
Metagenomic Analysis of the Species Composition and Seasonal Distribution of Marine Dinoflagellate Communities in Four Korean Coastal Regions


Abstract
:1. Introduction
2. Materials and Methods
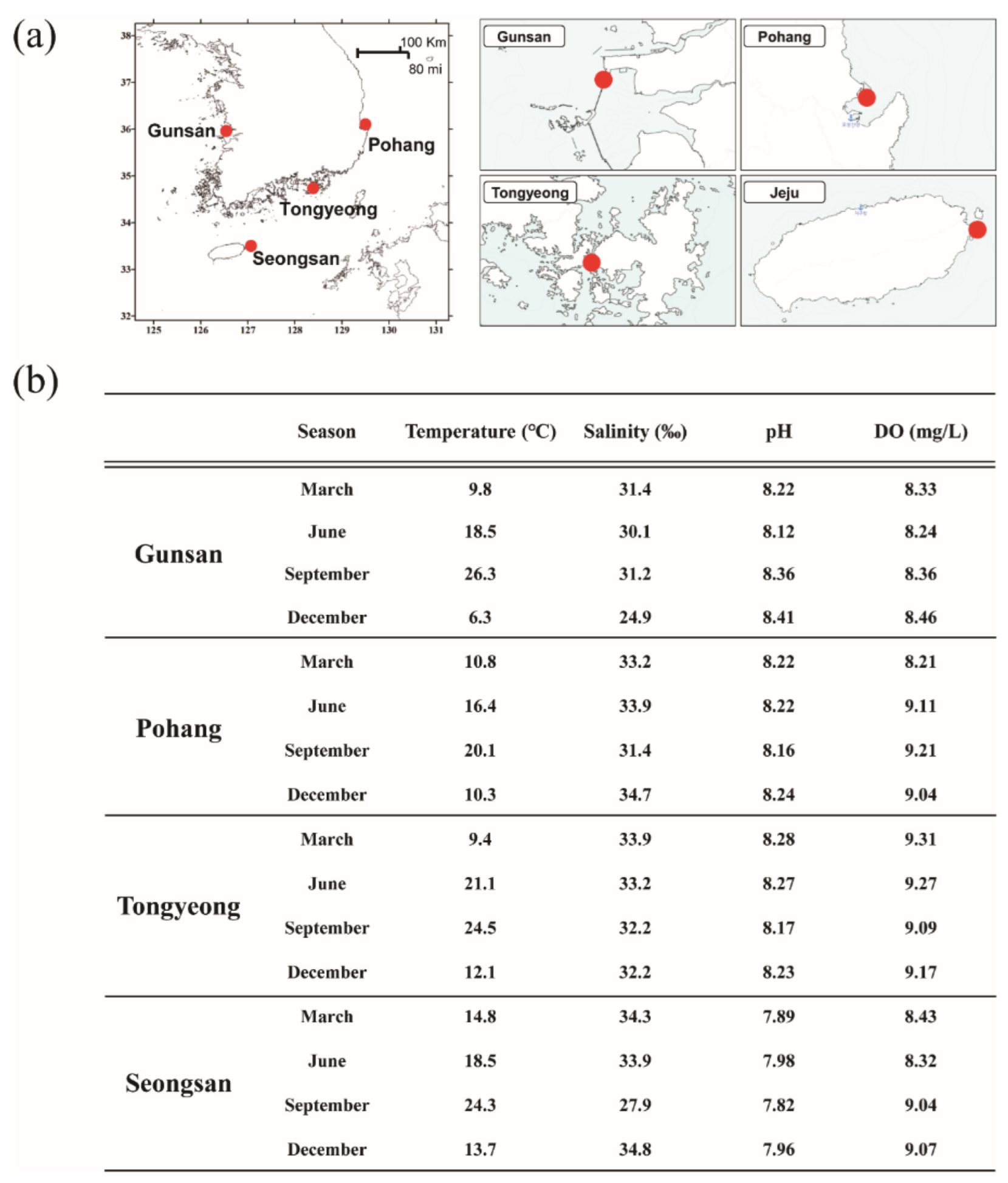
2.1. Study Areas and Seawater Sample Collecting
2.2. DNA Extraction, Library Preparation, and NGS
2.3. Customized Dinoflagellate Reference Databases for CLC Workflows
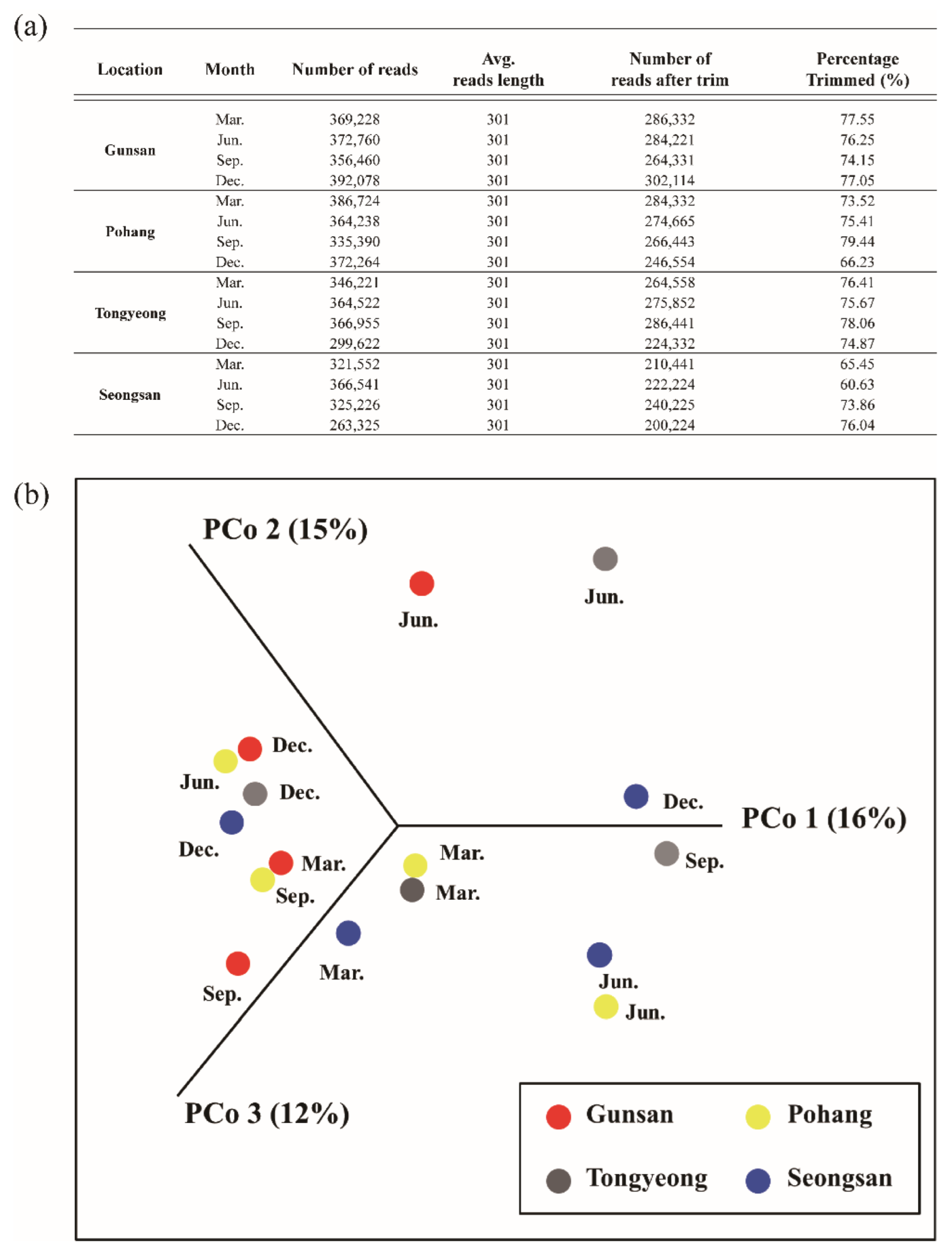
2.4. Data Quality Control and Taxonomic Profiling
3. Results
3.1. Environmental Characteristics of Sampling Sites
3.2. Metagenome Comparisons
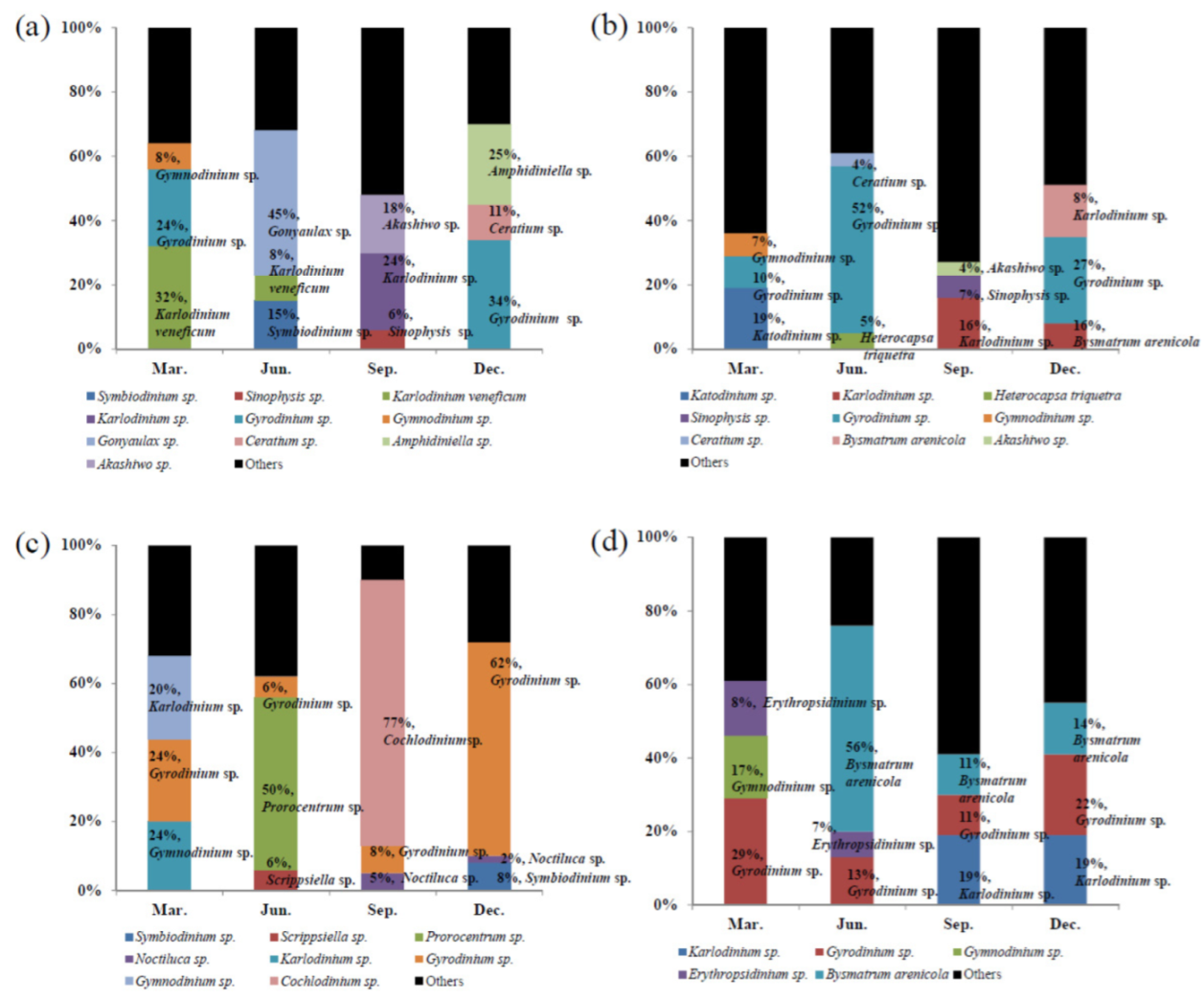
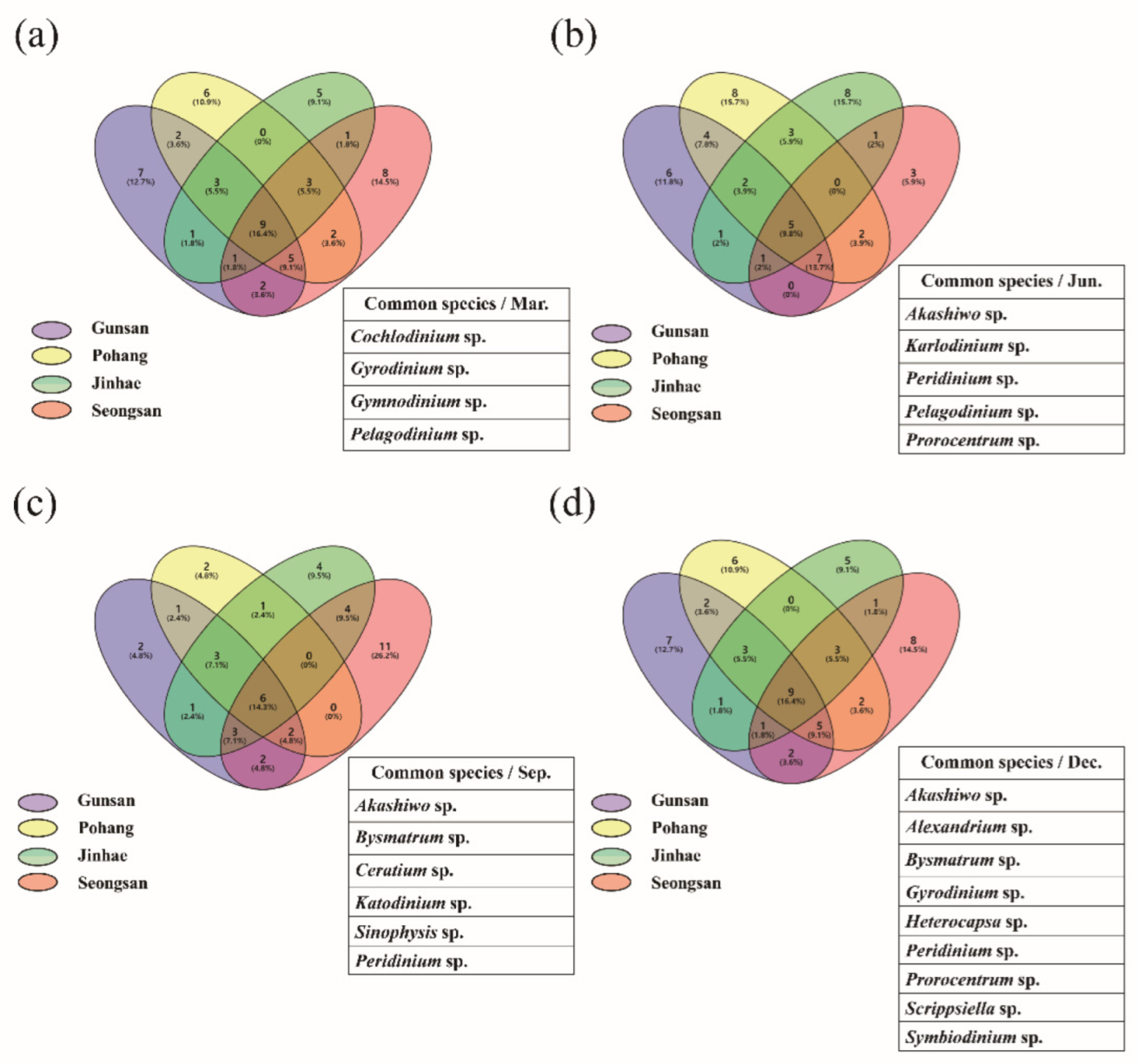
3.3. Metagenomic Analysis of the Dinoflagellate Species Composition
3.4. Comparison of Metagenomic Analysis and Microscopic Observation
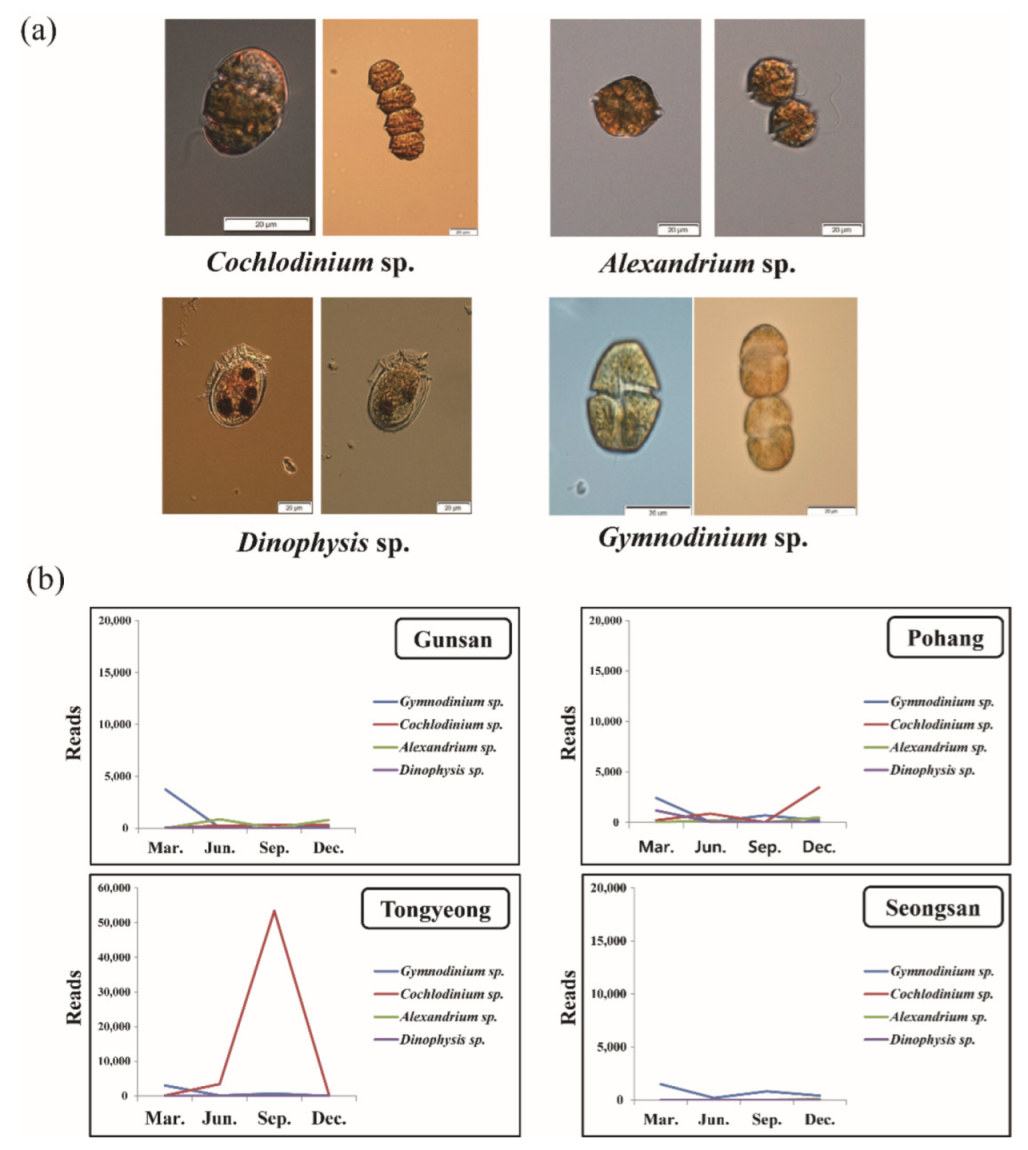
3.5. Seasonal Distribution of Harmful Species Based on Metagenomic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lessard, E.J. Oceanic Heterotrophic Dinoflagellates: Distribution, Abundance, and Role as Microzooplankton; Rhode Island University: Kingston, NY, USA, 1984. [Google Scholar]
- Jeong, H.J.; Yoo, Y.D.; Kim, J.S.; Seong, K.A.; Kang, N.S.; Kim, T.H. Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci. J. 2010, 45, 65–91. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.-K.; Ortiz, L.C.; Vukasin, G.D.; Chen, Y.; Shin, D.D.; Kenny, T.W. An oven-controlled MEMS oscillator (OCMO) with sub 10 MW, ±1.5 ppb stability over temperature. In Proceedings of the 2019 20th International Conference on Solid-State Sensors, Actuators and Microsystems & Eurosensors XXXIII (TRANSDUCERS & EUROSENSORS XXXIII), Berlin, Germany, 23–27 June 2019; pp. 2072–2075. [Google Scholar]
- Zohdi, E.; Abbaspour, M. Harmful algal blooms (red tide): A review of causes, impacts and approaches to monitoring and prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Couet, D.; Pringault, O.; Bancon-Montigny, C.; Briant, N.; Poulichet, F.E.; Delpoux, S.; Yahia, O.K.-D.; Hela, B.; Herve, F.; Rovillon, G. Effects of copper and butyltin compounds on the growth, photosynthetic activity and toxin production of two HAB dinoflagellates: The planktonic Alexandrium catenella and the benthic Ostreopsis cf ovata. Aquat. Toxicol. 2018, 196, 154–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Pančić, M.; Andersen, K.H.; Kiørboe, T. The cost of toxin production in phytoplankton: The case of PST producing dinoflagellates. ISME J. 2019, 13, 64–75. [Google Scholar] [CrossRef]
