals; Davison, B.H., Lee, J.W., Finkelstein, M., McMillan, J.D., Eds.; Springer Book Archive; Humana Press: Totowa, NJ, USA, 2003; pp. 141–153. [Google Scholar]
- Zhang, J.; Gu, F.; Zhu, J.Y.; Zalesny, R.S. Using a combined hydrolysis factor to optimize high titer ethanol production from sulfite-pretreated poplar without detoxification. Bioresour. Technol. 2015, 186, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Kundu, C.; Jeong, S.; Lee, J. Bioethanol production from deacetylated yellow poplar pretreated with oxalic acid recovered through electrodialysis. Bioresour. Technol. 2016, 208, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Weil, J.; Sarikaya, A.; Rau, S.L.; Goetz, J.; Ladisch, C.M.; Brewer, M.; Hendrickson, R.; Ladisch, M.R. Pretreatment of yellow poplar sawdust by pressure cooking in water. Appl. Biochem. Biotechnol. 1997, 68, 21–40. [Google Scholar] [CrossRef]
- Pažitný, A.; Russ, A.; Boháček, Š.; Stankovská, M.; Ihnát, V. Effect of steam explosion on enzymatic hydrolysis of various parts of poplar tree. Wood Res. 2020, 65, 579–590. [Google Scholar] [CrossRef]
- Da Silva, T.L.; Santo, R.; Reis, A.; Passarinho, P.C. Effect of Furfural on Saccharomyces carlsbergensis Growth, Physiology and Ethanol Production. Appl. Biochem. Biotechnol. 2017, 182, 708–720. [Google Scholar] [CrossRef]
- Clarkson, S.M.; Hamilton-Brehm, S.D.; Giannone, R.J.; Engle, N.L.; Tschaplinski, T.J.; Hettich, R.L.; Elkins, J.G. A comparative multidimensional LC-MS proteomic analysis reveals mechanisms for furan aldehyde detoxification in Thermoanaerobacter pseudethanolicus 39E. Biotechnol. Biofuels 2014, 7, 165. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cheng, G.; Joshua, C.; He, Z.; Sun, X.; Li, R.; Liu, L.; Yuan, Q. Furfural tolerance and detoxification mechanism in Candida tropicalis. Biotechnol. Biofuels 2016, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Taherzadeh, M.J.; Gustafsson, L.; Niklasson, C.; Liden, G. Conversion of furfural in aerobic and anaerobic batch fermentation of glucose by Saccharomyces cerevisiae. J. Biosci. Bioeng. 1990, 87, 169–174. [Google Scholar] [CrossRef]
- Horváth, I.S.; Franzén, C.J.; Taherzadeh, M.J.; Niklasson, C.; Lidén, G. Effects of furfural on the respiratory metabolism of Saccharomyces cerevisiae in glucose-limited chemostats. Appl. Environ. Microb. 2003, 69, 4076–4086. [Google Scholar] [CrossRef] [Green Version]
- Colombi, B.L.; Zanoni, P.R.S.; Tavares, L.B.B. Effect of phenolic compounds on bioconversion of glucose to ethanol by yeast Saccharomyces cerevisiae PE-2. Can. J. Chem. Eng. 2018, 96, 1444–1450. [Google Scholar] [CrossRef]
- Kelly, C.; Jones, O.; Barnhart, C.; Lajoie, C. Effect of Furfural, Vanillin and Syringaldehyde on Candida guilliermondii Growth and Xylitol Biosynthesis. In Biotechnology for Fuels and Chemicals; Adney, W.S., McMillan, J.D., Mielenz, J., Klasson, K.T., Eds.; E-Publishing Inc.: Chicago, IL, USA; Humana Press: Totowa, NJ, USA, 2008; pp. 615–626. [Google Scholar]
- Yu, X.; Zeng, J.; Zheng, Y.; Chen, S. Effect of lignocellulose degradation products on microbial biomass and lipid production by the oleaginous yeast Cryptococcus curvatus. Process Biochem. 2014, 49, 457–465. [Google Scholar] [CrossRef]
