
Antibiotic Resistance Properties among Pseudomonas spp. Associated with Salmon Processing Environments


Abstract
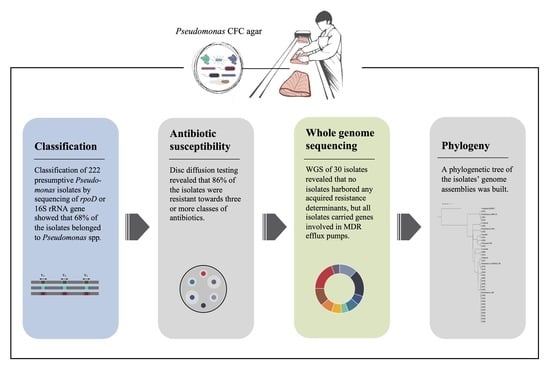
:
1. Introduction
2. Materials and Methods
2.1. Sampling and Preparation of Isolates
2.2. Classification of Presumptive Pseudomonas Isolates by Sequencing of rpoD Gene or 16S rRNA Gene
2.3. Antibiotic Susceptibility by Disc Diffusion Assay
2.4. Whole Genome Sequencing
2.5. Data Analysis of Sequences
3. Results
3.1. Species Relation and Diversity on Pseudomonas CFC Agar
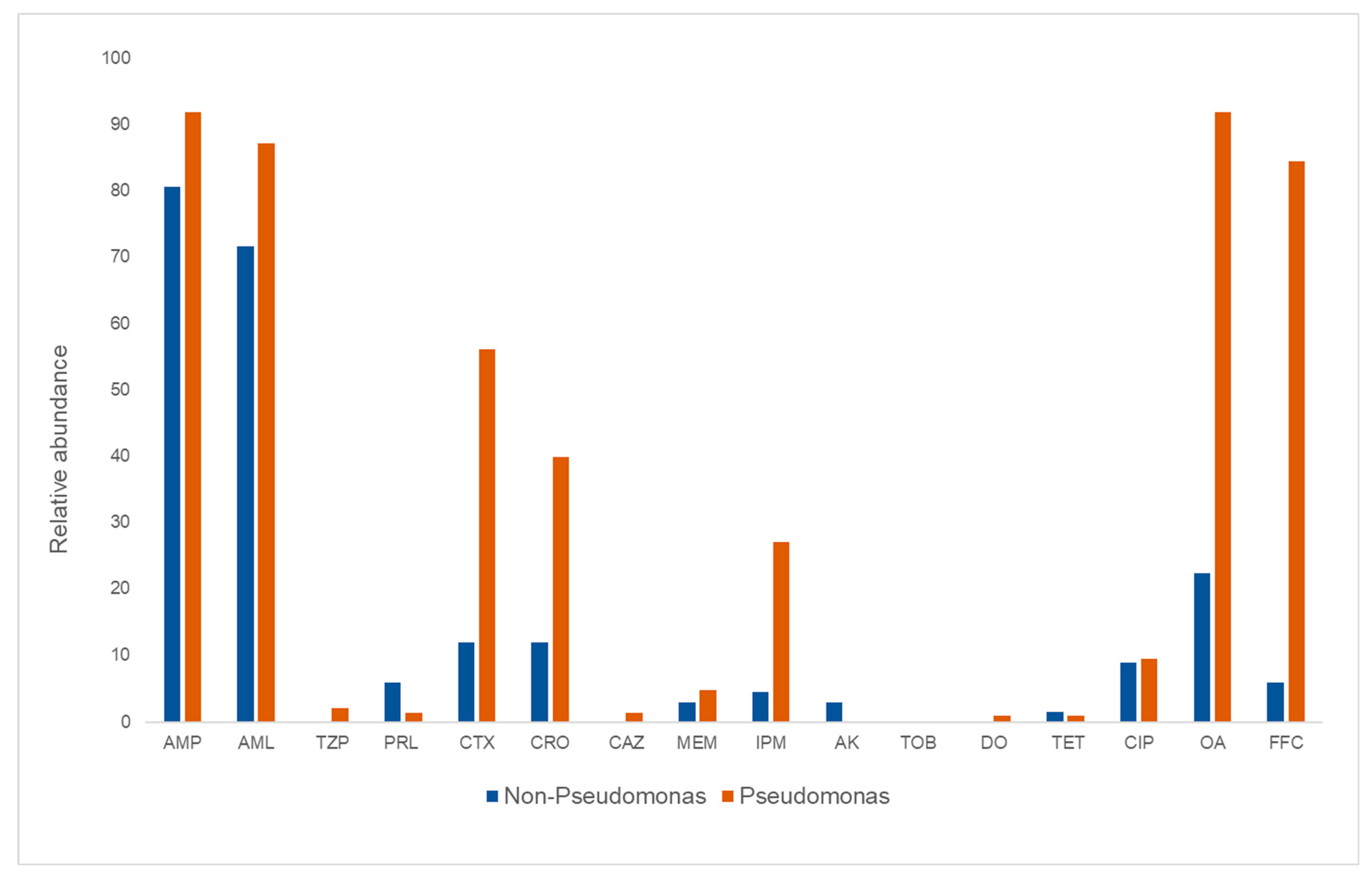
3.2. Phenotypic Antimicrobial Susceptibility
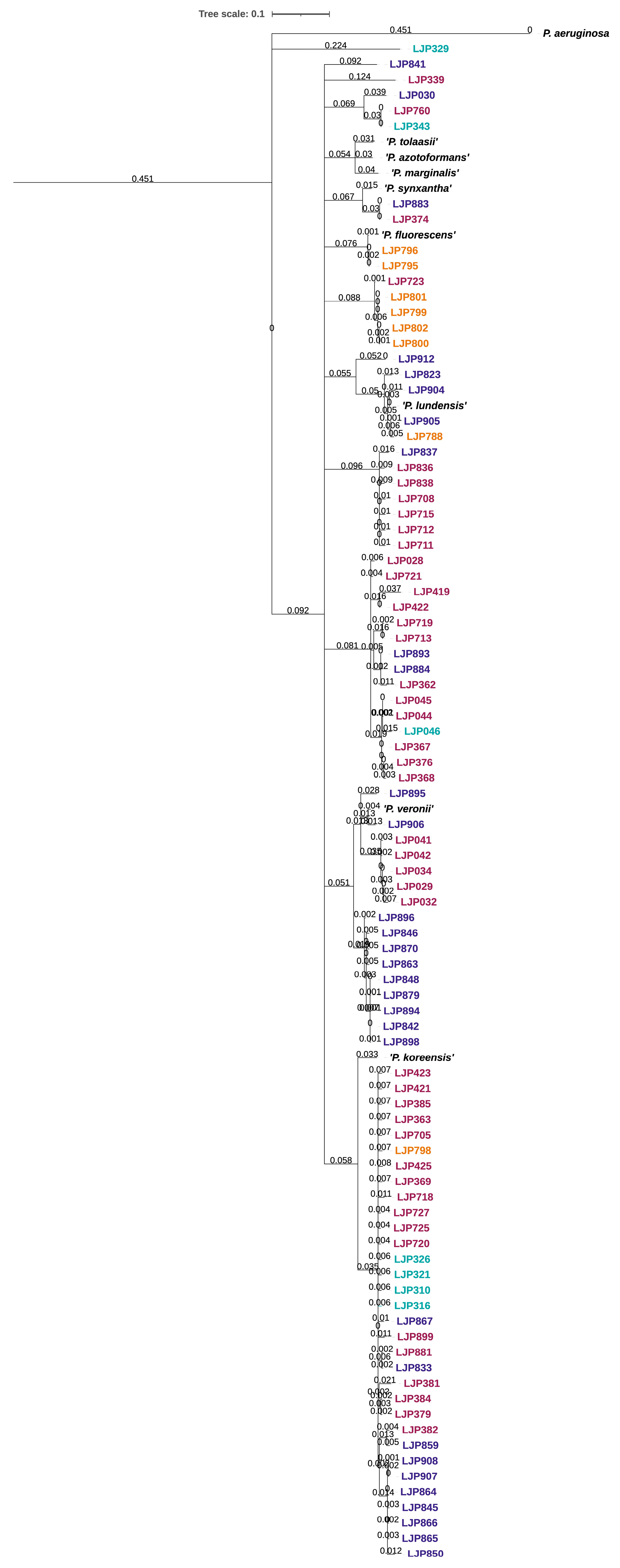
3.3. Genomic Characterization Based on WGS Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- ECDC. Surveillance of Antimicrobial Resistance in Europe 2018; ECDC: Stockholm, Sweden, 2019.
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations; Wellcome Collection: London, UK, 2014. [Google Scholar]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Capita, R.; Alonso-Calleja, C. Antibiotic-resistant bacteria: A challenge for the food industry. Crit. Rev. Food Sci. Nutr. 2013, 53, 11–48. [Google Scholar] [CrossRef] [PubMed]
- WHO. Joint FAO/WHO Expert Meeting in Collaboration with OIE on Foodborne Antimicrobial Resistance: Role of the Environment, Crops and Biocides—Meeting Report; WHO: Roma, Italy, 2019. [Google Scholar]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in aquaculture. Sci. Rep. 2020, 10, 21878. [Google Scholar] [CrossRef] [PubMed]
- NORM/NORM-VET 2020: Usage of Antimicrobial Agents and Occurence of Antimicrobial Resistance in Norway; Norwegian Institute of Public Health: Tromsø, Norway; Oslo, Norway, 2021.