- Jantschke, A.; Pinkas, I.; Schertel, A.; Addadi, L.; Weiner, S. Biomineralization pathways in calcifying dinoflagellates: Uptake, storage in MgCaP-rich bodies and formation of the shell. Acta Biomater. 2020, 102, 427–439. [Google Scholar] [CrossRef]
- Choi, H.; Liu, X.; Kim, H.I.; Kim, D.; Park, T.; Song, S. A Facile Surface Passivation Enables Thermally Stable and Efficient Planar Perovskite Solar Cells Using a Novel IDTT-Based Small Molecule Additive. Adv. Energy Mater. 2021, 11, 2003829. [Google Scholar] [CrossRef]
- Chan, K.K.; Tan, T.J.; Narayanan, K.K.; Procko, E. An engineered decoy receptor for SARS-CoV-2 broadly binds protein S sequence variants. Sci. Adv. 2021, 7, eabf1738. [Google Scholar] [CrossRef]
- Lee, E.S.; Kim, E.-K.; Choi, Y.-H.; Jung, Y.H.; Kim, S.Y.; Koh, J.W.; Choi, E.K.; Cheon, J.-E.; Kim, H.-S. Factors associated with neurodevelopment in preterm infants with systematic inflammation. BMC Pediatrics 2021, 21, 114. [Google Scholar] [CrossRef]
- Muhammad, B.L.; Kim, T.; Ki, J.-S. 18S rRNA analysis reveals high diversity of phytoplankton with emphasis on a naked Dinoflagellate Gymnodinium sp. at the Han river (Korea). Diversity 2021, 13, 73. [Google Scholar] [CrossRef]
- Akbar, M.A.; Sang, J.; Khan, A.A.; Amin, F.-E.; Hussain, S.; Sohail, M.K.; Xiang, H.; Cai, B. Statistical analysis of the effects of heavyweight and lightweight methodologies on the six-pointed star model. IEEE Access 2018, 6, 8066–8079. [Google Scholar] [CrossRef]
- Gran-Stadniczeñko, S.; Egge, E.; Hostyeva, V.; Logares, R.; Eikrem, W.; Edvardsen, B. Protist diversity and seasonal dynamics in Skagerrak plankton communities as revealed by metabarcoding and microscopy. J. Eukaryot. Microbiol. 2019, 66, 494–513. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Kuczynski, J.; Stombaugh, J.; Walters, W.A.; González, A.; Caporaso, J.G.; Knight, R. Using QIIME to analyze 16S rRNA gene sequences from microbial communities. Curr. Protoc. Microbiol. 