
Shallow-Water Hydrothermal Vents as Natural Accelerators of Bacterial Antibiotic Resistance in Marine Coastal Areas

 , , , ,
, , , , 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
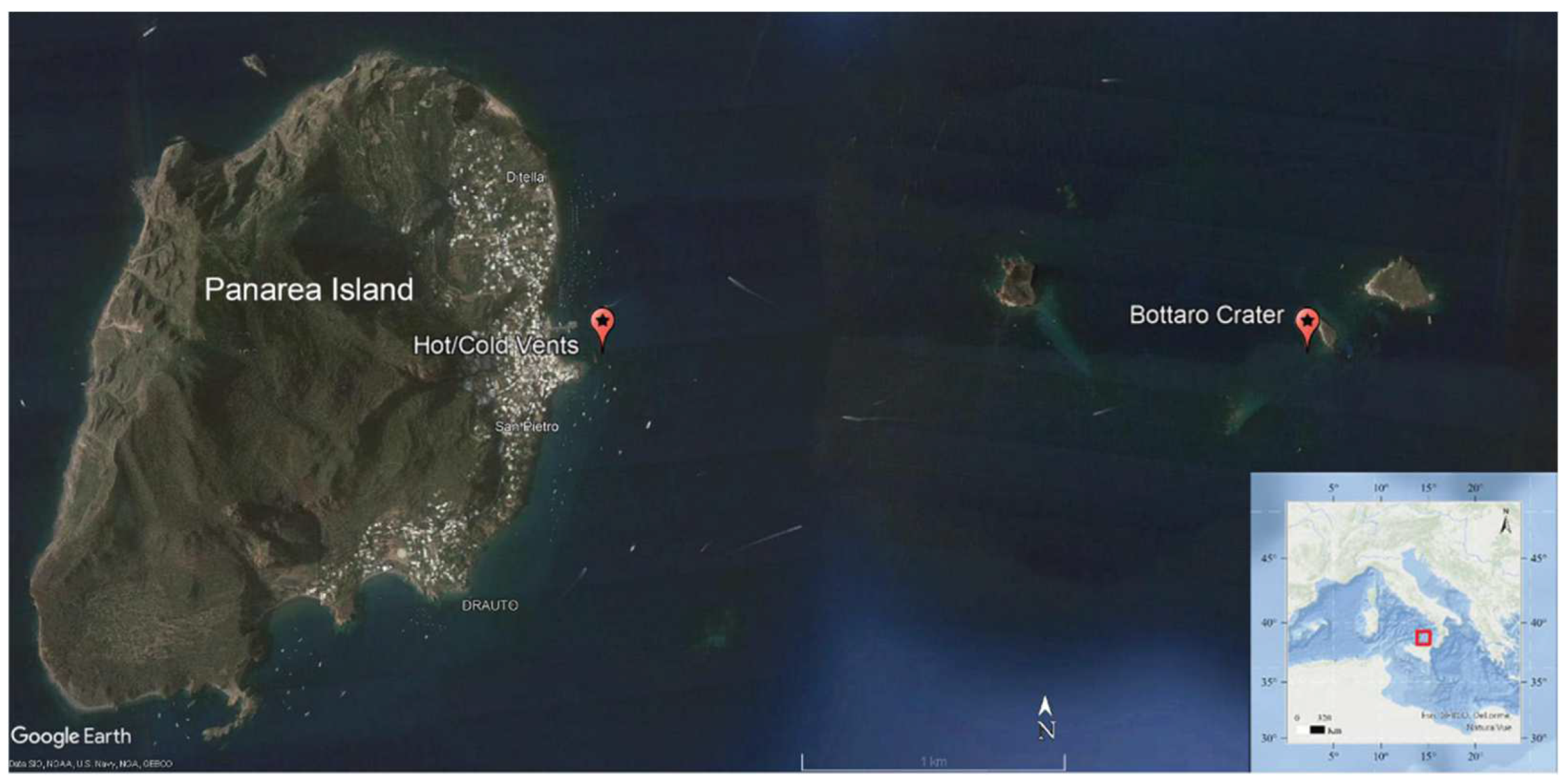
2.1. Sampling Area and Environmental Characterization
2.2. Isolation and Preliminary Characterization of Bacteria from Sediment and Water Samples
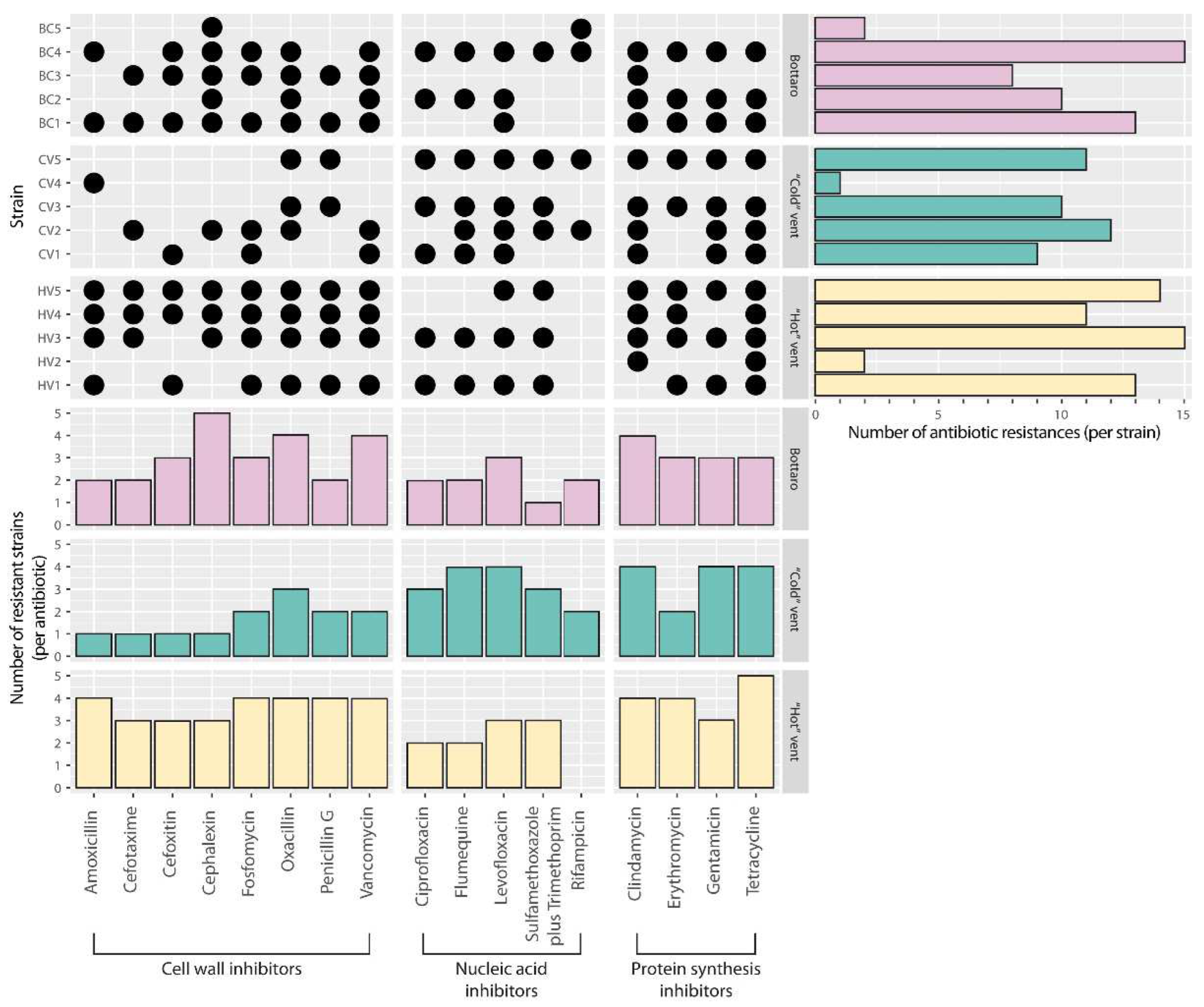
2.3. Antibiotic Susceptibility Tests
2.4. Taxonomic Identification of Bacterial Strains and Screening of AR Genes in Phylogenetically Closest Bacteria
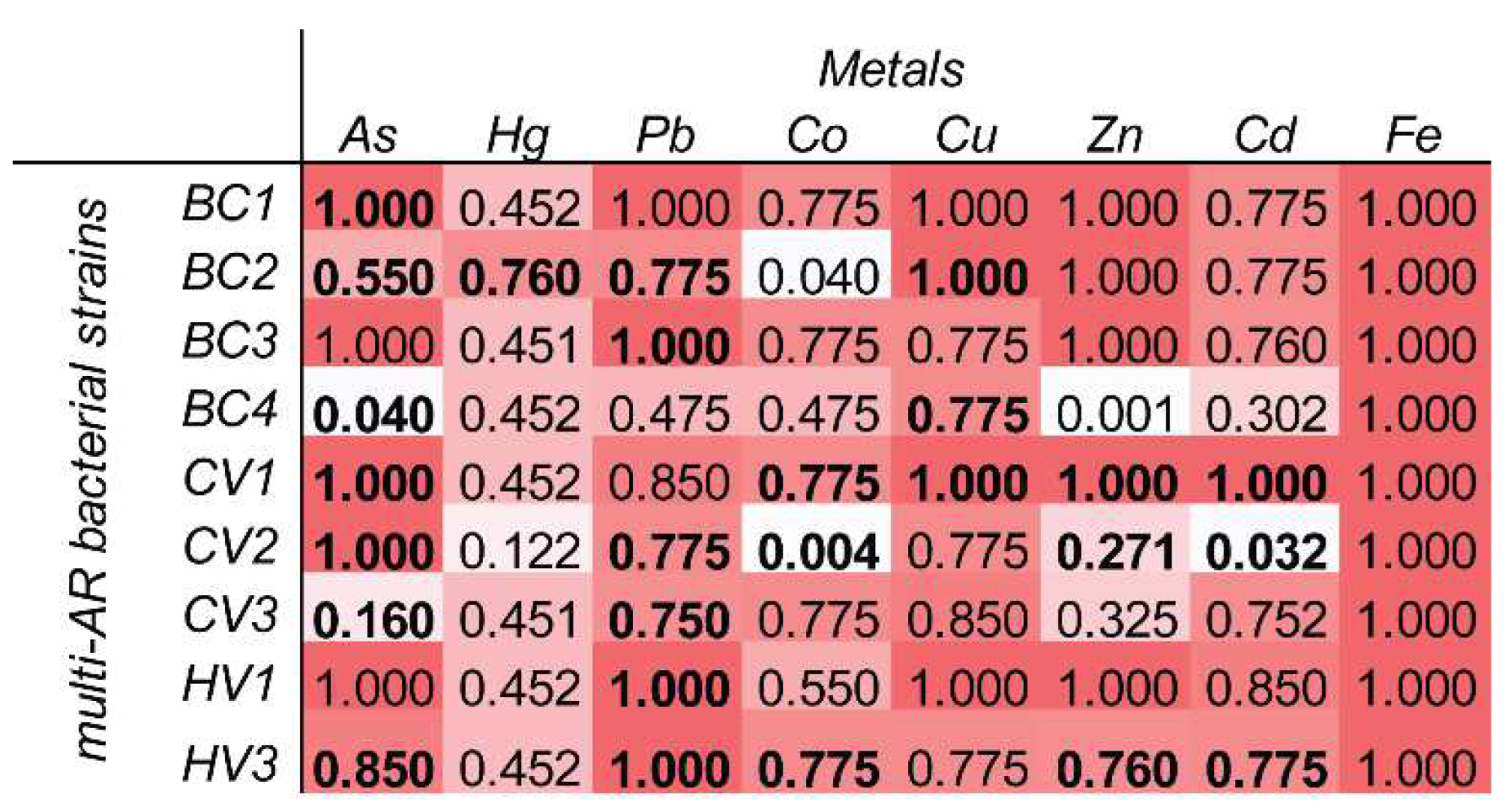
2.5. Multi-AR Isolates’ In Vitro Physiological Tests
2.6. Physiological and Genetic Traits for HM Tolerance in Antibiotic-Resistant Bacterial Strains
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morehead, M.S.; Scarbrough, C. Emergence of global antibiotic resistance. Prim. Care 2018, 45, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, R. In the beginning there was antibiotic resistance. In The End of an Antibiotic Era; Springer: New York, NY, USA, 2021; pp. 15–45. [Google Scholar]
- Allen, H.K.; Moe, L.A.; Rodbumrer, J.; Gaarder, A.; Handelsman, J. Functional metagenomics reveals diverse β-lactamases in a remote Alaskan soil. ISME J. 2009, 3, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The shared antibiotic resistome of soil bacteria and human pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segawa, T.; Takeuchi, N.; Rivera, A.; Yamada, A.; Yoshimura, Y.; Barcaza, G.; Shinbori, K.; Motoyama, H.; Kohshima, S.; Ushida, K. Distribution of antibiotic resistance genes in glacier environments. Environ. Microbiol. Rep. 2013, 5, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.K.; Forsberg, K.J.; Dantas, G. Improved annotation of antibiotic resistance determinants reveals microbial resistomes cluster by ecology. ISME J. 2015, 9, 207–216. [Google Scholar] [CrossRef]
- Miller, R.V.; Gammon, K.; Day, M.J. Antibiotic resistance among bacteria isolated from seawater and penguin fecal samples collected near Palmer Station, Antarctica. Can. J. Microbiol. 2009, 55, 37–45. [Google Scholar] [CrossRef]
- Martiny, A.C.; Martiny, J.B.H.; Weihe, C.; Field, A.; Ellis, J. Functional metagenomics reveals previously unrecognized diversity of antibiotic resistance genes in gulls. Front. Microbiol. 2011, 2, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatosy, S.M.; Martiny, A.C. The ocean as a global reservoir of antibiotic resistance genes. Appl. Environ. Microbiol. 2015, 81, 7593. [Google Scholar] [CrossRef] [Green Version]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.-N. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Dong, P.; Wang, H.; Fang, T.; Wang, Y.; Ye, Q. Assessment of extracellular antibiotic resistance genes (eARGs) in typical environmental samples and the transforming ability of eARG. Environ. Int. 2019, 125, 90–96. [Google Scholar] [CrossRef]
