
Combination of High-Pressure Treatment at 500 MPa and Biopreservation with a Lactococcus lactis Strain for Lowering the Bacterial Growth during Storage of Diced Cooked Ham with Reduced Nitrite Salt

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Ham Sampling and Microbiota Recovery
2.3. Challenge Tests
2.4. HP Treatments
2.5. DNA Preparation and Amplicon Sequencing
2.6. Operational Taxonomic Unit (OTU) Analysis and Accession Numbers
2.7. Beta-Diversity and Statistical Analysis
3. Results
3.1. Diced Cooked Ham Selection for Microbiota Recovery
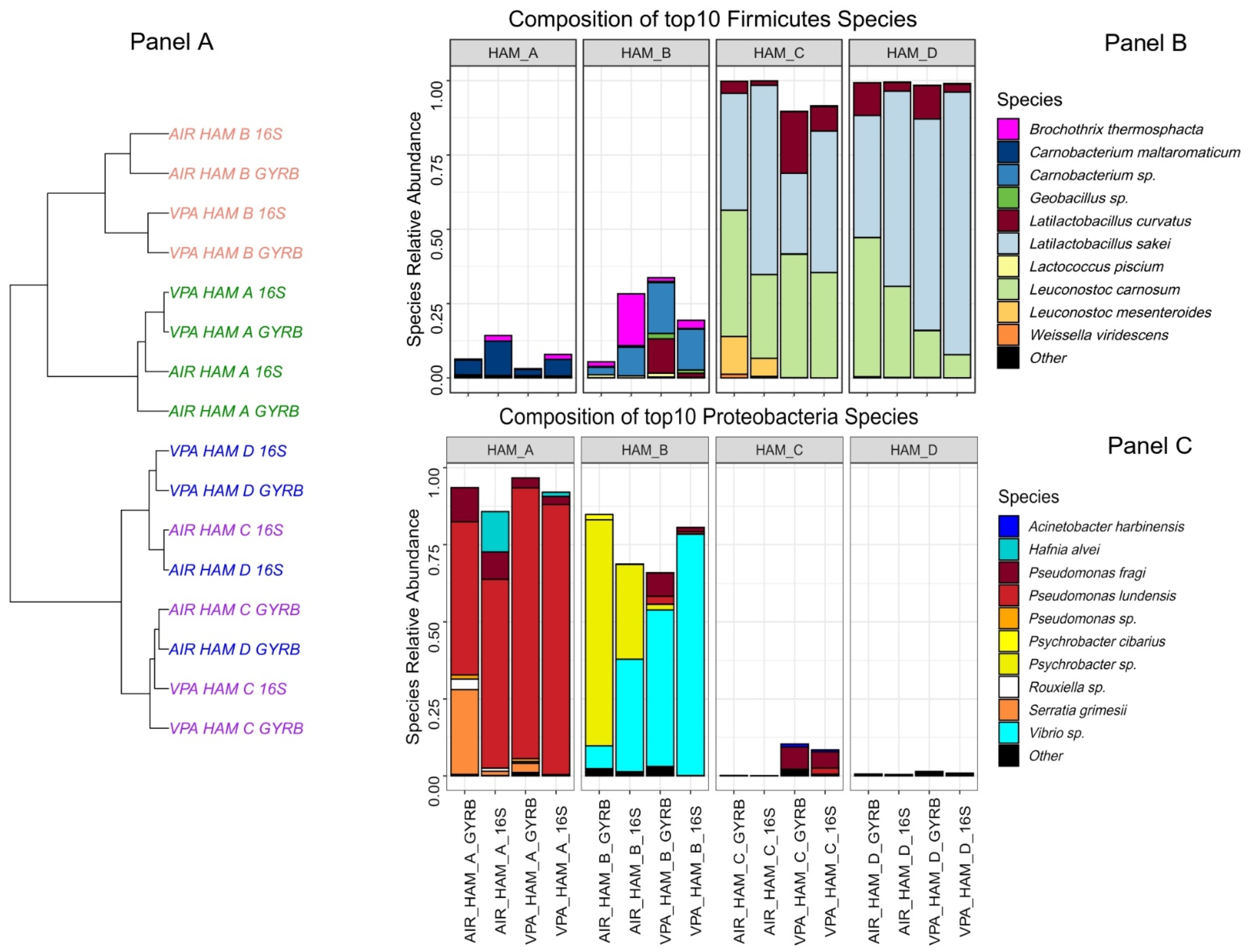
3.2. Four Different Diced Cooked Ham Samples Can Be Distinguished by the Diversity of Their Bacterial Communities
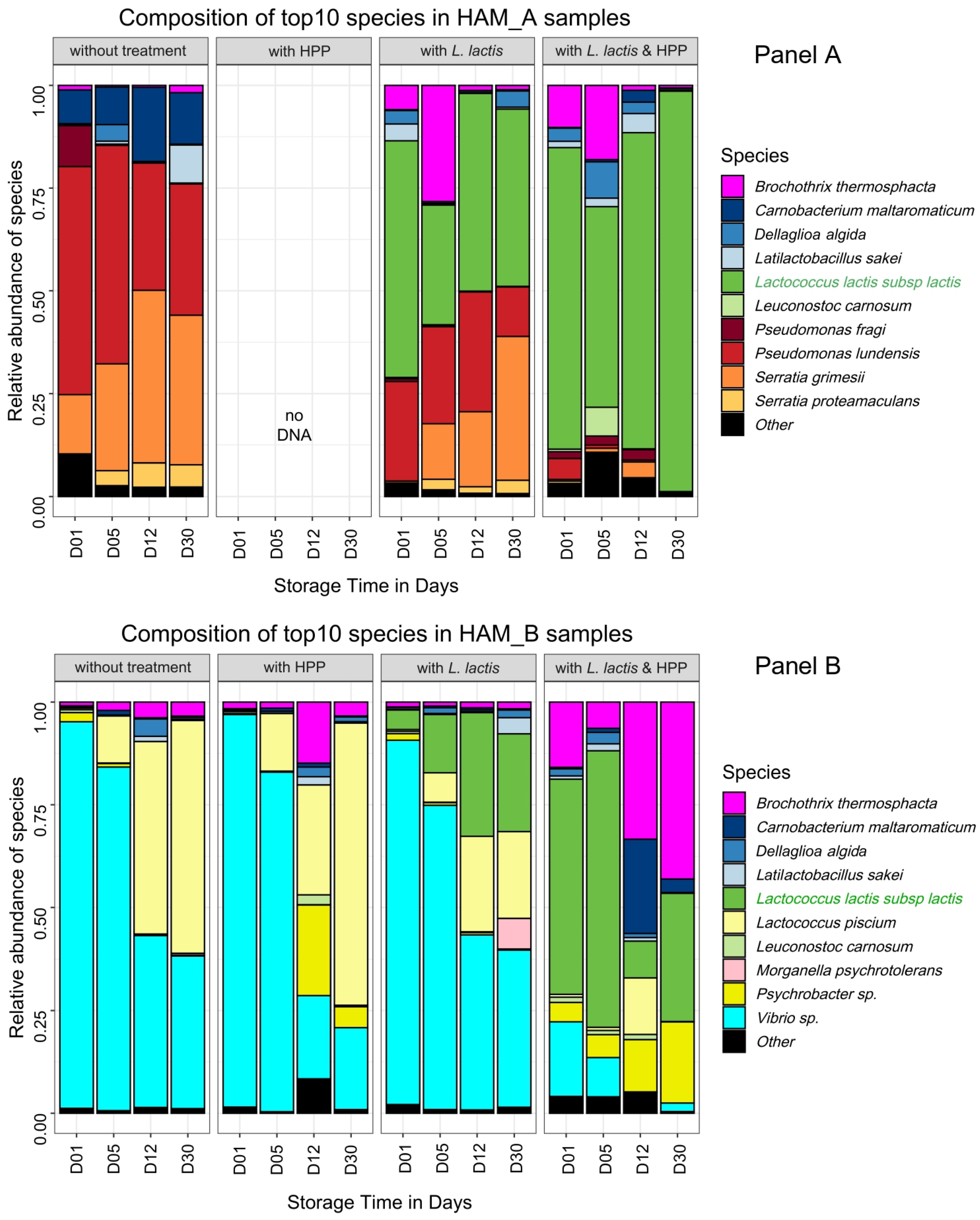
3.3. Combined Effect of HPP and Biopreservation on Bacterial Community Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alahakoon, A.U.; Jayasena, D.D.; Ramachandra, S.; Jo, C. Alternatives to nitrite in processed meat: Up to date. Trends Food Sci. Technol. 2015, 45, 37–49. [Google Scholar] [CrossRef]
- Majou, D.; Christieans, S. Mechanisms of the bactericidal effects of nitrate and nitrite in cured meats. Meat Sci. 2018, 145, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Sindelar, J.J.; Milkowski, A.L. Human safety controversies surrounding nitrate and nitrite in the diet. Nitric Oxide Biol. Chem. 2012, 26, 259–266. [Google Scholar] [CrossRef] [PubMed]
- EFSA. EFSA Explains Risk Assessment: Nitrites and Nitrates Added to Food. 2017. Available online: https://op.europa.eu/en/publication-detail/-/publication/20f25e61-5562-11e7-a5ca-01aa75ed71a1 (accessed on 1 December 2021).
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Domenico, A.D.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambré, C.; et al. Re-evaluation of potassium nitrite (E 249) and sodium nitrite (E 250) as food additives. EFSA J. 2017, 15, e04786. [Google Scholar] [CrossRef]