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current status of the use of antibiotics and the antimicrobial resistance in the Chilean salmon farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [Green Version]
- Parmley, J.; Leung, Z.; Léger, D.; Finley, R.; Irwin, R.; Pintar, K.; Engelhardt, R. One Health and Food Safety—The Canadian Experience: A Holistic Approach toward Enteric Bacterial Pathogens and Antimicrobial Resistance Surveillance; National Academic Press: Washington, DC, USA, 2012. [Google Scholar]
- Sanseverino, I.; Navarro Cuenca, A.; Loos, R.; Marinov, D.; Lettieri, T. State of the Art on the Contribution of Water to Antimicrobial Resistance; EUR 29592 EN; Publications Office of the European Union: Luxembourg, 2018. [CrossRef]
- Hudson, J.A.; Frewer, L.J.; Jones, G.; Brereton, P.A.; Whittingham, M.J.; Stewart, G. The agri-food chain and antimicrobial resistance: A review. Trends Food Sci. Technol. 2017, 69, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Brunton, L.A.; Desbois, A.P.; Garza, M.; Wieland, B.; Mohan, C.V.; Häsler, B.; Tam, C.C.; Le, P.N.T.; Phuong, N.T.; Van, P.T.; et al. Identifying hotspots for antibiotic resistance emergence and selection, and elucidating pathways to human exposure: Application of a systems-thinking approach to aquaculture systems. Sci. Total Environ. 2019, 687, 1344–1356. [Google Scholar] [CrossRef]
- Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [Green Version]
- Sørum, H. Antimicrobial Drug Resistance in Fish Pathogens. In Antimicrobial Resistance in Bacteria of Animal Origin; ASM Press: Washington, DC, USA, 2005; pp. 213–238. [Google Scholar]
- Heir, E.; Moen, B.; Åsli, A.W.; Sunde, M.; Langsrud, S. Antibiotic resistance and phylogeny of Pseudomonas spp. isolated over three decades from chicken meat in the Norwegian food chain. Microorganisms 2021, 9, 207. [Google Scholar] [CrossRef]
- Lee, H.-J.; Hoel, S.; Lunestad, B.-T.; Lerfall, J.; Jakobsen, A.N. Aeromonas spp. isolated from ready-to-eat seafood on the Norwegian market: Prevalence, putative virulence factors and antimicrobial resistance. J. Appl. Microbiol. 2021, 130, 1380–1393. [Google Scholar] [CrossRef]
- Delannoy, S.; Hoffer, C.; Youf, R.; Dauvergne, E.; Webb, H.E.; Brauge, T.; Tran, M.-L.; Midelet, G.; Granier, S.A.; Haenni, M.; et al. High Throughput Screening of Antimicrobial Resistance Genes in Gram-Negative Seafood Bacteria. Microorganisms 2022, 10, 1225. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Moen, B.; Heir, E.; Hansen, A.Å.; Langsrud, S. Contamination of salmon fillets and processing plants with spoilage bacteria. Int. J. Food Microbiol. 2016, 237, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomassen, G.M.B.; Krych, Ł.; Knøchel, S.; Mehli, L. Bacterial community development and diversity during the first year of production in a new salmon processing plant. Food Microbiol. 2022, submitted.
- Stellato, G.; Utter, D.R.; Voorhis, A.; De Angelis, M.; Eren, A.M.; Ercolini, D. A few Pseudomonas oligotypes dominate in the meat and dairy processing environment. Front. Microbiol. 2017, 8, 264. [Google Scholar] [CrossRef] [Green Version]
- Peter, S.; Oberhettinger, P.; Schuele, L.; Dinkelacker, A.; Vogel, W.; Dörfel, D.; Bezdan, D.; Ossowski, S.; Marschal, M.; Liese, J.; et al. Genomic characterisation of clinical and environmental Pseudomonas putida group strains and determination of their role in the transfer of antimicrobial resistance genes to Pseudomonas aeruginosa. BMC Genom. 2017, 18, 859. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Park, S.-H.; Park, H.B.; Park, K.-H.; Kim, S.-H.; Jung, S.-I.; Shin, J.-H.; Jang, H.-C.; Kang, S.J. Nosocomial Pseudomonas putida Bacteremia: High Rates of Carbapenem Resistance and Mortality. Chonnam. Med. J. 2012, 48, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Mulet, M.; Bennasar, A.; Lalucat, J.; García-Valdés, E. An rpoD-based PCR procedure for the identification of Pseudomonas species and for their detection in environmental samples. Mol. Cell. Probes 2009, 23, 140–147. [Google Scholar] [CrossRef]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing. PLoS Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating deep phylogenetic relationships among Cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- EUCAST. EUCAST Disk Diffusion Method for Antimicrobial Susceptibility Testing Version 9.0. 2021. Available online: https://www.eucast.org/ast_of_bacteria/disk_diffusion_methodology/ (accessed on 1 October 2021).
- EUCAST. EUCAST Clinical Breakpoint Tables V.12.0. 2022. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 15 March 2022).