- Ullah, A.; Orij, R.; Brul, S.; Smits, G.J. Quantitative analysis of the modes of growth inhibition by weak organic acids in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 8377–8387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Holzwarth, G.; Penner, M.H.; Patton-Vogt, J.; Bakalinsky, A.T. Overexpression of acetyl-CoA synthetase in Saccharomyces cerevisiae increases acetic acid tolerance. FEMS Microbiol. Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Feng, H.; Yuan, W.; Li, Y.; Hou, S.; Zhong, S.; Bai, F. Enhanced fermentative performance under stresses of multiple lignocellulose-derived inhibitors by overexpression of a typical 2-Cys peroxiredoxin from Kluyveromyces marxianus. Biotechnol. Biofuels 2017, 10, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannattasio, S.; Guaragnella, N.; Ždralević, M.; Marra, E. Molecular mechanisms of Saccharomyces cerevisiae stress adaptation and programmed cell death in response to acetic acid. Front. Microbiol. 2013, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, G.; Pais, C. The Influence of Acetic and Other Weak Carboxylic Acids on Growth and Cellular Death of the Yeast Yarrowia lipolytica. Food Technol. Biotechnol. 2000, 38, 27–32. [Google Scholar]
- Spagnuolo, M.; Shabbir Hussain, M.; Gambill, L.; Blenner, M. Alternative Substrate Metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Greetham, D.; Hart, A.J.; Tucker, G.A. Presence of low concentrations of acetic acid improves yeast tolerance to hydroxymethylfurfural (HMF) and furfural. Biomass Bioenerg. 2016, 85, 53–60. [Google Scholar] [CrossRef]
- Zinser, E.; Sperka-Gottlieb, C.D.; Fasch, E.V.; Kohlwein, S.D.; Paltauf, F.; Daum, G. Phospholipid synthesis and lipid composition of subcellular membranes in the unicellular eukaryote Saccharomyces cerevisiae. J. Bacteriol. 1991, 173, 2026–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandre, H.; Rousseaux, I.; Charpentier, C. Relationship between ethanol tolerance, lipid composition and plasma membrane fluidity in Saccharomyces cerevisiae and Kloeckera apiculata. FEMS Microbiol. Lett. 1994, 124, 17–22. [Google Scholar] [CrossRef]
- Murga, M.L.F.; De Valdez, G.F.; Disalvo, A.E. Changes in the surface potential of Lactobacillus acidophilus under freeze-thawing stress. Cryobiology 2000, 41, 10–16. [Google Scholar] [CrossRef]
- Lavaisse, L.M.; Hollmann, A.; Nazareno, M.A.; Disalvo, E.A. Zeta potential changes of Saccharomyces cerevisiae during fermentative and respiratory cycles. Colloid. Surface B 2019, 174, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Monlau, F.; Steyer, J.P.; Carrere, H. Effect of lignin-derived and furan compounds found in lignocellulosic hydrolysates on biomethane production. Bioresour. Technol. 2012, 104, 90–99. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GC- MS | Retention Time (min) | Compound | Origin | Retention Time (min) | Compound | Origin |
| 1.878 | Furfural (C5H4O2) | Pentose (Hemicellulose) | 8.365 | Vanillin (C8H8O3) | Guaiacyl unit (Lignin) | |
| 3.217 | Phenol (C6H6O) | ρ-Coumaryl unit (Lignin) | 11.187 | Vanillic acid (C8H8O4) | Guaiacyl unit (Lignin) | |
| 5.695 | Benzoic acid (C7H6O2) | ρ-Coumaryl unit (Lignin) | 11.593 | Syringaldehyde (C9H10O4) | Syringyl unit (Lignin) | |
| 6.069 | 5-HMF (C6H6O3) | Hexose (Cellulose) | 12.473 | Coniferyl aldehyde (C10H10O3) | Guaiacyl unit (Lignin) | |
