
Community Assembly and Co-Occurrence Patterns of Microeukaryotes in Thermokarst Lakes of the Yellow River Source Area


Abstract
:1. Introduction
2. Methods
2.1. Study Area, Field Sampling, and Chemical Analyses
2.2. DNA Extraction, PCR, and Sequencing
2.3. Analyses
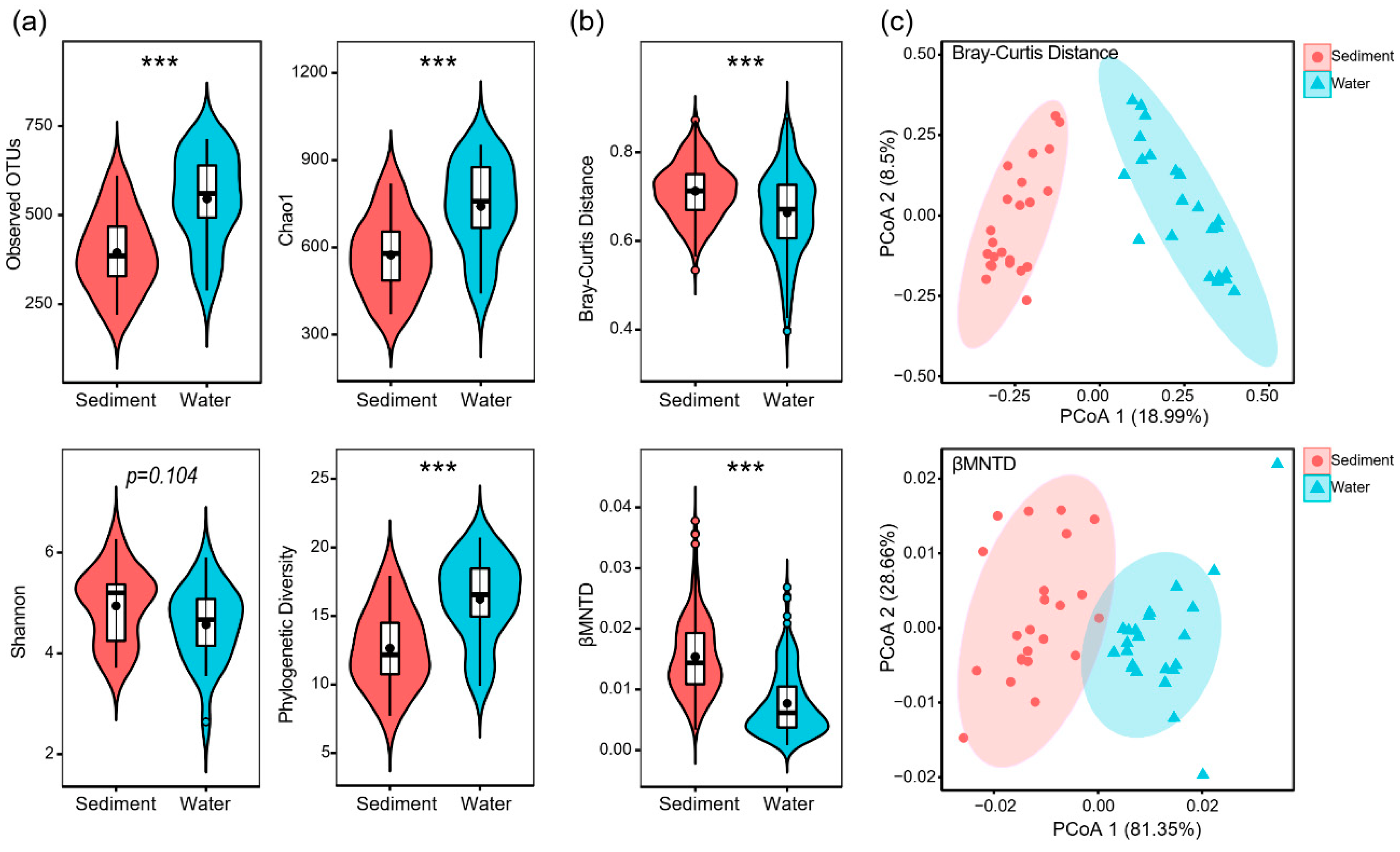
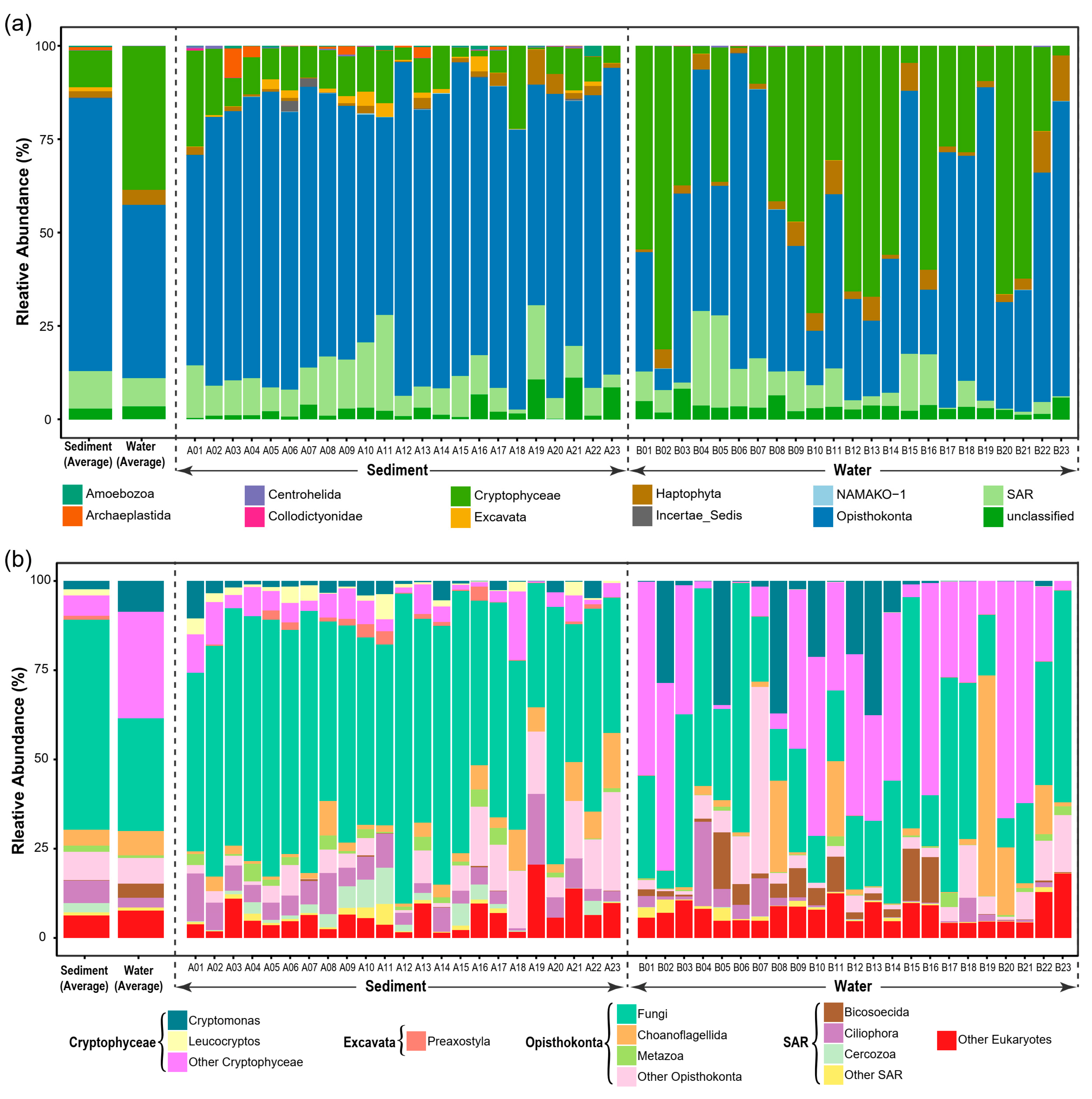
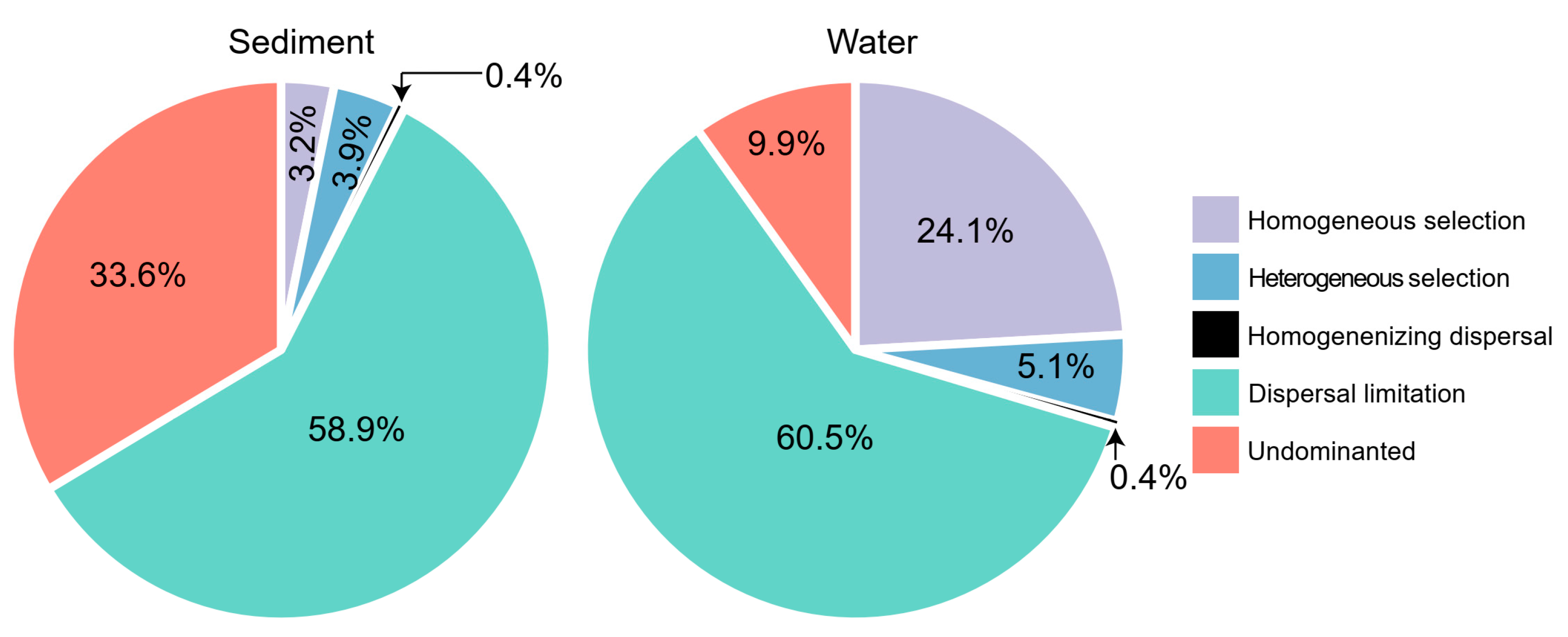
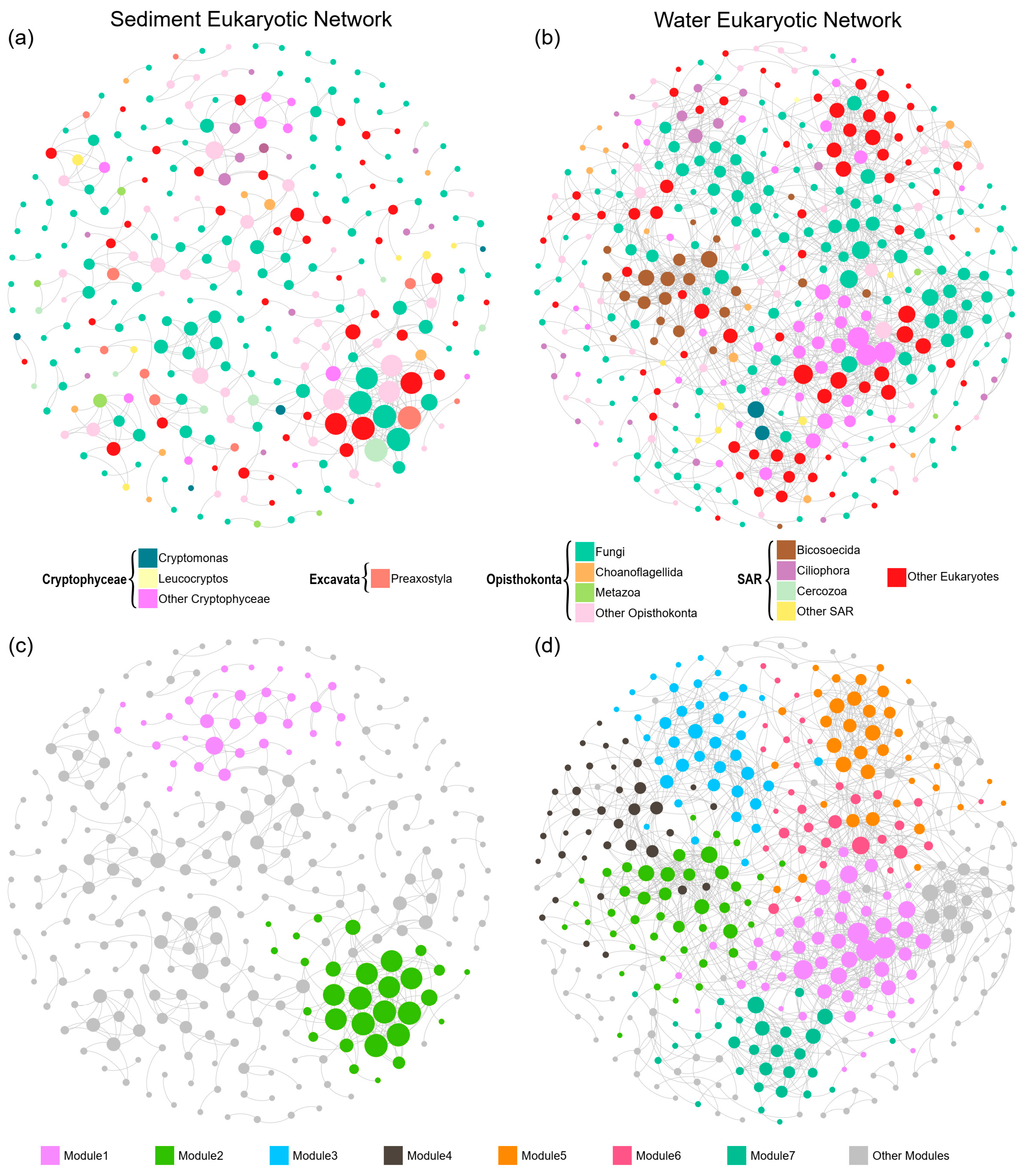
3. Results
3.1. General Patterns of Microeukaryotic Communities in Sediment and Water
3.2. Assembly Processes
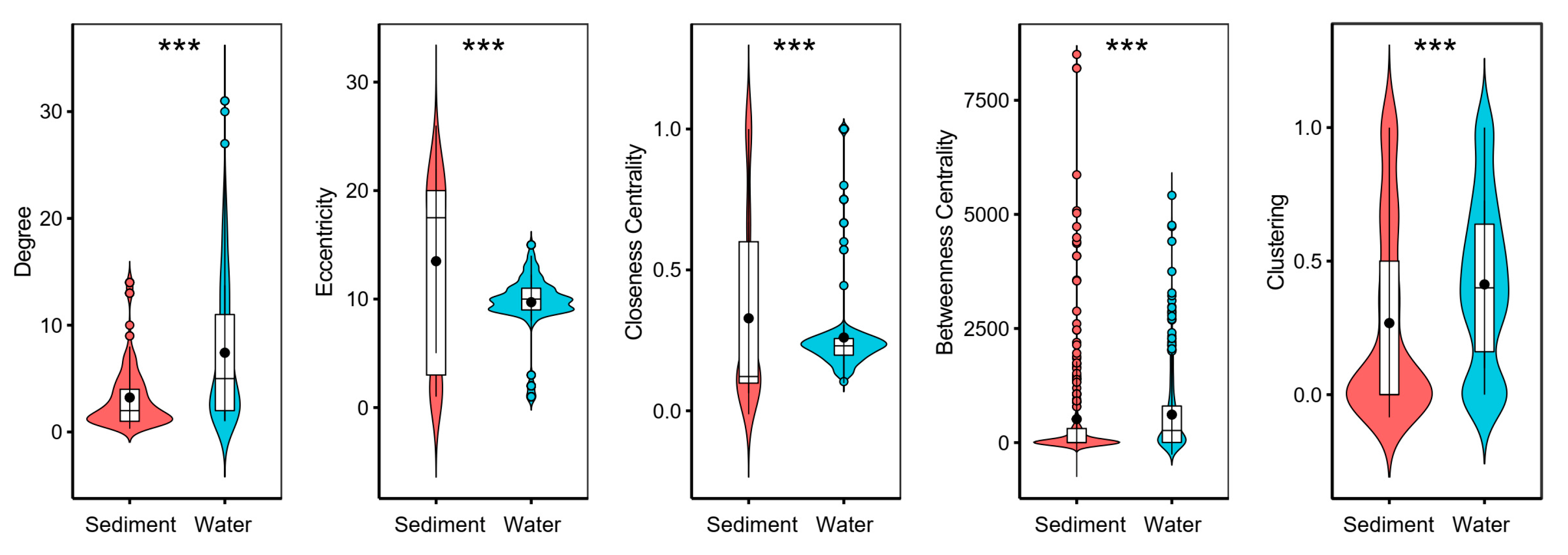
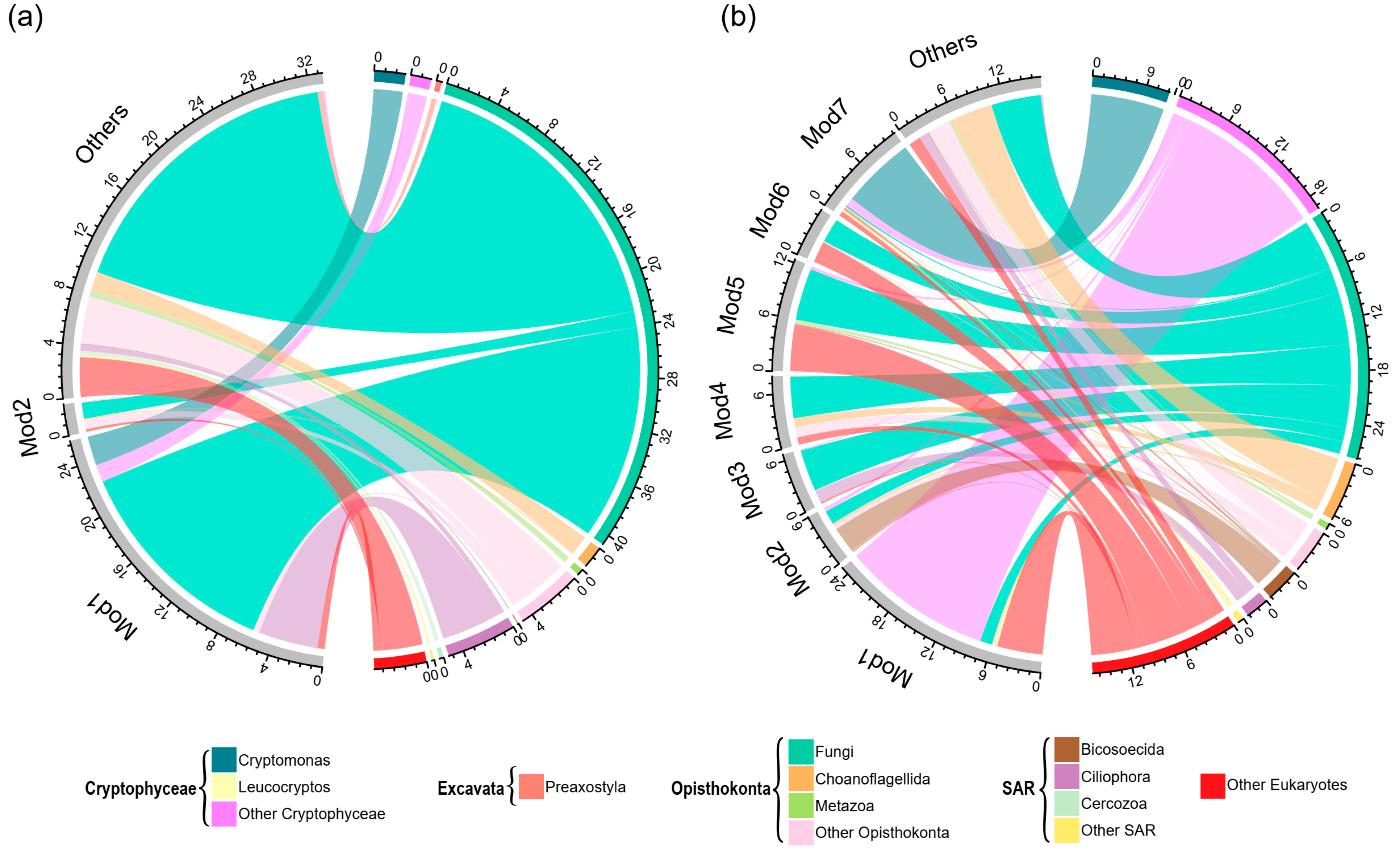
3.3. Co-Occurrence Networks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kokelj, S.V.; Jorgenson, M.T. Advances in Thermokarst Research. Permafr. Periglac. Process. 2013, 24, 108–119. [Google Scholar] [CrossRef]
- Farquharson, L.M.; Mann, D.H.; Grosse, G.; Jones, B.M.; Romanovsky, V.E. Spatial distribution of thermokarst terrain in Arctic Alaska. Geomorphology 2016, 273, 116–133. [Google Scholar] [CrossRef] [Green Version]
- Niu, F.; Lin, Z.; Liu, H.; Lu, J. Characteristics of thermokarst lakes and their influence on permafrost in Qinghai–Tibet Plateau. Geomorphology 2011, 132, 222–233. [Google Scholar] [CrossRef]
- Luo, D.-L.; Jin, H.-J.; Dong-Liang, L.; Ma, Q.; Duan, S.-Q.; Li, G.-S. Variation of alpine lakes from 1986 to 2019 in the Headwater Area of the Yellow River, Tibetan Plateau using Google Earth Engine. Adv. Clim. Chang. Res. 2020, 11, 11–21. [Google Scholar] [CrossRef]
- Veremeeva, A.; Nitze, I.; Günther, F.; Grosse, G.; Rivkina, E. Geomorphological and Climatic Drivers of Thermokarst Lake Area Increase Trend (1999–2018) in the Kolyma Lowland Yedoma Region, North-Eastern Siberia. Remote Sens. 2021, 13, 178. [Google Scholar] [CrossRef]
- Vincent, W.F.; Lemay, M.; Allard, M.; Wolfe, B.B. Adapting to Permafrost Change: A Science Framework. EOS 2013, 94, 373–375. [Google Scholar] [CrossRef]