- Miranda, C.D.; Smith, P.; Rojas, R.; Contreras-Lynch, S.; Vega, J.M.A. Antimicrobial Susceptibility of Flavobacterium psychrophilum from Chilean Salmon Farms and Their Epidemiological Cut-Off Values Using Agar Dilution and Disk Diffusion Methods. Front. Microbiol. 2016, 7, 1880. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.; Larvor, E.; Jouy, E.; Kempf, I.; Le Bouquin, S.; Chauvin, C.; Boitard, P.-M.; Jamin, M.; Le Breton, A.; Thuillier, B.; et al. Agreement between the categorization of isolates of Aeromonas salmonicida and Yersinia ruckeri by disc diffusion and MIC tests performed at 22 °C. J. Fish Dis. 2021, 44, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasman, H.; Saputra, D.; Sicheritz-Pontén, T.; Lund, O.; Svendsen, C.A.; Frimodt-Møller, N.; Aarestrup, F.M. Rapid Whole-Genome Sequencing for detection and Characterization of Microorganisms Directly from Clinical Samples. J. Clin. Microbiol. 2014, 52, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529. [Google Scholar] [CrossRef] [Green Version]
- Clausen, P.T.L.C.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Leekitcharoenphon, P.; Nielsen, E.M.; Kaas, R.S.; Lund, O.; Aarestrup, F.M. Evaluation of whole genome sequencing for outbreak detection of Salmonella enterica. PLoS ONE 2014, 9, e87991. [Google Scholar] [CrossRef] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in bacterial taxonomy: Impact on the genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, L.; Lood, C.; Höfte, M.; Vandamme, P.; Rokni-Zadeh, H.; van Noort, V.; Lavigne, R.; De Mot, R. The Ever-Expanding Pseudomonas Genus: Description of 43 New Species and Partition of the Pseudomonas putida Group. Microorganisms 2021, 9, 1766. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.; Lood, C.; Rokni-Zadeh, H.; van Noort, V.; Lavigne, R.; De Mot, R. Reliable identification of Environmental Pseudomonas isolates using the rpoD gene. Microorganisms 2020, 8, 1166. [Google Scholar] [CrossRef]
- Tryfinopoulou, P.; Drosinos, E.H.; Nychas, G.J.E. Performance of Pseudomonas CFC-selective medium in the fish storage ecosystems. J. Microbiol. Methods 2001, 47, 243–247. [Google Scholar] [CrossRef]
- Mulet, M.; Lalucat, J.; García-Valdés, E. DNA sequence-based analysis of the Pseudomonas species. Environ. Microbiol. 2010, 12, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Gomila, M.; Peña, A.; Mulet, M.; Lalucat, J.; García-Valdés, E. Phylogenomics and systematics in Pseudomonas. Front. Microbiol. 2015, 6, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midtlyng, P.J.; Grave, K.; Horsberg, T.E. What has been done to minimize the use of antibacterial and antiparasitic drugs in Norwegian aquaculture? Aquac. Res. 2011, 42, 28–34. [Google Scholar] [CrossRef]
- Hancock, R.E.; Speert, D.P. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and impact on treatment. Drug Resist. Updat. 2000, 3, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.J. Antimicrobial Resistance. In Sherris Medical Microbiology—An Introduction to Infectious Diseases, 4th ed.; Ryan, K.J., Ray, C.G., Eds.; McGraw Hill, Medical Publishing Division: New York, NY, USA, 2000. [Google Scholar]
- Meng, L.; Liu, H.; Lan, T.; Dong, L.; Hu, H.; Zhao, S.; Zhang, Y.; Zheng, N.; Wang, J. Antibiotic resistance patterns of Pseudomonas spp. isolated from raw milk revealed by whole genome sequencing. Front. Microbiol. 2020, 11, 1005. [Google Scholar] [CrossRef] [PubMed]
- Decimo, M.; Silvetti, T.; Brasca, M. Antibiotic Resistance Patterns of Gram-Negative Psychrotrophic Bacteria from Bulk Tank Milk. J. Food Sci. 2016, 81, M944–M951. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, C.; Gutiérrez, M.S.; Venegas, L.; Sapag, C.; Araya, C.; Caruffo, M.; López, P.; Reyes-Jara, A.; Toro, M.; González-Rocha, G.; et al. Microbiota composition and susceptibility to florfenicol and oxytetracycline of bacterial isolates from mussels (Mytilus spp.) reared on different years and distance from salmon farms. Environ. Res. 2022, 204, 112068. [Google Scholar] [CrossRef]