- Ebmeyer, S.; Kristiansson, E.; Larsson, D.G.J. A framework for identifying the recent origins of mobile antibiotic resistance genes. Commun. Biol. 2021, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Komijani, M.; Shamabadi, N.S.; Shahin, K.; Eghbalpour, F.; Tahsili, M.R.; Bahram, M. Heavy metal pollution promotes antibiotic resistance potential in the aquatic environment. Environ. Pollut. 2021, 274, 116569. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.H.; Li, X.; Rashid, A.; Su, J.; Xu, J.; Brejnrod, A.D.; Su, J.-Q.; Wu, Y.; Zhu, Y.-G.; Zhou, S.G. Co-selection of antibiotic resistance genes, and mobile genetic elements in the presence of heavy metals in poultry farm environments. Sci. Total Environ. 2021, 755, 142702. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.-Y.; He, L.-Y.; Gao, F.-Z.; Zhang, M.; Chen, S.; Wu, D.-L.; Liu, Y.-S.; He, L.-X.; Bai, H.; Ying, G.-G. Antibiotic resistance genes in surface water and groundwater from mining affected environments. Sci. Total Environ. 2021, 772, 145516. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, Y.; Liu, L.; Li, L.-Y.; Lin, H.; Wu, X.-Y.; Bi, W.-J.; Wang, L.-T.; Mao, D.-Q.; Luo, Y. The prevalence of ampicillin-resistant opportunistic pathogenic bacteria undergoing selective stress of heavy metal pollutants in the Xiangjiang River, China. Environ. Pollut. 2021, 268, 115362. [Google Scholar] [CrossRef]
- Tauch, A.; Schlüter, A.; Bischoff, N.; Goesmann, A.; Meyer, F.; Pühler, A. The 79,370-bp conjugative plasmid pB4 consists of an IncP-1β backbone loaded with a chromate resistance transposon, the strA-strB streptomycin resistance gene pair, the oxacillinase gene bla NPS-1, and a tripartite antibiotic efflux system of the resistan. Mol. Genet. Genom. 2003, 268, 570–584. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Major uncertainties and future research opportunities in metal–antibiotic co-selection. Trends Microbiol. 2006, 4, 176–182. [Google Scholar] [CrossRef]
- Pal, C.; Asiani, K.; Arya, S.; Rensing, C.; Stekel, D.J.; Larsson, D.G.J.; Hobman, J.L. Metal resistance and its association with antibiotic resistance. Adv. Microb. Physiol. 2017, 70, 261–313. [Google Scholar]
- Stepanauskas, R.; Glenn, T.C.; Jagoe, C.H.; Tuckfield, R.C.; Lindell, A.H.; King, C.J.; McArthur, J.V. Coselection for microbial resistance to metals and antibiotics in freshwater microcosms. Environ. Microbiol. 2006, 8, 1510–1514. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 417. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Kimura, M.; Agusa, T.; Rahman, H.M. Vanadium accelerates horizontal transfer of tet(M) gene from marine Photobacterium to Escherichia coli. FEMS Microbiol. Lett. 2012, 336, 52–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.; Shin, C.; Shin, Y.; Kim, P.H.; il Park, J.; Kim, M.; Park, B.; So, J.-S. Heavy metal and antibiotic co-resistance in Vibrio parahaemolyticus isolated from shellfish. Mar. Pollut. Bull. 2020, 156, 111246. [Google Scholar] [CrossRef] [PubMed]
- Ruelas-Inzunza, J.; Soto, L.A.; Páez-Osuna, F. Heavy-metal accumulation in the hydrothermal vent clam Vesicomya gigas from Guaymas basin, Gulf of California. Deep Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 757–761. [Google Scholar] [CrossRef]
- Chen, X.-G.; Lyu, S.-S.; Garbe-Schönberg, D.; Lebrato, M.; Li, X.; Zhang, H.-Y.; Zhang, P.-P.; Chen, C.-T.A.; Ye, Y. Heavy metals from Kueishantao shallow-sea hydrothermal vents, offshore northeast Taiwan. J. Mar. Syst. 2018, 180, 211–219. [Google Scholar] [CrossRef]