- Biskaborn, B.K.; Smith, S.L.; Noetzli, J.; Matthes, H.; Vieira, G.; Streletskiy, D.A.; Schoeneich, P.; Romanovsky, V.E.; Lewkowicz, A.G.; Abramov, A.; et al. Permafrost is warming at a global scale. Nat. Commun. 2019, 10, 264. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Niu, F.; Lin, Z.; Liu, M.; Yin, G. Thermokarst lake changes between 1969 and 2010 in the Beilu River Basin, Qinghai–Tibet Plateau, China. Sci. Bull. 2015, 60, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Grosse, G.; Jones, B.M.; Arp, C.D. Thermokarst Lake, Drainage, and Drained Basins, in Treatise on Geomorphology; Shroder, J., Giardino, R., Harbor, J., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2013; pp. 1–29. [Google Scholar]
- Sepulveda-Jauregui, A.; Anthony, K.M.W.; Martinez-Cruz, K.; Greene, S.; Thalasso, F. Methane and carbon dioxide emissions from 40 lakes along a north–south latitudinal transect in Alaska. Biogeosciences 2015, 12, 3197–3223. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Zhang, C.; Li, X.; Ma, K.; Cui, B. Abundant and rare bacterial taxa structuring differently in sediment and water in thermokarst lakes in the Yellow River Source area, Qinghai-Tibet Plateau. bioRxiv 2021, 445882. [Google Scholar] [CrossRef]
- Toniolo, H.; Kodial, P.; Hinzman, L.; Yoshikawa, K. Spatio-temporal evolution of a thermokarst in Interior Alaska. Cold Reg. Sci. Technol. 2009, 56, 39–49. [Google Scholar] [CrossRef]
- Niu, F.; Lin, Z.; Lu, J.; Luo, J.; Wang, H. Assessment of terrain susceptibility to thermokarst lake development along the Qinghai–Tibet engineering corridor, China. Environ. Earth Sci. 2014, 73, 5631–5642. [Google Scholar] [CrossRef]
- Smith, L.C.; Sheng, Y.; Macdonald, G.M.; Hinzman, L.D. Disappearing Arctic Lakes. Science 2005, 308, 1429. [Google Scholar] [CrossRef] [Green Version]