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Domenico, A.D.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambré, C.; et al. Re-evaluation of sodium nitrate (E 251) and potassium nitrate (E 252) as food additives. EFSA J. 2017, 15, e04787. [Google Scholar] [CrossRef]
- Roobab, U.; Khan, A.W.; Lorenzo, J.M.; Arshad, R.N.; Chen, B.-R.; Zeng, X.-A.; Bekhit, A.E.-D.; Suleman, R.; Aadil, R.M. A systematic review of clean-label alternatives to synthetic additives in raw and processed meat with a special emphasis on high-pressure processing (2018–2021). Food Res. Int. 2021, 150, 110792. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Serra-Castelló, C.; Dalgaard, P.; Garriga, M.; Jofré, A. New insights on Listeria monocytogenes growth in pressurised cooked ham: A piezo-stimulation effect enhanced by organic acids during storage. Int. J. Food Microbiol. 2019, 290, 150–158. [Google Scholar] [CrossRef]
- Han, Y.; Jiang, Y.; Xu, X.; Sun, X.; Xu, B.; Zhou, G. Effect of high pressure treatment on microbial populations of sliced vacuum-packed cooked ham. Meat Sci. 2011, 88, 682–688. [Google Scholar] [CrossRef]
- Hereu, A.; Dalgaard, P.; Garriga, M.; Aymerich, T.; Bover-Cid, S. Analysing and modelling the growth behaviour of Listeria monocytogenes on RTE cooked meat products after a high pressure treatment at 400 MPa. Int. J. Food Microbiol. 2014, 186, 84–94. [Google Scholar] [CrossRef]
- Jofré, A.; Garriga, M.; Aymerich, T. Inhibition of Listeria monocytogenes in cooked ham through active packaging with natural antimicrobials and high-pressure processing. J. Food Prot. 2007, 70, 2498–2502. [Google Scholar] [CrossRef]
- Jofré, A.; Aymerich, T.; Garriga, M. Assessment of the effectiveness of antimicrobial packaging combined with high pressure to control Salmonella sp. in cooked ham. Food Control 2008, 19, 634–638. [Google Scholar] [CrossRef]