- Farias, P.; Espírito Santo, C.; Branco, R.; Francisco, R.; Santos, S.; Hansen, L.; Sorensen, S.; Morais, P.V. Natural hot spots for gain of multiple resistances: Arsenic and antibiotic resistances in heterotrophic, aerobic bacteria from marine hydrothermal vent fields. Appl. Environ. Microbiol. 2015, 81, 2534–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andaloro, F.; Romeo, T.; Renzi, M.; Guerranti, C.; Perra, G.; Consoli, P.; Perzia, P.; Focardi, S.E. Alteration of potential harmful elements levels in sediments and biota from the central Mediterranean Sea (Aeolian Archipelago) following an episode of intense volcanic activity. Environ. Monit. Assess. 2012, 184, 4035–4047. [Google Scholar] [CrossRef]
- Tarasov, V.G.; Gebruk, A.V.; Mironov, A.N.; Moskalev, L.I. Deep-sea and shallow-water hydrothermal vent communities: Two different phenomena? Chem. Geol. 2005, 224, 5–39. [Google Scholar] [CrossRef]
- Rastelli, E.; Corinaldesi, C.; Dell’Anno, A.; Tangherlini, M.; Martorelli, E.; Ingrassia, M.; Chiocci, F.L.; Lo Martire, M.; Danovaro, R. High potential for temperate viruses to drive carbon cycling in chemoautotrophy-dominated shallow-water hydrothermal vents. Environ. Microbiol. 2017, 19, 4432–4446. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, W.; Bao, W.; Chen, C.-T.A.; Liu, S.; Cai, Y. Recent Developments of Exploration and Detection of Shallow-Water Hydrothermal Systems. Sustainability 2020, 12, 9109. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Italiano, F.; Maugeri, T.L. The submarine hydrothermal system of Panarea (Southern Italy): Biogeochemical processes at the thermal fluids-sea bottom interface. Ann. Geophys. 2006, 49, 783–792. [Google Scholar] [CrossRef]
- Chiodini, G.; Caliro, S.; Caramanna, G.; Granieri, D.; Minopoli, C.; Moretti, R.; Perotta, L.; Ventura, G. Geochemistry of the submarine gaseous emissions of Panarea (Aeolian Islands, Southern Italy): Magmatic vs. hydrothermal origin and implications for volcanic surveillance. Pure Appl. Geophys. 2006, 163, 759–780. [Google Scholar] [CrossRef]
- Caracausi, A.; Ditta, M.; Italiano, F.; Longo, M.; Nuccio, P.M.; Paonita, A. Massive submarine gas output during the volcanic unrest off Panarea Island (Aeolian arc, Italy): Inferences for explosive conditions. Geochem. J. 2005, 39, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Esposito, A.; Giordano, G.; Anzidei, M. The 2002–2003 submarine gas eruption at Panarea volcano (Aeolian Islands, Italy): Volcanology of the seafloor and implications for the hazard scenario. Mar. Geol. 2006, 227, 119–134. [Google Scholar] [CrossRef]
- Aliani, S.; Bortoluzzi, G.; Caramanna, G.; Raffa, F. Seawater dynamics and environmental settings after November 2002 gas eruption off Bottaro (Panarea, Aeolian Islands, Mediterranean Sea). Cont. Shelf Res. 2010, 30, 1338–1348. [Google Scholar] [CrossRef]
- Caramanna, G.; Voltattorni, N.; Maroto-Valer, M.M. Is Panarea Island (Italy) a valid and cost-effective natural laboratory for the development of detection and monitoring techniques for submarine CO2 seepage? Greenh. Gases Sci. Technol. 2011, 1, 200–210. [Google Scholar] [CrossRef]
- Italiano, F. Hydrothermal fluids vented at shallow depths at the Aeolian islands: Relationships with volcanic and geothermal systems. Res. Shallow Mar. Fresh Water Syst. 2009, 22, 55–60. [Google Scholar]