- Schaechter, M. Eukaryotic Microbes; Academic Press: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Salmaso, N.; Boscaini, A.; Pindo, M. Unraveling the Diversity of Eukaryotic Microplankton in a Large and Deep Perialpine Lake Using a High Throughput Sequencing Approach. Front. Microbiol. 2020, 11, 789. [Google Scholar] [CrossRef]
- Pearman, J.K.; Biessy, L.; Thomson-Laing, G.; Waters, S.; Vandergoes, M.J.; Howarth, J.D.; Rees, A.; Moy, C.; Pochon, X.; Wood, S.A. Local factors drive bacterial and microeukaryotic community composition in lake surface sediment collected across an altitudinal gradient. FEMS Microbiol. Ecol. 2020, 96, fiaa070. [Google Scholar] [CrossRef]
- Zubkov, M.V.; Tarran, G.A. High bacterivory by the smallest phytoplankton in the North Atlantic Ocean. Nature 2008, 455, 224–226. [Google Scholar] [CrossRef]
- Sherr, B.; Sherr, E.; Caron, D.; Vaulot, D.; Worden, A. Oceanic Protists. Oceanography 2007, 20, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Genet. 2012, 10, 497–506. [Google Scholar] [CrossRef]
- Capo, E.; Ninnes, S.; Domaizon, I.; Bertilsson, S.; Bigler, C.; Wang, X.-R.; Bindler, R.; Rydberg, J. Landscape Setting Drives the Microbial Eukaryotic Community Structure in Four Swedish Mountain Lakes over the Holocene. Microorganisms 2021, 9, 355. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, H.; Yang, J.R.; Liu, M.; Huang, B.; Yang, J. Distinct patterns and processes of abundant and rare eukaryotic plankton communities following a reservoir cyanobacterial bloom. ISME J. 2018, 12, e2263. [Google Scholar] [CrossRef] [Green Version]