- Miranda, C.D.; Rojas, R. Occurrence of florfenicol resistance in bacteria associated with two Chilean salmon farms with different history of antibacterial usage. Aquaculture 2007, 266, 39–46. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Tomova, A.; López, A.; Maldonado, M.A.; Henríquez, L.A.; Ivanova, L.; Moy, F.; Godfrey, H.P.; Cabello, F.C. Salmon aquaculture and antimicrobial resistance in the marine environment. PLoS ONE 2012, 7, e42724. [Google Scholar] [CrossRef]
- Fernández-Alarcón, C.; Miranda, C.D.; Singer, R.S.; López, Y.; Rojas, R.; Bello, H.; Domínguez, M.; González-Rocha, G. Detection of the floR Gene in a Diversity of Florfenicol Resistant Gram-Negative Bacilli from Freshwater Salmon Farms in Chile. Zoonoses Public Health 2010, 57, 181–188. [Google Scholar] [CrossRef]
- Adesoji, A.T.; Call, D.R. Molecular analysis of florfenicol-resistant bacteria isolated from drinking water distribution systems in Southwestern Nigeria. J. Glob. Antimicrob. Resist. 2020, 23, 340–344. [Google Scholar] [CrossRef]
- Umasuthan, N.; Valderrama, K.; Vasquez, I.; Segovia, C.; Hossain, A.; Cao, T.; Gnanagobal, H.; Monk, J.; Boyce, D.; Santander, J. A Novel Marine Pathogen Isolated from Wild Cunners (Tautogolabrus adspersus): Comparative Genomics and Transcriptome Profiling of Pseudomonas sp. Strain J380. Microorganisms 2021, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Kverme, K.O.; Haugland, G.T.; Hannisdal, R.; Kallekleiv, M.; Colquhoun, D.J.; Lunestad, B.T.; Wergeland, H.I.; Samuelsen, O.B. Pharmacokinetics of florfenicol in lumpfish (Cyclopterus lumpus L.) after a single oral administration. Aquaculture 2019, 512, 734279. [Google Scholar] [CrossRef]
- Grave, K.; Helgesen, K.O. Antibakterielle Midler til Oppdrettsfisk—Rekvirering, Forbruk og Diagnoser 2013–2017 (Norwegian); Norwegian Veterinary Institute: Oslo, Norway, 2018. [Google Scholar]
- Kerry, J.; Hiney, M.; Coyne, R.; Cazabon, D.; NicGabhainn, S.; Smith, P. Frequency and distribution of resistance to oxytetracycline in micro-organisms isolated from marine fish farm sediments following therapeutic use of oxytetracycline. Aquaculture 1994, 123, 43–54. [Google Scholar] [CrossRef]
- Sengeløv, G.; Agersø, Y.; Halling-Sørensen, B.; Baloda, S.B.; Andersen, J.S.; Jensen, L.B. Bacterial antibiotic resistance levels in Danish farmland as a result of treatment with pig manure slurry. Environ. Int. 2003, 28, 587–595. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Tacão, M.; Correia, A.; Henriques, I.S. Low Prevalence of Carbapenem-Resistant Bacteria in River Water: Resistance Is Mostly Related to Intrinsic Mechanisms. Microb. Drug Resist. 2015, 21, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Goli, H.R.; Nahaei, M.R.; Rezaee, M.A.; Hasani, A.; Kafil, H.S.; Aghazadeh, M.; Nikbakht, M.; Khalili, Y. Role of MexAB-OprM and MexXY-OprM efflux pumps and class 1 integrons in resistance to antibiotics in burn and Intensive Care Unit isolates of Pseudomonas aeruginosa. J. Infect. Public Health 2018, 11, 364–372. [Google Scholar] [CrossRef]
- Breidenstein, E.B.; de la Fuente-Núñez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Kakoullis, L.; Papachristodoulou, E.; Chra, P.; Panos, G. Mechanisms of Antibiotic Resistance in Important Gram-Positive and Gram-Negative Pathogens and Novel Antibiotic Solutions. Antibiotics 2021, 10, 415. [Google Scholar] [CrossRef]
- Coyne, S.; Rosenfeld, N.; Lambert, T.; Courvalin, P.; Périchon, B. Overexpression of resistance-nodulation-cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4389–4393. [Google Scholar] [CrossRef] [Green Version]