- Jofré, A.; Garriga, M.; Aymerich, T. Inhibition of Salmonella sp. Listeria monocytogenes and Staphylococcus aureus in cooked ham by combining antimicrobials, high hydrostatic pressure and refrigeration. Meat Sci. 2008, 78, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Koseki, S.; Mizuno, Y.; Yamamoto, K. Predictive modelling of the recovery of Listeria monocytogenes on sliced cooked ham after high pressure processing. Int. J. Food Microbiol. 2007, 119, 300–307. [Google Scholar] [CrossRef] [PubMed]
- López-Caballero, M.E.; Carballo, J.; Jiménez-Colmenero, F. Microbiological changes in pressurized, prepackaged sliced cooked ham. J. Food Prot. 1999, 62, 1411–1415. [Google Scholar] [CrossRef] [Green Version]
- Marcos, B.; Jofré, A.; Aymerich, T.; Monfort, J.M.; Garriga, M. Combined effect of natural antimicrobials and high pressure processing to prevent Listeria monocytogenes growth after a cold chain break during storage of cooked ham. Food Control 2008, 19, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Pietrasik, Z.; Gaudette, N.J.; Johnston, S.P. The use of high pressure processing to enhance the quality and shelf life of reduced sodium naturally cured restructured cooked hams. Meat Sci. 2016, 116, 102–109. [Google Scholar] [CrossRef]
- Pingen, S.; Sudhaus, N.; Becker, A.; Krischek, C.; Klein, G. High pressure as an alternative processing step for ham production. Meat Sci. 2016, 118, 22–27. [Google Scholar] [CrossRef]
- Stiles, M.E. Biopreservation by lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 331–345. [Google Scholar] [CrossRef]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Co-culture experiments demonstrate the usefulness of Lactobacillus sakei 10A to prolong the shelf-life of a model cooked ham. Int. J. Food Microbiol. 