- Coesel, S.N.; Durham, B.P.; Groussman, R.D.; Hu, S.K.; Caron, D.A.; Morales, R.L.; Ribalet, F.; Armbrust, E.V. Diel transcriptional oscillations of light-sensitive regulatory elements in open-ocean eukaryotic plankton communities. Proc. Natl. Acad. Sci. USA 2021, 118, e2011038118. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; López-García, P.; Deschamps, P.; Moreira, D.; Restoux, G.; Bertolino, P.; Jardillier, L. Marked seasonality and high spatial variability of protist communities in shallow freshwater systems. ISME J. 2015, 9, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying communi-ty assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ning, D. Stochastic Community Assembly: Does It Matter in Microbial Ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002-17. [Google Scholar] [CrossRef] [Green Version]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and Processes of Microbial Community Assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Chase, J.M. Stochastic Community Assembly Causes Higher Biodiversity in More Productive Environments. Science 2010, 328, 1388–1391. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Deng, Y.; Zhang, P.; Xue, K.; Liang, Y.; Van Nostrand, J.; Yang, Y.; He, Z.; Wu, L.; Stahl, D.A.; et al. Stochasticity, succession, and environmental perturbations in a fluidic ecosystem. Proc. Natl. Acad. Sci. USA 2014, 111, E836–E845. [Google Scholar] [CrossRef] [Green Version]
- Chave, J. Neutral theory and community ecology. Ecol. Lett. 2004, 7, 241–253. [Google Scholar] [CrossRef]
- Aguilar, P.; Sommaruga, R. The balance between deterministic and stochastic processes in structuring lake bacterioplankton community over time. Mol. Ecol. 2020, 29, 3117–3130. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.; Lin, X.; Konopka, A.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef] [Green Version]
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Mccabe, D.J. Species co-occurrence: A meta-analysis of J. M. Diamond’s assembly rules model. Ecology 2002, 83, 2091–2096. [Google Scholar] [CrossRef]