- Rogelja, M.; Cibic, T.; Pennesi, C.; De Vittor, C. Microphytobenthic community composition and primary production at gas and thermal vents in the Aeolian Islands (Tyrrhenian Sea, Italy). Mar. Environ. Res. 2016, 118, 31–44. [Google Scholar] [CrossRef]
- Gaglioti, M.; Gambi, M.C. The natural laboratory of the CO2 vents off Panarea (Aeolian Islands, Italy): A special ecological setting and a further stepping stone for some alien macrophytes. Not. SIBM 2018, 74, 111–117. [Google Scholar]
- Sieland, R.; Steinbrückner, D.; Hamel, M.; Merkel, B.; Schipek, M. Geochemical Investigations and Gas Quantification of Submarine Fluid Discharges in the Hydrothermal System of Panarea (Aeolian Islands, Italy). In Proceedings of the 1st International Workshop Research in Shallow Marine and Fresh Water Systems, Freiberg, Germany, 14–16 May 2009; p. 87. [Google Scholar]
- Mishra, A.K.; Santos, R.; Hall-Spencer, J.M. Elevated trace elements in sediments and seagrasses at CO2 seeps. Mar. Environ. Res. 2020, 153, 104810. [Google Scholar] [CrossRef]
- Ingrosso, G.; Giani, M.; Comici, C.; Kralj, M.; Piacentino, S.; De Vittor, C.; Del Negro, P. Drivers of the carbonate system seasonal variations in a Mediterranean gulf. Estuar. Coast. Shelf Sci. 2016, 168, 58–70. [Google Scholar] [CrossRef]
- Pearson, T.H.; Stanley, S.O. Comparative measurement of the redox potential of marine sediments as a rapid means of assessing the effect of organic pollution. Mar. Biol. 1979, 53, 371–379. [Google Scholar] [CrossRef]
- Buck, J.D. Nonstaining (KOH) method for determination of gram reactions of marine bacteria. Appl. Environ. Microbiol. 1982, 44, 992–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- NCCLS. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-Sixth Edition; Approved document M7-A6; NCCLS: Wayne, PA, USA, 2003. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. Nucleic Acid Tech. Bact. Syst. 1991, 6, 115–175. [Google Scholar]
- Olsen, C.; Qaadri, K.; Moir, R.; Kearse, M.; Buxton, S.; Cheung, M. Geneious R7: A Bioinformatics Platform for Biologists. In Proceedings of the International Plant and Animal Genome Conference XXIII, San Diego, CA, USA, 10–14 January 2014. [Google Scholar]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Spagnoli, F.; Andaloro, F.; Canese, S.; Capaccioni, B.; Iassunto, R.A. New recent insights into the hydrothermal system of the Panarea Island (Aeolian Archipelago, South Tyrrhenian Sea). Mem. Descr. Carta Geol. Ital. 2020, 105, 85–90. [Google Scholar]
- Koonin, E.V. Horizontal gene transfer: Essentiality and evolvability in prokaryotes, and roles in evolutionary transitions. F1000Research 2016, 5, 1805. [Google Scholar] [CrossRef] [Green Version]