2012, S27, 1E.5.1–1E.5.20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, E.O.; Suggett, D.J.; Voolstra, C.R.; Pettay, D.T.; Clark, D.R.; Pogoreutz, C.; Sampayo, E.M.; Warner, M.E.; Smith, D.J. Coral microbiome composition along the northern Red Sea suggests high plasticity of bacterial and specificity of endosymbiotic dinoflagellate communities. Microbiome 2020, 8, 1–16. [Google Scholar]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative genomic and transcriptomic characterization of the toxigenic marine dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef] [Green Version]
- Eichholz, K.; Beszteri, B.; John, U. Putative monofunctional type I polyketide synthase units: A dinoflagellate-specific feature? PLoS ONE 2012, 7, e48624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, D.C.; Farinholt, N.; Gates, C.; Shumaker, A.; Wagner, N.E.; Bienfang, P.; Bhattacharya, D. Analysis of G. ambierdiscus transcriptome data supports ancient origins of mixotrophic pathways in dinoflagellates. Environ. Microbiol. 2016, 18, 4501–4510. [Google Scholar] [CrossRef]
- Liu, H.; Stephens, T.G.; González-Pech, R.A.; Beltran, V.H.; Lapeyre, B.; Bongaerts, P.; Cooke, I.; Aranda, M.; Bourne, D.G.; Forêt, S.; et al. Symbiodinium genomes reveal adaptive evolution of functions related to coral-dinoflagellate symbiosis. Commun. Biol. 2018, 1, 95. [Google Scholar] [CrossRef] [Green Version]
- Lim, A.S.; Jeong, H.J. Benthic dinoflagellates in Korean waters. Algae 2021, 36, 91–109. [Google Scholar] [CrossRef]
- Kim, H.S.; Yih, W.; Kim, J.H.; Myung, G.; Jeong, H.J. Abundance of epiphytic dinoflagellates from coastal waters off Jeju Island, Korea during autumn 2009. Ocean Sci. J. 2021, 46, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Kim, S.H.; Jung, M.M.; Lee, J.B. New record of dinoflagellates around Jeju Island. J. Ecol. Environ. 2013, 36, 273–291. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Rahman, M.; An, S.J.; Lee, J.B. Seasonal abundance of epiphytic dinoflagellates around coastal waters of Jeju Island, Korea. J. Mar. Sci. Technol. 2013, 21, 20. [Google Scholar]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.; Breiner, H.W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F. A list of free-living dinoflagellate species in the world’s oceans. Acta Bot. Croat. 2005, 64, 129–212. [Google Scholar]
- Llanos-Rivera, A.; Álvarez-Muñoz, K.; Astuya-Villalón, A.; López-Rosales, L.; García-Camacho, F.; Sánchez-Mirón, A.; Rodriguez, J. Sublethal Effects of the Toxic Alga Karlodinium veneficum on Fish. Res. Sq. 2021, 1, e1021203. [Google Scholar]