- Banerjee, S.; Schlaeppi, K.; Van Der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling Interactions in the Microbiome: A Network Perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef]
- Carter, J.L.; Topping, B.R.; Kuwabara, J.S.; Balistrieri, L.S.; Woods, P.F.; Berelson, W.M.; Fend, S.V. Importance of Sediment? Water Interactions in Coeur Alene Lake, Idaho, USA: Management Implications. Environ. Manag. 2003, 32, 348–359. [Google Scholar] [CrossRef]
- Parker, S.R.; West, R.F.; Boyd, E.S.; Feyhl-Buska, J.; Gammons, C.H.; Johnston, T.B.; Williams, G.P.; Poulson, S.R. Biogeochemical and microbial seasonal dynamics between water column and sediment processes in a productive mountain lake: Georgetown Lake, MT, USA. J. Geophys. Res. Biogeosci. 2016, 121, 2064–2081. [Google Scholar] [CrossRef] [Green Version]
- Serikova, S.; Pokrovsky, O.S.; Laudon, H.; Krickov, I.V.; Lim, A.G.; Manasypov, R.M.; Karlsson, J. High carbon emissions from thermokarst lakes of Western Siberia. Nat. Commun. 2019, 10, 1552. [Google Scholar] [CrossRef] [Green Version]
- Manasypov, R.M.; Pokrovsky, O.S.; Shirokova, L.S.; Auda, Y.; Zinner, N.S.; Vorobyev, S.N.; Kirpotin, S.N. Biogeochemistry of macrophytes, sediments and porewaters in thermokarst lakes of permafrost peatlands, western Siberia. Sci. Total Environ. 2021, 763, 144201. [Google Scholar] [CrossRef]
- Kokelj, S.V.; Jenkins, R.E.; Milburn, D.; Burn, C.R.; Snow, N. The influence of thermokarst disturbance on the water quality of small upland lakes, Mackenzie Delta region, Northwest Territories, Canada. Permafr. Periglac. Process. 2005, 16, 343–353. [Google Scholar] [CrossRef]
- Ren, Z.; Zhang, C.; Li, X.; Ma, K.; Zhang, Z.; Feng, K.; Cui, B. Bacterial communities present distinct co-occurrence net-works in sediment and water of the thermokarst lakes in the Yellow River Source Area. Front. Microbiol. 2021, 12, e716732. [Google Scholar] [CrossRef]