- Palma, M.; Zurita, J.; Ferreras, J.A.; Worgall, S.; Larone, D.H.; Shi, L.; Campagne, F.; Quadri, L.E.N. Pseudomonas aeruginosa SoxR does not conform to the archetypal paradigm for SoxR-dependent regulation of the bacterial oxidative stress adaptive response. Infect. Immun. 2005, 73, 2958–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Varela, M.; Corral, J.; Aranda, J.; Barbé, J. Functional Characterization of AbaQ, a Novel Efflux Pump Mediating Quinolone Resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2018, 62, e00906–e00918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.; Kronvall, G. Effect of incubation temperature and time on the precision of data generated by antibiotic disc diffusion assays. J. Fish Dis. 2015, 38, 629–636. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
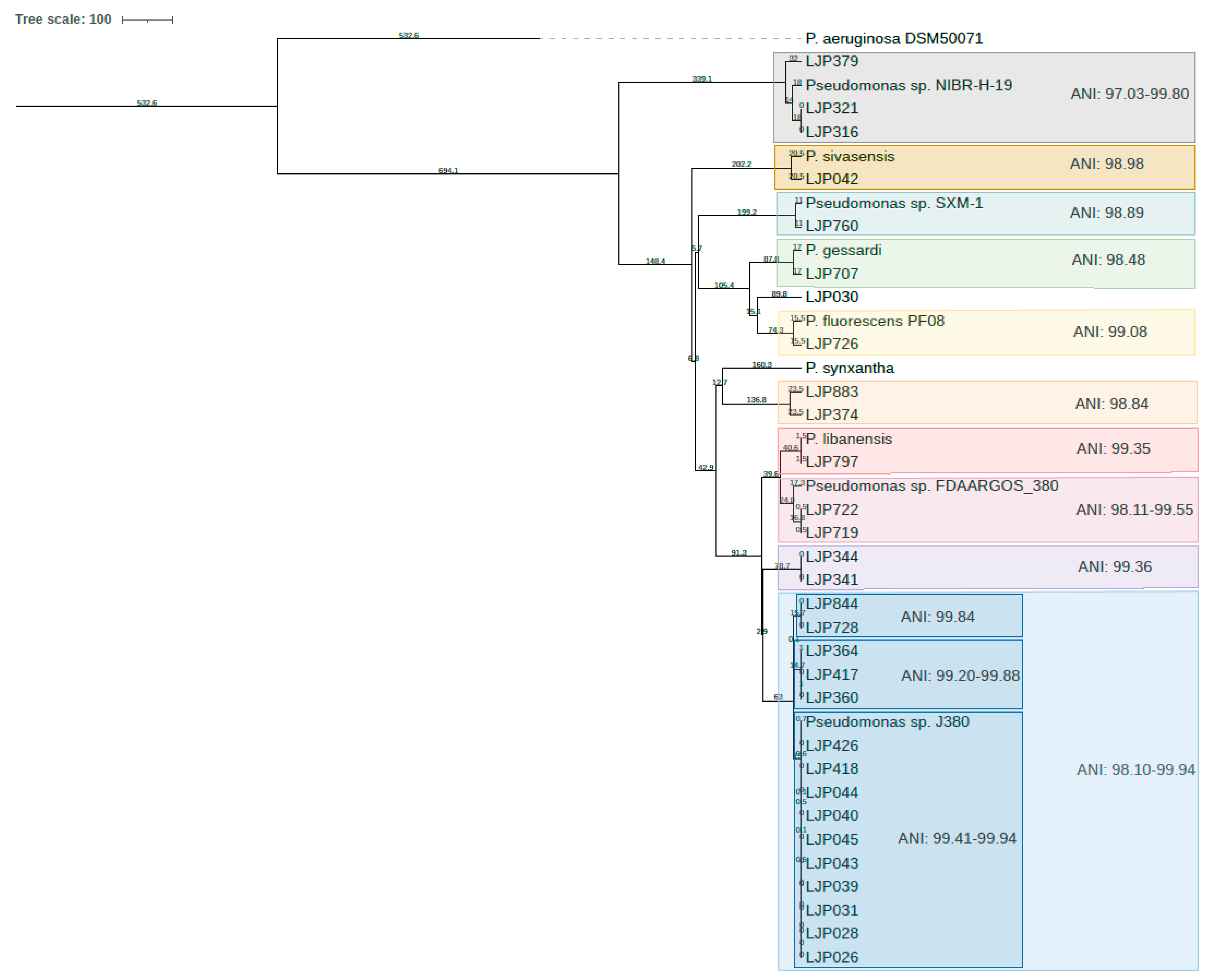{kind=link}
| Sampling Point | Sampling Point Category | Sampling Type | Sampling Area |
|---|---|---|---|
| Inlet water | Contact surface, slaughter | Water | 100 mL |
| Drain slaughter dep. | Non-contact | Cloth | 30 cm × 30 cm |
| Conveyor slaughter dep. | Contact, slaughter | Cloth | 30 cm × 30 cm |
| Gutting machine, suction | Contact, slaughter | Swab | 10 cm × 10 cm |
| Head cutter knife | Contact, slaughter | Cloth | 2 cm × Ø25 cm |
| Conveyor fillet dep. | Contact, fillet | Cloth | 30 cm × 30 cm |
| Drain fillet dep. | Non-contact | Cloth | 30 cm × 30 cm |
| Fish fillet before packaging | Fish | Fish fillet | 25 g |
| Skin, gutted whole fish | Fish | Swab | 10 cm × 10 cm |
| Gills, gutted whole fish | Fish | Swab | Gills on both side of fish |
| Sampl. Time | Sampl. Point | Isolate NO (LJP) | Taxonomic Classification | Resistance Profiles | Res. to # Antib. Classes |
|---|---|---|---|---|---|
| 1 | CSL | 035, 040 | Pseuddomonas fluorescens (2) | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 1 | CSL | 044 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, CAZ, IPM, OA, FFC | 5 |
| 1 | CSL | 045 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, CIP, OA, FFC | 4 |
| 1 | CSL | 042 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 1 | CSL | 033, 038 | Pseudomonas tolaasii (2) | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 1 | CSL | 037 | Pseudomonas umsongensis | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 1 | CSL | 043 | Pseudomonas synxantha | AMP, AML, CTX, CRO, IPM, CIP, OA, FFC | 5 |
| 1 | CSL | 046 | Pseudomonas sp. | AMP, AML, CTX, IPM, OA, FFC | 5 |
| 1 | CSL | 032, 034, 041 | Unclassified Pseudomonas (3) | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 1 | CSL | 039 | Pseudomonas cedrina | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 1 | G | 028 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, IPM, CIP, DO, OA, FFC | 6 |
| 1 | G | 030 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, MEM, IPM, OA, FFC | 5 |
| 1 | G | 027 | Pseudomonas reactans | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 1 | G | 029 | Unclassified Pseudomonas | AMP, AML, IPM, OA, FFC | 4 |
| 1 | G | 026 | Pseudomonas azotoformans | AMP, AML, CTX, CRO, IPM, CIP, OA, FFC | 5 |
| 1 | G | 031 | Pseudomonas cedrina | AMP, AML, CTX, IPM, OA, FFC | 5 |
| 1 | IW | 009 | Pseudomonas brenneri | susceptible | 0 |
| 2 | IW | 310, 326 | Pseudomonas fluorescens (2) | AMP, AML, OA, FFC | 3 |
| 2 | IW | 316 | Pseudomonas fluorescens | AMP, AML, CIP, OA, FFC | 3 |
| 2 | IW | 321 | Pseudomonas fluorescens | AMP, AML, CTX, OA, FFC | 4 |
| 2 | IW | 314, 315 | Pseudomonas guineae (2) | OA | 1 |
| 2 | IW | 309 | Pseudomonas marincola | OA, FFC | 2 |
| 2 | IW | 312 | Pseudomonas pseudoalcaligenes | AMP | 1 |
| 2 | IW | 313, 320 | Unclassified Pseudomonas (2) | OA, FFC | 2 |
| 2 | IW | 311 | Unclassified Pseudomonas | AMP | 1 |
| 2 | IW | 327, 329 | Unclassified Pseudomonas (2) | OA | 1 |
| 2 | CSL | 339 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 2 | G | 366, 371 | Pseudomonas azotoformans (2) | AMP, AML, IPM, OA, FFC | 4 |
| 2 | G | 375 | Pseudomonas azotoformans | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 2 | G | 360 | Pseudomonas cedrina | AMP, AML, CTX, IPM, OA, FFC | 5 |
| 2 | G | 362 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 5 |