- Mata, M.T.; Baquero, F.; Perez-Diaz, J.C. A multidrug efflux transporter in Listeria monocytogenes. FEMS Microbiol. Lett. 2000, 187, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Aendekerk, S.; Ghysels, B.; Cornelis, P.; Baysse, C. Characterization of a new efflux pump, MexGHI-OpmD, from Pseudomonas aeruginosa that confers resistance to vanadium. Microbiology 2002, 148, 2371–2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shallcross, L.J.; Davies, D.S.C. Antibiotic overuse: A key driver of antimicrobial resistance. Br. J. Gen. Pract. 2014, 604–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinegani, A.A.S.; Younessi, N. Antibiotic resistance of bacteria isolated from heavy metal-polluted soils with different land uses. J. Glob. Antimicrob. Resist. 2017, 10, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Timková, I.; Lachká, M.; Kisková, J.; Maliničová, L.; Nosáľová, L.; Pristaš, P.; Sedláková-Kaduková, J. High frequency of antibiotic tolerance in deep subsurface heterotrophic cultivable bacteria from the Rozália Gold Mine, Slovakia. Environ. Sci. Pollut. Res. 2020, 27, 44036–44044. [Google Scholar] [CrossRef]
- Esposito, V.; Andaloro, F.; Canese, S.; Bortoluzzi, G.; Bo, M.; Di Bella, M.; Italiano, F.; Sabatino, G.; Battaglia, P.; Consoli, P. Exceptional discovery of a shallow-water hydrothermal site in the SW area of Basiluzzo islet (Aeolian archipelago, South Tyrrhenian Sea): An environment to preserve. PLoS ONE 2018, 13, e0190710. [Google Scholar] [CrossRef]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef]
- Dickinson, A.W.; Power, A.; Hansen, M.G.; Brandt, K.K.; Piliposian, G.; Appleby, P.; O’Neill, P.A.; Jones, R.T.; Sierocinski, P.; Koskella, B. Heavy metal pollution and co-selection for antibiotic resistance: A microbial palaeontology approach. Environ. Int. 2019, 132, 105117. [Google Scholar] [CrossRef]
- Beaber, J.W.; Hochhut, B.; Waldor, M.K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 2004, 427, 72–74. [Google Scholar] [CrossRef]
- Zhai, Y.; He, Z.; Kang, Y.; Yu, H.; Wang, J.; Du, P.; Zhang, Z.; Hu, S.; Gao, Z. Complete nucleotide sequence of pH11, an IncHI2 plasmid conferring multi-antibiotic resistance and multi-heavy metal resistance genes in a clinical Klebsiella pneumoniae isolate. Plasmid 2016, 86, 26–31. [Google Scholar] [CrossRef]
- Peltier, E.; Vincent, J.; Finn, C.; Graham, D.W. Zinc-induced antibiotic resistance in activated sludge bioreactors. Water Res. 2010, 44, 3829–3836. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Chen, L.; Ye, C.; Yu, X. Co-selection of antibiotic resistance via copper shock loading on bacteria from a drinking water bio-filter. Environ. Pollut. 2018, 233, 132–141. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, J.; Acinas, S.G.; Massana, R.; Rodríguez-Valera, F. Prevalence and microdiversity of Alteromonas macleodii-like microorganisms in different oceanic regions. Environ. Microbiol. 2002, 4, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas. Mar. Drugs 2007, 5, 220–241. [Google Scholar] [CrossRef]
- Lian, F.-B.; Li, Y.-Q.; Zhang, J.; Jiang, S.; Du, Z.-J. Sulfitobacter maritimus sp. nov., isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 2021, 71, 4659. [Google Scholar] [CrossRef]
- Zhu, S.; Xue, Z.; Huang, Y.; Chen, X.; Ren, N.; Chen, T.; Chen, Y.; Yang, J.; Chen, J. Muricauda sediminis sp. nov., isolated from western Pacific Ocean sediment. Int. J. Syst. Evol. Microbiol. 2021, 71, 4757. [Google Scholar] [CrossRef]
- Xu, M.; Xin, Y.; Yu, Y.; Zhang, J.; Zhou, Y.; Liu, H.; Tian, J.; Li, Y. Erythrobacter nanhaisediminis sp. nov., isolated from marine sediment of the South China Sea. Int. J. Syst. Evol. Microbiol. 2010, 60, 2215–2220. [Google Scholar] [CrossRef] [Green Version]