- Elleuch, J.; Ben Amor, F.; Barkallah, M.; Haj Salah, J.; Smith, K.F.; Aleya, L.; Fendri, I.; Abdelkafi, S. q-PCR-based assay for the toxic dinoflagellate Karenia selliformis monitoring along the Tunisian coasts. Environ. Sci. Pollut. Res. 2021, 28, 57486–57498. [Google Scholar] [CrossRef]
- Abad, D.; Albaina, A.; Aguirre, M.; Laza-Martínez, A.; Uriarte, I.; Iriarte, A.; Villate, F.; Estonba, A. Is metabarcoding suitable for estuarine plankton monitoring? A comparative study with microscopy. Mar. Biol. 2016, 163, 149. [Google Scholar] [CrossRef]
- Metfies, K.; von Appen, W.-J.; Kilias, E.; Nicolaus, A.; Nöthig, E.-M. Biogeography and photosynthetic biomass of arctic marine pico-eukaroytes during summer of the record sea ice minimum 2012. PLoS ONE 2016, 11, e0148512. [Google Scholar] [CrossRef] [Green Version]
- Langer, J.A.; Sharma, R.; Schmidt, S.I.; Bahrdt, S.; Horn, H.G.; Algueró-Muñiz, M.; Nam, B.; Achterberg, E.P.; Riebesell, U.; Boersma, M. Community barcoding reveals little effect of ocean acidification on the composition of coastal plankton communities: Evidence from a long-term mesocosm study in the Gullmar Fjord, Skagerrak. PLoS ONE 2017, 12, e0175808. [Google Scholar] [CrossRef]
- Chai, Z.Y.; He, Z.L.; Deng, Y.Y.; Yang, Y.F.; Tang, Y.Z. Cultivation of seaweed Gracilaria lemaneiformis enhanced biodiversity in a eukaryotic plankton community as revealed via metagenomic analyses. Mol. Ecol. 2018, 27, 1081–1093. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Richlen, M.L.; Brandt, M.E.; Smith, T.B. Effects of grazing, nutrients, and depth on the ciguatera-causing dinoflagellate Gambierdiscus in the US Virgin Islands. Mar. Ecol. Prog. Ser. 2015, 531, 91–104. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jeong, H.J.; Ok, J.H.; Kang, H.C.; You, J.H. Spatial-temporal distributions of the newly described mixotrophic dinoflagellate Gymnodinium smaydae in Korean coastal waters. Algae 2020, 35, 225–236. [Google Scholar] [CrossRef]
- Horiguchi, T.; Pienaar, R.N. Validation of Bysmatrum arenicola horiguchi et pienaar, sp. nov.(dinophyceae)(Note). J. Phycol. 2000, 36, 237. [Google Scholar] [CrossRef]