| 2 | G | 383 | Pseudomonas fluorescens | AMP, AML, CTX, OA, FFC | 4 |
| 2 | G | 374 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, CAZ, CIP, OA, FFC | 4 |
| 2 | G | 364 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 2 | G | 370 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 2 | G | 373a | Pseudomonas fluorescens | AMP, AML, IPM, OA, FFC | 4 |
| 2 | G | 369 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 2 | G | 363, 381, 382, 384, 385 | Pseudomonas fluorescens (5) | AMP, AML, OA, FFC | 3 |
| 2 | G | 379 | Pseudomonas fluorescens | AMP, AML, TET, OA, FFC | 4 |
| 2 | G | 365, 372 | Unclassified Pseudomonas | AMP, AML, OA, FFC | 3 |
| 2 | G | 367 | Unclassified Pseudomonas | AMP, AML, CTX, IPM, OA, FFC | 5 |
| 2 | G | 368, 376 | Unclassified Pseudomonas (2) | AMP, AML, CRO, IPM, OA, FFC | 5 |
| 2 | G | 380 | Unclassified Pseudomonas | AMP, AML, CRO, IPM, CIP, OA, FFC | 5 |
| 2 | HCK | 421 | Pseudomoans lurida | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 2 | HCK | 422 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 2 | HCK | 419, 423, 425 | Pseudomonas fluorescens (3) | AMP, AML, OA, FFC | 3 |
| 2 | HCK | 417, 426 | Pseudomonas fluorescens (2) | AMP, AML, CTX, CRO, IPM, CIP, OA, FFC | 5 |
| 2 | HCK | 418 | Pseudomonas marginalis | AMP, AML, CTX, CRO, IPM, CIP, OA, FFC | 5 |
| 2 | CSK | 344 | Pseudomonas azotoformans | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 2 | CSK | 341 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 2 | CSK | 343 | Unclassified Pseudomonas | AMP, AML, CRO, OA, FFC | 4 |
| 3 | DS | 710 | Pseudomonas azotoformans | AMP, AML, CTX, OA, FFC | 4 |
| 3 | DS | 713 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | DS | 705 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | DS | 706 | Pseudomonas gessardii | AMP, AML, TZP, CTX, CRO, MEM, OA, FFC | 5 |
| 3 | DS | 707 | Pseudomonas gessardii | AMP, AML, TZP, PRL, CTX, CRO, MEM, OA, FFC | 5 |
| 3 | DS | 714 | Pseudomonas gessardii | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | DS | 716 | Pseudomonas gessardii | AMP, AML, TZP, CTX, CRO, MEM, OA, FFC | 5 |
| 3 | DS | 718 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | DS | 708, 711, 715 | Unclassified Pseudomonas (3) | AMP, AML, CTX, OA, FFC | 4 |
| 3 | DS | 712 | Unclassified Pseudomonas | AMP, AML, CTX, OA, FFC | 4 |
| 3 | DS | 709 | Unclassified Pseudomonas | AMP, AML, OA, FFC | 3 |
| 3 | CSL | 722 | Pseudomonas fluorescens | AMP, AML, CTX, IPM, OA, FFC | 5 |
| 3 | CSL | 721, 719 | Pseudomonas fluorescens (2) | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 3 | CSL | 720, 727 | Pseudomonas fluorescens (2) | AMP, AML, OA, FFC | 3 |
| 3 | CSL | 725 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | CSL | 726 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, MEM, IPM, OA, FFC | 5 |
| 3 | CSL | 728 | Pseudomonas paralactis | AMP, AML, CTX, IPM, CIP, OA, FFC | 5 |
| 3 | CSL | 724 | Pseudomonas poae | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | CSL | 723 | Unclassified Pseudomonas | AMP, AML, CTX, CRO, MEM, IPM, OA, FFC | 5 |
| 3 | HCK | 760 | Pseudomonas putida | AMP, AML, CRO, OA, FFC | 4 |
| 3 | CSK | 788 | Pseudomonas lundensis | AMP | 1 |
| 3 | DF | 796 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | DF | 799, 800, 801 | Unclassified Pseudomonas (3) | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | DF | 802 | Unclassified Pseudomonas | AMP, AML, CTX, OA, FFC | 4 |
| 3 | DF | 795 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | DF | 798 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | DF | 797 | Pseudomonas azotoformans | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 3 | G2 | 823 | Pseudomonas anguilliseptica | susceptible | 0 |
| 3 | S1 | 844 | Pseudomonas azotoformans | AMP, AML, CTX, CRO, IPM, CIP, OA, FFC | 5 |
| 3 | S1 | 840 | Pseudomonas extremaustralis | AMP, AML, CTX, OA, FFC | 4 |