- Tonon, L.A.C.; Moreira, A.P.B.; Thompson, F. The family erythrobacteraceae. Prokaryotes 2014, 213–235. [Google Scholar]
- Mawlankar, R.R.; Mual, P.; Sonalkar, V.V.; Thorat, M.N.; Verma, A.; Srinivasan, K.; Dastager, S.G. Microbacterium enclense sp. nov., isolated from sediment sample. Int. J. Syst. Evol. Microbiol. 2015, 65, 2064–2070. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Y.; Zhang, G. Nocardioides flavus sp. nov., isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 2016, 66, 5275–5280. [Google Scholar] [CrossRef]
- López-Pérez, M.; Rodriguez-Valera, F. Pangenome evolution in the marine bacterium Alteromonas. Genome Biol. Evol. 2016, 8, 1556–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.W.; Lee, H.; Kwon, B.-O.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kim, J.-J. Blastococcus litoris sp. nov., isolated from sea-tidal flat sediment. Int. J. Syst. Evol. Microbiol. 2018, 68, 3435–3440. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Zhu, S.; Chen, T.; Ren, N.; Chen, X.; Chen, Y.; Xue, Z.; Shen, X.; Huang, Y.; Yang, J. Muricauda oceani sp. nov., isolated from the East Pacific Ocean. Int. J. Syst. Evol. Microbiol. 2020, 70, 3839–3844. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, S. Kocuria coralli sp. nov., a novel actinobacterium isolated from coral reef seawater. Int. J. Syst. Evol. Microbiol. 2020, 70, 785–789. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Lentini, V.; Spanò, A.; Maugeri, T.L. New bacilli from shallow hydrothermal vents of Panarea Island (Italy) and their biotechnological potential. J. Appl. Microbiol. 2012, 112, 1102–1112. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M. The PATRIC Bioinformatics Resource Center: Expanding data and analysis capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-H.; Oh, T.-K.; Park, Y.-H. Erythrobacter seohaensis sp. nov. and Erythrobacter gaetbuli sp. nov., isolated from a tidal flat of the Yellow Sea in Korea. Int. J. Syst. Evol. Microbiol. 2005, 55, 71–75. [Google Scholar] [CrossRef]
- Ahn, I.-Y.; Kang, Y.-C.; Choi, J.-W. The influence of industrial effluents on intertidal benthic communities in Panweol, Kyeonggi Bay (Yellow Sea) on the west coast of Korea. Mar. Pollut. Bull. 1995, 30, 200–206. [Google Scholar] [CrossRef]
- Kyung Tae, K.I.M.; Eun Soo, K.I.M.; Sung Rok, C.H.O.; Kong Tae, R.A.; Lee, J.M. Distribution of heavy metals in the environmental samples of the Saemangeum coastal area. Coast. Environ. Ecosyst. Korea 2010, 71–90. [Google Scholar]
- Yuan, H.; Song, J.; Li, X.; Li, N.; Duan, L. Distribution and contamination of heavy metals in surface sediments of the South Yellow Sea. Mar. Pollut. Bull. 2012, 64, 2151–2159. [Google Scholar] [CrossRef]
- Murray, N.J.; Ma, Z.; Fuller, R.A. Tidal flats of the Yellow Sea: A review of ecosystem status and anthropogenic threats. Austral. Ecol. 2015, 40, 472–481. [Google Scholar] [CrossRef]