- Shearer, T.L.; Coffroth, M.A. DNA BARCODING: Barcoding corals: Limited by interspecific divergence, not intraspecific variation. Mol. Ecol. Resour. 2008, 8, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.S.; Jeong, H.J.; Jang, T.Y.; Kang, N.S.; Lee, S.Y.; Yoo, Y.D.; Kim, H.S. Morphology and molecular characterization of the epiphytic dinoflagellate Prorocentrum cf. rhathymum in temperate waters off Jeju Island, Korea. Ocean Sci. J. 2013, 48, 1–17. [Google Scholar] [CrossRef]
- Edvardsen, B.; Shalchian-Tabrizi, K.; Jakobsen, K.S.; Medlin, L.K.; Dahl, E.; Brubak, S.; Paasche, E. Genetic variability and molecular phylogeny of dinophysis species (dinophyceae) from norwegian waters inferred from single cell analyses of rDNA 1. J. Phycol. 2003, 39, 395–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Location | March | June | September | December | ||||
| Dinoflagellate Reads | Unidentified | Dinoflagellate Reads | Unidentified | Dinoflagellate Reads | Unidentified | Dinoflagellate Reads | Unidentified | |
| Gunsan | 47,588 | 16,263 | 36,787 | 3084 | 39,780 | 14,265 | 87,273 | 11,245 |
| Pohang | 34,641 | 16,445 | 76,439 | 12,304 | 7222 | 4621 | 75,121 | 14,332 |
| Tongyeong | 12,397 | 3606 | 22,549 | 2855 | 69,468 | 1425 | 69,819 | 14,224 |
| Seongsan | 8924 | 2060 | 60,769 | 8994 | 30,197 | 4962 | 42,927 | 8644 |
| (b) | ||||||||
| Location | March | June | September | December | ||||
| Proportion (%) | Species | Proportion (%) | Species | Proportion (%) | Species | Proportion (%) | Species | |
| Gunsan | 32.1 | Karlodinium veneficum | 45.2 | Gonyaulax sp. | 24.1 | Karlodinium sp. | 34.2 | Gyrodinium sp. |
| 24.2 | Gyrodinium sp. | 15.3 | Symbiodinium sp. | 17.6 | Akashiwo sp. | 24.8 | Amphidiniella sp. | |
| 7.9 | Gymnodinium sp. | 7.9 | Karlodinium veneficum | 5.5 | Sinophysis sp. | 10.5 | Ceratium sp. | |
| 0.4 | Noctiluca scintillans | 6.3 | Ceratium sp. | 4.2 | Peridinium sp. | 4.5 | Heterocapsa triquetra | |
| 0.3 | Symbiodinium sp. | 3.1 | Pelagodinium sp. | 2.9 | Scrippsiella trochoidea | 2.5 | Karlodinium sp. | |
| 0.3 | Protoperidinium sp. | 2.4 | Dissodinium pseudolunula | 2.5 | Katodinium sp. | 2.4 | Peridinium sp. | |
| 0.3 | Pelagodinium sp. | 2.4 | Alexandrium sp. | 1.7 | Pelagodinium sp. | 2.1 | Noctiluca scintillans | |
| 0.2 | Scrippsiella sp. | 1.7 | Gyrodinium sp. | 1.3 | Gyrodinium sp. | 1.3 | Katodinium sp. | |
| 0.2 | Dinophysis sp. | 1.2 | Azadinium sp. | 0.8 | Cochlodinium sp. | 0.9 | Akashiwo sp. | |