- Qiu, J. China: The third pole. Nature 2008, 454, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Yao, T.; Thompson, L.G.; Mosbrugger, V.; Zhang, F.; Ma, Y.; Luo, T.; Xu, B.; Yang, X.; Joswiak, D.R.; Wang, W.; et al. Third Pole Environment (TPE). Environ. Dev. 2012, 3, 52–64. [Google Scholar] [CrossRef]
- Qin, Y.; Yang, D.; Gao, B.; Wang, T.; Chen, J.; Chen, Y.; Wang, Y.; Zheng, G. Impacts of climate warming on the frozen ground and eco-hydrology in the Yellow River source region, China. Sci. Total Environ. 2017, 605–606, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Yao, T.; Piao, S.; Bolch, T.; Xie, H.; Chen, D.; Gao, Y.; O’Reilly, C.M.; Shum, C.K.; Yang, K.; et al. Extensive and drastically different alpine lake changes on Asia’s high plateaus during the past four decades. Geophys. Res. Lett. 2017, 44, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Zhang, L.; Liu, C.; Shao, Q.; Fukushima, Y. Changes in stream flow regime in headwater catchments of the Yellow River basin since the 1950s. Hydrol. Process. 2007, 21, 886–893. [Google Scholar] [CrossRef]
- Hu, Y.; Maskey, S.; Uhlenbrook, S. Trends in temperature and rainfall extremes in the Yellow River source region, China. Clim. Chang. 2012, 110, 403–429. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Sheng, Y.; Wu, J.; Feng, Z.; Ning, Z.; Hu, X.; Zhang, X. Landform-related permafrost characteristics in the source area of the Yellow River, eastern Qinghai-Tibet Plateau. Geomorphology 2016, 269, 104–111. [Google Scholar] [CrossRef]
- Dancer, W.S.; Eliason, R.; Lekhakul, S. Microwave assisted soil and waste dissolution for estimation of total phosphorus. Commun. Soil Sci. Plant Anal. 1998, 29, 1997–2006. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Csardi, M.G. Package ‘Igraph’. 2013. Available online: https://cran.r-project.org/web/packages/igraph/index.html (accessed on 1 August 2021).