- Chung, C.-S.; Song, K.-H.; Choi, K.-Y.; Kim, Y.-I.; Kim, H.-E.; Jung, J.-M.; Kim, C.-J. Variations in the concentrations of heavy metals through enforcement of a rest-year system and dredged sediment capping at the Yellow Sea-Byung dumping site, Korea. Mar. Pollut. Bull. 2017, 124, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-M.; Giovannoni, S.J.; Ferriera, S.; Johnson, J.; Cho, J.-C. Complete genome sequence of Erythrobacter litoralis HTCC2594. J. Bacteriol. 2009, 191, 2419. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.P.; Gorshkova, N.M.; Sawabe, T.; Zhukova, N.V.; Hayashi, K.; Kurilenko, V.V.; Alexeeva, Y.; Buljan, V.; Nicolau, D.V.; Mikhailov, V.V. Sulfitobacter delicatus sp. nov. and Sulfitobacter dubius sp. nov., respectively from a starfish (Stellaster equestris) and sea grass (Zostera marina). Int. J. Syst. Evol. Microbiol. 2004, 54, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, C.; Farias, P.; Branco, R.; Morais, P.V. Tungsten accumulation by highly tolerant marine hydrothermal Sulfitobacter dubius strains carrying a tupBCA cluster. Syst. Appl. Microbiol. 2017, 40, 388–395. [Google Scholar] [CrossRef]
- Buchanan, J.A.; Scherer, S.W. Contemplating effects of genomic structural variation. Genet. Med. 2008, 10, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Wan, K.; Zeng, J.; Lin, W.; Ye, C.; Yu, X. Co-selection and stability of bacterial antibiotic resistance by arsenic pollution accidents in source water. Environ. Int. 2020, 135, 105351. [Google Scholar] [CrossRef]
- Zhao, X.; Shen, J.-P.; Zhang, L.-M.; Du, S.; Hu, H.-W.; He, J.-Z. Arsenic and cadmium as predominant factors shaping the distribution patterns of antibiotic resistance genes in polluted paddy soils. J. Hazard. Mater. 2020, 389, 121838. [Google Scholar] [CrossRef]
- Fu, J.; Zhong, C.; Zhang, P.; Gao, Q.; Zong, G.; Zhou, Y.; Cao, G. A Novel Mobile Element ICERspD18B in Rheinheimera sp. D18 Contributes to Antibiotic and Arsenic Resistance. Front. Microbiol. 2020, 11, 3275. [Google Scholar] [CrossRef]
- Morais, S.; Costa, F.G.; Pereira, M.D.L. Heavy metals and human health. Env. Health Emerg. Issues Pract. 2012, 10, 227–245. [Google Scholar]
- Stankovic, S.; Jovic, M.; Stankovic, A.R.; Katsikas, L. Heavy metals in seafood mussels. Risks for human health. In Environmental Chemistry for a Sustainable World; Springer: Dordrecht, The Netherlands, 2012; pp. 311–373. [Google Scholar]
- Bosch, A.C.; O’Neill, B.; Sigge, G.O.; Kerwath, S.E.; Hoffman, L.C. Heavy metals in marine fish meat and consumer health: A review. J. Sci. Food Agric. 2016, 96, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, D.; Słowik, J.; Chilicka, K. Heavy metals and human health: Possible exposure pathways and the competition for protein binding sites. Molecules 2021, 26, 6060. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Source | Location | Depth (M) | T (°C) | C (ms cm−1) | ORP (mV) | O2 (mL L−1) | pH | S |
|---|---|---|---|---|---|---|---|---|
| Bottom water | Bottaro Crater | 7.5 | 19.26 | 50.46 | - | 5.48 | 8.17 | 37.75 |
| “Hot” vent | 11.41 | 25.39 | 57.75 | - | 4.75 | 8.07 | 38.17 | |
| “Cold” vent | 11 | 26.2 | 58.47 | - | 4.71 | 7.89 | 38.02 | |
| Sediment | Bottaro Crater | 7.5 | 28 | - | 106.27 | - | 6.78 | - |
| “Hot” vent | 11.7 | 48 | - | 8.07 | - | 5.54 | - | |
| “Cold” vent | 11 | 26 | - | 7.87 | - | 5.91 | - |
| Strain | Origin | Cell Shape | Gram Staining (+/−) | Colony Phenotype (Colour/Aspect) |
|---|---|---|---|---|
| BC1 | Bottaro Crater | cocci | - | white/creamy |