| 0.2 | Ceratium sp. | 1.1 | Amphidiniopsis sp. | 0.5 | Ceratium sp. | 0.7 | Gonyaulax sp. | |
| Pohang | 19.0 | Katodinium sp. | 52.1 | Gyrodinium sp. | 16.0 | Karlodinium sp. | 27.2 | Gyrodinium sp. |
| 10.1 | Gyrodinium sp. | 4.9 | Heterocapsa triquetra | 6.8 | Sinophysis sp. | 16.2 | Bysmatrum arenicola | |
| 6.9 | Gymnodinium sp. | 4.0 | Ceratium sp. | 4.1 | Akashiwo sp. | 7.7 | Karlodinium veneficum | |
| 5.4 | Azadinium sp. | 3.7 | Karlodinium sp. | 1.5 | Paragymnodinium sp. | 7.4 | Akashiwo sp. | |
| 3.3 | Dinophysis sp. | 2.8 | Heterocapsa circularisquama | 1.5 | Peridinium sp. | 7.3 | Ceratium sp. | |
| 1.5 | Pelagodinium sp. | 2.1 | Gonyaulax sp. | 1.0 | Amphidiniella sp. | 4.6 | Cochlodinium sp. | |
| 1.4 | Ceratium sp. | 1.9 | Pelagodinium sp. | 1.0 | Ceratium sp. | 2.9 | Azadinium sp. | |
| 0.9 | Gonyaulax spinifera | 1.1 | Prorocentrum sp. | 0.5 | Bysmatrum arenicola | 1.5 | Katodinium sp. | |
| 0.9 | Gonyaulax sp. | 0.9 | Peridinium sp. | 0.4 | Pelagodinium sp. | 0.6 | Alexandrium sp. | |
| 0.8 | Erythropsidinium sp. | 0.6 | Cochlodinium sp. | 0.4 | Scrippsiella trochoidea | 0.6 | Peridinium sp. | |
| Tong-yeong | 23.9 | Gyrodinium sp. | 50.2 | Prorocentrum sp. | 77.3 | Cochlodinium sp. | 62.3 | Gyrodinium sp. |
| 23.6 | Gymnodinium sp. | 5.9 | Gyrodinium sp. | 7.9 | Gyrodinium sp. | 7.5 | Symbiodinium sp. | |
| 19.7 | Karlodinium veneficum | 5.6 | Scrippsiella sp. | 5.0 | Noctiluca scintillans | 1.5 | Noctiluca scintillans | |
| 0.9 | Cochlodinium sp. | 5.3 | Karlodinium sp. | 3.7 | Bysmatrum arenicola | 0.6 | Karlodinium sp. | |
| 0.9 | Pelagodinium sp. | 3.8 | Noctiluca sp. | 1.7 | Protoperidinium sp. | 0.6 | Alexandrium sp. | |
| 0.6 | Noctiluca scintillans | 3.5 | Neoceratium sp. | 0.9 | Karlodinium sp. | 0.6 | Peridinium sp. | |
| 0.3 | Akashiwo sp. | 1.3 | Heterocapsa sp. | 0.5 | Ceratium sp. | 0.6 | Amphidiniopsis sp. | |
| 0.2 | Paragymnodinium sp. | 1.1 | Blastodinium sp. | 0.4 | Erythropsidinium sp. | 0.5 | Heterocapsa triquetra | |
| 0.2 | Pfiesteria piscicida | 1.1 | Protodinium sp. | 0.4 | Akashiwo sp. | 0.4 | Cochlodinium sp. | |
| 1.0 | Chytriodinium sp. | 0.3 | Heterocapsa triquetra | 0.2 | Scrippsiella trochoidea | |||
| Seong-san | 29.1 | Gyrodinium sp. | 56.1 | Bysmatrum arenicola | 19.2 | Karlodinium veneficum | 22.4 | Gyrodinium sp. |