- Levins, R. Evolution in Changing Environments: Some Theoretical Explorations (No. 2); Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Zhang, J.L. Species Association Analysis, Version 0.2.2. 2016. Available online: https://CRAN.R-project.org/package=spaa (accessed on 1 August 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Briée, C.; Moreira, D.; Lopez-Garcia, P. Archaeal and bacterial community composition of sediment and plankton from a suboxic freshwater pond. Res. Microbiol. 2007, 158, 213–227. [Google Scholar] [CrossRef]
- Gough, H.L.; Stahl, D. Microbial community structures in anoxic freshwater lake sediment along a metal contamination gradient. ISME J. 2010, 5, 543–558. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Ma, L.; Jiang, H.; Wu, G.; Dong, H. Salinity shapes microbial diversity and community structure in surface sediments of the Qinghai-Tibetan Lakes. Sci. Rep. 2016, 6, e25078. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Wang, F.; Qu, X.; Elser, J.J.; Liu, Y.; Chu, L. Taxonomic and Functional Differences between Microbial Communities in Qinghai Lake and Its Input Streams. Front. Microbiol. 2017, 8, 2319. [Google Scholar] [CrossRef]
- Langenheder, S.; Wang, J.; Karjalainen, S.M.; Laamanen, T.M.; Tolonen, K.T.; Vilmi, A.; Heino, J. Bacterial metacommunity or-ganization in a highly connected aquatic system. FEMS Microbiol. Ecol. 2017, 93, fiw225. [Google Scholar]
- Ren, Z.; Qu, X.; Peng, W.; Yu, Y.; Zhang, M. Nutrients Drive the Structures of Bacterial Communities in Sediments and Surface Waters in the River-Lake System of Poyang Lake. Water 2019, 11, 930. [Google Scholar] [CrossRef] [Green Version]
- Cline, L.C.; Zak, D.R. Dispersal limitation structures fungal community assembly in a long-term glacial chronosequence. Environ. Microbiol. 2014, 16, 1538–1548. [Google Scholar] [CrossRef] [Green Version]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Konopka, A.E. Estimating and mapping ecological processes influencing microbial community assembly. Front. Microbiol. 2015, 6, 370. [Google Scholar] [CrossRef] [Green Version]
- Vigneron, A.; Lovejoy, C.; Cruaud, P.; Kalenitchenko, D.; Culley, A.; Vincent, W.F. Contrasting Winter Versus Summer Microbial Communities and Metabolic Functions in a Permafrost Thaw Lake. Front. Microbiol. 2019, 10, 1656. [Google Scholar] [CrossRef] [Green Version]
- Vargas Medrano, M. Recent Changes in the Zooplankton Communities of Arctic Tundra Ponds in Response to Warmer Temperatures and Nutrient Enrichment. Ph.D. Thesis, University of Texas at El Paso, El Paso, TX, USA, 2019. [Google Scholar]
- Telford, R.J.; Vandvik, V.; Birks, H.J.B. Dispersal Limitations Matter for Microbial Morphospecies. Science 2006, 312, 1015. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Jiang, H.; Dong, H.; Liu, Y. A comprehensive census of lake microbial diversity on a global scale. Sci. China Life Sci. 2019, 62, 1320–1331. [Google Scholar] [CrossRef]
- Liu, K.; Yao, T.; Pearce, D.A.; Jiao, N.; Zeng, Y.; Guo, B.; Liu, Y. Bacteria in the lakes of the Tibetan Plateau and polar regions. Sci. Total Environ. 2021, 754, 142248. [Google Scholar] [CrossRef]
- Lindström, E.S.; Langenheder, S. Local and regional factors influencing bacterial community assembly. Environ. Microbiol. Rep. 2011, 4, 1–9. [Google Scholar] [CrossRef]
- Bottos, E.M.; Kennedy, D.W.; Romero, E.B.; Fansler, S.J.; Brown, J.M.; Bramer, L.M.; Chu, R.K.; Tfaily, M.; Jansson, J.; Stegen, J.C. Dispersal limitation and thermodynamic constraints govern spatial structure of permafrost microbial communities. FEMS Microbiol. Ecol. 2018, 94, fiy110. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ren, Z.; Qu, X.; Zhang, M.; Yu, Y.; Peng, W. Seasonal Water Level Fluctuation and Concomitant Change of Nutrients Shift Microeukaryotic Communities in a Shallow Lake. Water 2020, 12, 2317. [Google Scholar] [CrossRef]
- Mikhailov, I.S.; Zakharova, Y.R.; Bukin, Y.S.; Galachyants, Y.P.; Petrova, D.; Sakirko, M.V.; Likhoshway, Y.V. Co-occurrence Networks Among Bacteria and Microbial Eukaryotes of Lake Baikal During a Spring Phytoplankton Bloom. Microb. Ecol. 2019, 77, 96–109. [Google Scholar] [CrossRef]
- Ren, Z.; Gao, H. Ecological networks reveal contrasting patterns of bacterial and fungal communities in glacier-fed streams in Central Asia. PeerJ 2019, 7, e7715. [Google Scholar] [CrossRef] [Green Version]
- Tu, Q.; Yan, Q.; Deng, Y.; Michaletz, S.T.; Buzzard, V.; Weiser, M.D.; Waide, R.; Ning, D.; Wu, L.; He, Z.; et al. Biogeographic patterns of microbial co-occurrence ecological networks in six American forests. Soil Biol. Biochem. 2020, 148, 107897. [Google Scholar] [CrossRef]
- Levy, R.; Borenstein, E. Metabolic modeling of species interaction in the human microbiome elucidates community-level assembly rules. Proc. Natl. Acad. Sci. USA 2013, 110, 12804–12809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Meng, Z.; Liu, X.; Zhang, X.H. Microbial assembly, interaction, functioning, activity and diversification: A review de-rived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef] [Green Version]