| BC2 | Bottaro Crater | rods | - | yellow/opalescent |
| BC3 | Bottaro Crater | cocci | + | white/pearl |
| BC4 | Bottaro Crater | cocci | + | orange-pink/brilliant |
| BC5 | Bottaro Crater | rods | - | yellow/brilliant |
| CV1 | “Cold” vent | cocci | + | yellow/brilliant |
| CV2 | “Cold” vent | rods | + | yellow-white/creamy |
| CV3 | “Cold” vent | cocci | - | orange/brilliant |
| CV4 | “Cold” vent | cocci | - | Brown/liquid |
| CV5 | “Cold” vent | rods | - | orange/creamy |
| HV1 | “Hot” vent | rods | - | white/creamy |
| HV2 | “Hot” vent | cocci | - | yellow/creamy |
| HV3 | “Hot” vent | rods | - | white/creamy |
| HV4 | “Hot” vent | rods | - | yellow-white/creamy |
| HV5 | “Hot” vent | rods | - | yellow/brilliant |
| Strain | Class | Family | Ref Strain | Identity | S Range | T Range |
|---|---|---|---|---|---|---|
| BC1 | Gammaproteobacteria | Aeromonadaceae | Alteromonas marina strain ROA053 | 100% | 30–150 | 15–40 °C |
| BC2 | Alphaproteobacteria | Sphingomonadaceae | Erythrobacter pelagi strain UST081027-248 | 99.8% | 30–150 | 4–40 °C |
| BC3 | Actinobacteria | Microbacteriaceae | Microbacterium invictum strain DC-200 | 99.7% | 30–150 | 15–40 °C |
| BC4 | Actinobacteria | Geodermathophiliaceae | Blastococcus deserti strain SYSU D8006 | 99.3% | 30–100 | 15–50 °C |
| CV1 | Actinobacteria | Micrococcaceae | Kocuria sp. strain HY2 | 100% | 30–100 | 4–40 °C |
| CV2 | Actinobacteria | Nocardioidaceae | Nocardioides marinus strain CL-DD14 | 99.5% | 30–150 | 15–40 °C |
| CV3 | Alphaproteobacteria | Sphingomonadaceae | Erythrobacter pelagi strain UST081027-248 | 99.0% | 30–50 | 4–40 °C |
| CV5 | Alphaproteobacteria | Sphingomonadaceae | Erythrobacter sp. strain S44 | 99.8% | 30–100 | 4–40 °C |
| HV1 | Gammaproteobacteria | Aeromonadaceae | Alteromonas macleodii strain NBRC 102226 | 99.7% | 30–150 | 15–40 °C |
| HV3 | Gammaproteobacteria | Aeromonadaceae | Pseudoalteromonas shioyasakiensis strain SE3 | 99.8% | 30–150 | 15–40 °C |
| HV4 | Alphaproteobacteria | Rhodobacteraceae | Sulfitobacter faviae strain S5-53 | 99.8% | 30–150 | 4–40 °C |
| HV5 | Firmicutes | Flavobacteriaceae | Muricauda taeanensis strain 105 | 99.8% | 30–100 | 15–50 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcadi, E.; Rastelli, E.; Tangherlini, M.; Rizzo, C.; Mancuso, M.; Sanfilippo, M.; Esposito, V.; Andaloro, F.; Romeo, T. Shallow-Water Hydrothermal Vents as Natural Accelerators of Bacterial Antibiotic Resistance in Marine Coastal Areas. Microorganisms 2022, 10, 479. https://doi.org/10.3390/microorganisms10020479
Arcadi E, Rastelli E, Tangherlini M, Rizzo C, Mancuso M, Sanfilippo M, Esposito V, Andaloro F, Romeo T. Shallow-Water Hydrothermal Vents as Natural Accelerators of Bacterial Antibiotic Resistance in Marine Coastal Areas. Microorganisms. 2022; 10(2):479. https://doi.org/10.3390/microorganisms10020479
Chicago/Turabian StyleArcadi, Erika, Eugenio Rastelli, Michael Tangherlini, Carmen Rizzo, Monique Mancuso, Marilena Sanfilippo, Valentina Esposito, Franco Andaloro, and Teresa Romeo. 2022. "Shallow-Water Hydrothermal Vents as Natural Accelerators of Bacterial Antibiotic Resistance in Marine Coastal Areas" Microorganisms 10, no. 2: 479. https://doi.org/10.3390/microorganisms10020479