| 16.6 | Gymnodinium sp. | 13.2 | Gyrodinium sp. | 11.1 | Bysmatrum arenicola | 19.3 | Karlodinium sp. | |
| 8.4 | Erythropsidinium sp. | 7.4 | Erythropsidinium sp. | 11.0 | Gyrodinium sp. | 13.9 | Bysmatrum arenicola | |
| 3.9 | Karlodinium sp. | 5.6 | Karlodinium sp. | 10.4 | Ceratium sp. | 6.8 | Ceratium sp. | |
| 1.7 | Heterocapsa sp. | 1.6 | Ceratium sp. | 8.2 | Peridinium sp. | 5.5 | Akashiwo sp. | |
| 0.7 | Paragymnodinium sp. | 1.3 | Pelagodinium sp. | 1217 | Akashiwo sp. | 2.3 | Heterocapsa sp. | |
| 0.3 | Azadinium sp. | 1.2 | Heterocapsa triquetra | 4.9 | Peridiniopsis sp. | 1.7 | Peridinium sp. | |
| 0.3 | Akashiwo sp. | 0.9 | Azadinium sp. | 4.0 | Gymnodinium catenatum | 1.5 | Azadinium sp. | |
| 0.2 | Cochlodinium sp. | 0.4 | Akashiwo sp. | 3.7 | Azadinium sp. | 1.1 | Noctiluca scintillans | |
| 0.2 | Pelagodinium sp. | 0.4 | Symbiodinium sp. | 2.6 | Heterocapsa circularisquama | 1.0 | Peridiniopsis sp. | |
| Location | March | June | September | December | ||||
|---|---|---|---|---|---|---|---|---|
| Cell/mL | Species | Cell/mL | Species | Cell/mL | Species | Cell/mL | Species | |
| Gunsan | 2.7 | Heterocapsa triquetra | 2.7 | Scrippsiella sp. | 0.8 | Gyrodinium sp. | 2.9 | Gymnodinium sp. |
| 0.9 | Gyrodinium sp. | 2.5 | Ceratium fusus | 0.1 | Peridiniopsis sp. | 2.9 | Gyrodinium sp. | |
| 0.9 | Prorocentrum micans | 2.5 | Heterocapsa rotundata | 0.1 | Protoperidinium divergence | |||
| 0.9 | Pyrocystis lunula | 1.8 | Gonyaulax sp. | |||||
| 1.2 | Prorocentrum sp. | |||||||
| 0.9 | Dissodinium pseudolunula | |||||||
| 0.6 | Ceratium sp. | |||||||
| 0.6 | Ceratium tripos | |||||||
| 0.6 | Karlodinium sp. | |||||||
| 0.6 | Prorocentrum micans | |||||||
| Pohang | 3.6 | Gymnodinium sp. | 77.8 | Heterocapsa rotundata | 1.7 | Heterocapsa rotundata | 0.4 | Gymnodinium sp. |
| 1.8 | Gyrodinium sp. | 12.1 | Heterocapsa triquetra | 1.7 | Scrippsiella sp. | |||
| 1.4 | Ceratium kofoidii | 7.8 | Gymnodinium sp. | 0.8 | Prorocentrum triestinum | |||
| 1.2 | Alexandrium sp. | 4.3 | Protopeidinium pyriforme | 0.8 | Gymnodinium sp. | |||
| 0.5 | Heterocapsa rotundata | 2.6 | Gyrodinium sp. | 0.8 | Gyrodinium sp. | |||
| 2.6 | Ceratium kofoidii | |||||||
| 1.7 | Alexandrium sp. | |||||||
| 0.9 | Amphidinium operculatum | |||||||
| Tongyeong | 1.6 | *Small thecated dinoflagellate | 341.1 | Prorocentrum triestinum | 2034 | Cochlodinium polykrikoides | 1.6 | Gymnodinium sp. |