- Mccann, K.S. The diversity-stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Kara, E.L.; Hanson, P.C.; Hu, Y.H.; Winslow, L.; McMahon, K.D. A decade of seasonal dynamics and co-occurrences within freshwater bacterioplankton communities from eutrophic Lake Mendota, WI, USA. ISME J. 2012, 7, 680–684. [Google Scholar] [CrossRef] [Green Version]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, Z.B.; Zak, D.R. Atmospheric N deposition alters connectance, but not functional potential among saprotrophic bacterial communities. Mol. Ecol. 2015, 24, 3170–3180. [Google Scholar] [CrossRef] [Green Version]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.N. The Geographic Mosaic of Coevolution; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Newman, M.E.J. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil micro-bial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Menezes, A.B.; Prendergast-Miller, M.; Richardson, A.E.; Toscas, P.; Farrell, M.; Macdonald, L.; Baker, G.; Wark, T.; Thrall, P.H. Network analysis reveals that bacteria and fungi form modules that correlate independently with soil parameters. Environ. Microbiol. 2015, 17, 2677–2689. [Google Scholar] [CrossRef]
- Toju, H.; Kishida, O.; Katayama, N.; Takagi, K. Networks Depicting the Fine-Scale Co-Occurrences of Fungi in Soil Horizons. PLoS ONE 2016, 11, e0165987. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Conductivity | SOC | TN | TP | |
|---|---|---|---|---|---|
| βNTI | −0.041 | 0.075 | 0.115 | 0.175 ** | 0.137 * |
| βMNTD | 0.197 ** | 0.084 | 0.145 * | 0.116 | 0.146 * |
| Bray–Curtis Distance | 0.250 ** | −0.026 | 0.173 ** | 0.052 | 0.248 ** |
| Module1 | 0.130 * | 0.040 | 0.255 ** | 0.107 | 0.358 ** |
| Module2 | −0.016 | 0.067 | −0.079 | −0.035 | 0.071 |
| Cryptomonas sp. | −0.015 | 0.074 | 0.223 ** | 0.131 * | 0.236 ** |
| Leucocryptos sp. | 0.138 * | −0.112 | −0.051 | 0.006 | 0.068 |
| Preaxostyla | −0.069 | 0.208 ** | −0.078 | −0.028 | −0.088 |
| Fungi | 0.190 ** | −0.039 | 0.152 * | 0.058 | 0.233 ** |
| Choanoflagellida | 0.043 | 0.039 | −0.119 | −0.091 | 0.004 |
| Metazoa | 0.198 ** | 0.187 ** | 0.056 | 0.069 | 0.127 * |
| Bicosoecida | NA | NA | NA | NA | NA |
| Ciliophora | 0.122 | −0.03 | 0.188 ** | 0.029 | 0.228 ** |
| Cercozoa | 0.172 ** | 0.165 ** | 0.037 | 0.098 | 0.281 ** |
| pH | Conductivity | DOC | TN | TP | |
|---|---|---|---|---|---|
| βNTI | −0.023 | 0.011 | −0.116 | 0.023 | 0.023 |
| βMNTD | −0.041 | 0.134 * | 0.067 | −0.007 | −0.004 |
| Bray–Curtis Distance | 0.071 | 0.144 * | 0.183 ** | −0.031 | −0.034 |
| Module1 | 0.118 | 0.063 | 0.129 * | −0.104 | −0.091 |
| Module2 | 0.126 * | 0.480 ** | 0.342 ** | 0.155 * | 0.267 ** |
| Module3 | −0.153 * | −0.043 | 0.021 | −0.113 | 0.112 |
| Module4 | −0.001 | 0.321 ** | 0.105 | 0.065 | 0.208 ** |
| Module5 | 0.082 | 0.032 | −0.044 | 0.048 | −0.063 |
| Module6 | 0.192 ** | 0.011 | −0.015 | 0.122 | 0.142 * |
| Module7 | 0.052 | 0.178 ** | 0.142 * | −0.042 | −0.005 |
| Cryptomonas sp. | 0.008 | 0.156 * | 0.133 * | −0.034 | 0.004 |
| Leucocryptos sp. | −0.029 | 0.373 ** | 0.170 * | −0.057 | −0.021 |
| Preaxostyla | NA | NA | NA | NA | NA |
| Fungi | 0.089 | 0.085 | 0.079 | 0.019 | 0.125 * |
| Choanoflagellida | 0.112 | 0.097 | 0.101 | 0.406 ** | 0.203 ** |
| Metazoa | 0.032 | 0.115 | 0.038 | 0.006 | 0.023 |
| Bicosoecida | 0.092 | 0.458 ** | 0.347 ** | 0.040 | 0.332 ** |
| Ciliophora | −0.155 * | 0.045 | −0.011 | −0.144 * | 0.121 |
| Cercozoa | 0.004 | −0.030 | −0.032 | 0.071 | −0.088 |
| Topological Parameters | Sediment | Water | ||
|---|---|---|---|---|
| This Study | Random | This Study | Random | |
| Number of Nodes | 284 | 284 | 376 | 376 |
| Number of Edges | 458 | 458 | 1395 | 1395 |
| Negative Edges | 12 | 12 | 140 | 140 |
| Average Degree | 3.225 | 3.225 | 7.420 | 7.420 |
| Graph Density | 0.011 | 0.011 | 0.02 | 0.02 |
| Average Path Length | 9.627 a | 4.821 ± 0.078 | 4.730 b | 3.177 ± 0.007 |
| Diameter | 26 a | 10.7 ± 0.879 | 15 b | 5.8 ± 0.413 |
| Clustering Coefficient | 0.460 a | 0.012 ± 0.005 | 0.528 b | 0.020 ± 0.002 |
| Centralization Degree | 0.038 a | 0.021 ± 0.003 | 0.063 b | 0.024 ± 0.004 |
| Centralization Betweenness | 0.201 a | 0.063 ± 0.014 | 0.069 b | 0.020 ± 0.004 |
| Centralization Closeness | 0.004 a | 0.013 ± 0.004 | 0.008 b | 0.097 ± 0.020 |
| Modularity | 0.836 a | 0.573 ± 0.01 | 0.639 b | 0.334 ± 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Z.; Ma, K.; Jia, X.; Wang, Q.; Zhang, C.; Li, X. Community Assembly and Co-Occurrence Patterns of Microeukaryotes in Thermokarst Lakes of the Yellow River Source Area. Microorganisms 2022, 10, 481. https://doi.org/10.3390/microorganisms10020481
Ren Z, Ma K, Jia X, Wang Q, Zhang C, Li X. Community Assembly and Co-Occurrence Patterns of Microeukaryotes in Thermokarst Lakes of the Yellow River Source Area. Microorganisms. 2022; 10(2):481. https://doi.org/10.3390/microorganisms10020481
Chicago/Turabian StyleRen, Ze, Kang Ma, Xuan Jia, Qing Wang, Cheng Zhang, and Xia Li. 2022. "Community Assembly and Co-Occurrence Patterns of Microeukaryotes in Thermokarst Lakes of the Yellow River Source Area" Microorganisms 10, no. 2: 481. https://doi.org/10.3390/microorganisms10020481