| HPLC | Compound | Origin | Concentration (g/L) | |||
| Furfural (C5H4O2) | Pentose (Hemicellulose) | 0.22 | ||||
| 5-HMF (C6H6O3) | Hexose (Cellulose) | 0.19 | ||||
| Vanillin (C8H8O3) | Guaiacyl unit (Lignin) | 0.04 | ||||
| Syringaldehyde (C9H10O4) | Syringyl unit (Lignin) | 0.08 | ||||
| Acetic acid (C2H4O2) | (Hemicellulose/Lignin) | 3.63 | ||||
| Glucose (C6H12O6) | (Cellulose/Hemicellulose) | 5.11 | ||||
| Xylose (C5H10O5) | (Hemicellulose) | 14.88 | ||||
| Biomass | Pretreatment Conditions | By-Products (g/L) | Reference |
|---|---|---|---|
| Yellow poplar wood (Liriodendron tulipifera) | Pressure cooking: less than 40 mesh size, 1:10 substrates/solvent (50% aqueous ethanol), 1% sodium hydroxide, 140–160 °C, 10 min. | Acetic acid: 4.2–4.9 | [31] |
| Yellow poplar | Pressure cooking: 1 × 5 mm particle size, 6.0–6.6% (w/w) solids content, 220–260 °C, 70–75 min. | Furfural: 1.6–2.7; 5-HMF: 0.90–1.55; Acetic acid: 2.3–3.1 | [32] |
| Poplar NE222 (Populus deltoides Bartr. ex Marsh × Populus nigra L.) | Sulfate pretreatment: 6–38 mm wood chips with 1–5 mm thicknesses, 25% (w/w) total solids loading, 2 mL/L sulfuric acid, 135 °C, 290 min. | Furfural: 0.9; Acetic acid: 16.7 | [33] |
| Yellow poplar wood (Liriodendron tulipifera) | Pre-deacetylation with NaOH, Oxalic acid hydrothermal pretreatment: 20–80 mesh size, 0.16 mol/L C2H2O4, 150 °C, 42 min. | Furfural: 2.73–3.09; 5-HMF: 0.16–0.21; Acetic acid:not detectable | [34] |
| Poplar | Steam explosion: 210 °C and 4 min residence time. | Furfural: 0.49; 5-HMF: 0.08; Acetic acid: 2.1 | [35] |
| White poplar (Populus alba L.) | Steam explosion: 0.7 mm particle size, pre-soaked in water for at least 1 h, 205 °C, 10 min residence time. | Furfural: 1.6; 5-HMF:0.6; Acetic acid: 4.9 (sapwood) and 6.0 (coppice) | [36] |
| Inhibitor | Inhibitor (g/L) | Biomass * (OD) | Inhibitor | Inhibitor (g/L) | Biomass * (OD) |
|---|---|---|---|---|---|
| Blank | 0.00 | 5.42 ± 0.32 | Blank | 0.00 | 5.42 ± 0.32 |
| Furfural | 0.22 | 6.09 ± 0.34 | Syringaldehyde | 0.08 | 5.71 ± 0.46 |
| 0.50 | 6.29 ± 0.36 | 0.50 | 5.11 ± 0.28 | ||
| 1.00 | 5.94 ± 0.32 | 1.00 | 5.32 ± 0.39 | ||
| 2.00 | 5.31 ± 0.54 | 2.00 | 5.03 ± 0.45 | ||
| 3.00 | 4.02 ± 0.53 | 3.00 | 3.74 ± 0.44 | ||
| Acetic acid | 1.00 | 5.31 ± 0.27 | Inhibitor cocktail | a | 4.16 ± 0.38 |
| 3.00 | 4.80 ± 0.36 | b | 4.45 ± 0.35 | ||
| 3.63 | 4.55 ± 0.33 | c | 5.84 ± 0.37 | ||
| 5.00 | 3.81 ± 0.32 | d | 4.07 ± 0.34 | ||
| 7.00 | 2.22 ± 0.24 | / | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhan, P.; Shao, L.; Zhang, L.; Qing, Y. Effects of Inhibitors Generated by Dilute Phosphoric Acid Plus Steam-Exploded Poplar on Saccharomyces cerevisiae Growth. Microorganisms 2022, 10, 1456. https://doi.org/10.3390/microorganisms10071456
Wang Y, Zhan P, Shao L, Zhang L, Qing Y. Effects of Inhibitors Generated by Dilute Phosphoric Acid Plus Steam-Exploded Poplar on Saccharomyces cerevisiae Growth. Microorganisms. 2022; 10(7):1456. https://doi.org/10.3390/microorganisms10071456
Chicago/Turabian StyleWang, Yanan, Peng Zhan, Lishu Shao, Lin Zhang, and Yan Qing. 2022. "Effects of Inhibitors Generated by Dilute Phosphoric Acid Plus Steam-Exploded Poplar on Saccharomyces cerevisiae Growth" Microorganisms 10, no. 7: 1456. https://doi.org/10.3390/microorganisms10071456