| 0.7 | *Small naked dinoflagellate | 18.0 | *Small naked dinoflagellate | 28.8 | Karlodinium sp. | |||
| 0.1 | Alexandrium sp. | 17.0 | *Small thecated dinoflagellate | 18.0 | Gyrodinium sp. | |||
| 0.1 | Gymnodinium sp. | 14.9 | Scrippsiella sp. | 5.4 | Prorocentrum sp. | |||
| 0.1 | Karlodinium sp. | 11.7 | Peridinium sp. | 3.6 | Bysmatrum sp. | |||
| 10.6 | Alexandrium sp. | 3.6 | Ceratium sp. | |||||
| 3.2 | Heterocapsa sp. | 1.8 | Alexandrium sp. | |||||
| 3.2 | Scrippsiella trochoidea | 1.8 | Heterocapsa sp. | |||||
| 2.1 | Protoperidinium sp. | |||||||
| 1.1 | Gonyaulax sp. | |||||||
| 1.1 | Gymnodinium sp. | |||||||
| Seongsan | 0.6 | *Small naked dinoflagellate | 5.8 | Azadinium sp. | 0.6 | *Small naked dinoflagellate | 1.3 | Bysmatrum sp. |
| 0.2 | Bysmatrum sp. | 5.8 | Bysmatrum sp. | 0.5 | Peridiniopsis sp. | 1.0 | Gymnodinium sp. | |
| 0.2 | Katodinium sp. | 5.4 | *Small naked dinoflagellate | 0.3 | Gymnodinium sp. | 0.3 | Gyrodinium sp. | |
| 0.2 | Prorocentrum sp. | 2.4 | *Small thecated dinoflagellate | 0.3 | Prorocentrum minimum | |||
| 1.4 | Gymnodinium sp. | |||||||
| 1.0 | Protoperidinium pellucidum | |||||||
| 0.3 | Heterocapsa sp. | |||||||
| 0.3 | Peridiniopsis sp. | |||||||
| 0.3 | Prorocentrum sp. | |||||||
| 0.3 | Protoperidinium sp. | |||||||
| 0.3 | Woloszynskia sp. | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, J.; Kang, H.W.; Moon, S.J.; Hyung, J.-H.; Lee, E.S.; Park, J. Metagenomic Analysis of the Species Composition and Seasonal Distribution of Marine Dinoflagellate Communities in Four Korean Coastal Regions. Microorganisms 2022, 10, 1459. https://doi.org/10.3390/microorganisms10071459
Hwang J, Kang HW, Moon SJ, Hyung J-H, Lee ES, Park J. Metagenomic Analysis of the Species Composition and Seasonal Distribution of Marine Dinoflagellate Communities in Four Korean Coastal Regions. Microorganisms. 2022; 10(7):1459. https://doi.org/10.3390/microorganisms10071459
Chicago/Turabian StyleHwang, Jinik, Hee Woong Kang, Seung Joo Moon, Jun-Ho Hyung, Eun Sun Lee, and Jaeyeon Park. 2022. "Metagenomic Analysis of the Species Composition and Seasonal Distribution of Marine Dinoflagellate Communities in Four Korean Coastal Regions" Microorganisms 10, no. 7: 1459. https://doi.org/10.3390/microorganisms10071459