| 3 | S5 | 910 | Pseudomonas extremaustralis | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | S1 | 843 | Pseudomonas extremaustralis | AMP, AML, CTX, OA | 3 |
| 3 | S5 | 899 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | S1 | 836 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | S3 | 867 | Pseudomonas fluorescens | AMP, AML, CIP, OA, FFC | 3 |
| 3 | S4 | 880 | Pseudomonas fluorescens | AMP, AML, CTX, OA, FFC | 4 |
| 3 | S4 | 881 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | S4 | 883 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, IPM, CIP, OA, FFC | 5 |
| 3 | S4 | 889 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, MEM, IPM, OA, FFC | 5 |
| 3 | S5 | 906 | Pseudomonas fluorescens | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | S5 | 907, 908 | Pseudomonas fluorescens (2) | AMP, AML, OA | 2 |
| 3 | S1 | 845 | Pseudomonas fluorescens | AMP, AML, OA, FFC | 3 |
| 3 | S2 | 850 | Pseudomonas fluorescens | AMP | 1 |
| 3 | S2 | 859 | Pseudomonas fluorescens | susceptible | 0 |
| 3 | S3 | 864 | Pseudomonas fluorescens | AMP, AML, OA | 2 |
| 3 | S3 | 865 | Pseudomonas fluorescens | AMP, AML | 1 |
| 3 | S3 | 866 | Pseudomonas fluorescens | AML | 1 |
| 3 | S4 | 887 | Pseudomonas fluorescens | AMP | 1 |
| 3 | S5 | 912 | Pseudomonas fragi | AMP, AML, PRL, CTX, OA, FFC | 4 |
| 3 | S1 | 835 | Pseudomonas gessardii | AMP, AML, CTX, OA, FFC | 4 |
| 3 | S4 | 893 | Pseudomonas libanensis/fluorescens | AMP, AML, OA | 2 |
| 3 | S4 | 882 | Pseudomonas libanesis | AMP, AML, OA, FFC | 3 |
| 3 | S4 | 888 | Pseudomonas libanesis | AMP, AML, CTX, CRO, IPM, OA, FFC | 5 |
| 3 | F2 | 833 | Pseudomonas lundensis | AMP, OA, FFC | 3 |
| 3 | S5 | 905 | Pseudomonas lundensis | AMP, OA, FFC | 3 |
| 3 | S5 | 904 | Pseudomonas lundensis | AMP | 1 |
| 3 | S5 | 895 | Pseudomonas veronii | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | S4 | 884 | Pseudomonas libanensis | AMP, AML, CTX, OA, FFC | 4 |
| 3 | S5 | 896 | Unclassified Pseudomonas | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | S1 | 846 | Unclassified Pseudomonas | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | S3 | 863, 870 | Unclassified Pseudomonas (2) | AMP, AML, CTX, CRO, OA, FFC | 4 |
| 3 | S1 | 838, 839, 842, 837 | Unclassified Pseudomonas (4) | AMP, AML, CTX, OA, FFC | 4 |
| 3 | S1 | 841 | Unclassified Pseudomonas | AMP, AML, OA, FFC | 3 |
| 3 | S2 | 848 | Unclassified Pseudomonas | AMP, AML, CTX, OA, FFC | 4 |
| 3 | S4 | 879 | Unclassified Pseudomonas | AMP, AML, OA, FFC | 3 |
| 3 | S5 | 894 | Unclassified Pseudomonas | susceptible | 0 |
| 3 | S5 | 898, 903 | Unclassified Pseudomonas (2) | AMP, AML, CTX, OA | 3 |
| Isolates (LJP) | Antimicrobial Resistance Determinants | Predicted Antimicrobial Resistance |
|---|---|---|
| 726, 030, 707 | adeF | fluoroquinolone; tetracycline |
| 418 | adeF, soxR | fluoroquinolone; cephalosporin; glycylcycline; penam; tetracycline; rifamycin; phenicol; disinfecting agents and antiseptics |
| 026, 028, 031, 039, 040, 042, 043, 044, 045, 316, 321, 341, 344, 360, 364, 374, 379, 417, 426, 719, 722, 728, 760, 797, 844, 883 | adeF, soxR, AbaQ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomassen, G.M.B.; Reiche, T.; Tennfjord, C.E.; Mehli, L. Antibiotic Resistance Properties among Pseudomonas spp. Associated with Salmon Processing Environments. Microorganisms 2022, 10, 1420. https://doi.org/10.3390/microorganisms10071420
Thomassen GMB, Reiche T, Tennfjord CE, Mehli L. Antibiotic Resistance Properties among Pseudomonas spp. Associated with Salmon Processing Environments. Microorganisms. 2022; 10(7):1420. https://doi.org/10.3390/microorganisms10071420
Chicago/Turabian StyleThomassen, Gunn Merethe Bjørge, Thorben Reiche, Christine Eikås Tennfjord, and Lisbeth Mehli. 2022. "Antibiotic Resistance Properties among Pseudomonas spp. Associated with Salmon Processing Environments" Microorganisms 10, no. 7: 1420. https://doi.org/10.3390/microorganisms10071420