2006, 108, 68–77. [Google Scholar] [CrossRef]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Evaluation of meat born lactic acid bacteria as protective cultures for the biopreservation of cooked meat products. Int. J. Food Microbiol. 2004, 96, 149–164. [Google Scholar] [CrossRef]
- de Oliveira, T.L.C.; Ramos, A.L.S.; Ramos, E.M.; Piccoli, R.H.; Cristianini, M. Natural antimicrobials as additional hurdles to preservation of foods by high pressure processing. Trends Food Sci. Technol. 2015, 45, 60–85. [Google Scholar] [CrossRef]
- Liu, G.; Wang, Y.; Gui, M.; Zheng, H.; Dai, R.; Li, P. Combined effect of high hydrostatic pressure and enterocin LM-2 on the refrigerated shelf life of ready-to-eat sliced vacuum-packed cooked ham. Food Control 2012, 24, 64–71. [Google Scholar] [CrossRef]
- Teixeira, J.S.; Repková, L.; Gänzle, M.G.; McMullen, L.M. Effect of Pressure, reconstituted RTE meat microbiota, and antimicrobials on survival and post-pressure growth of Listeria monocytogenes on ham. Front. Microbiol. 2018, 9, 1979. [Google Scholar] [CrossRef]
- Ramaroson, M.; Guillou, S.; Rossero, A.; Rezé, S.; Anthoine, V.; Moriceau, N.; Martin, J.-L.; Duranton, F.; Zagorec, M. Selection procedure of bioprotective cultures for their combined use with High Pressure Processing to control spore-forming bacteria in cooked ham. Int. J. Food Microbiol. 2018, 276, 28–38. [Google Scholar] [CrossRef]
- Modugno, C.; Kmiha, S.; Simonin, H.; Aouadhi, C.; Diosdado Cañizares, E.; Lang, E.; André, S.; Mejri, S.; Maaroufi, A.; Perrier-Cornet, J.M. High pressure sensitization of heat-resistant and pathogenic foodborne spores to nisin. Food Microbiol. 2019, 84, 103244. [Google Scholar] [CrossRef]
- Najjari, A.; Ouzari, H.; Boudabous, A.; Zagorec, M. Method for reliable isolation of Lactobacillus sakei strains originating from Tunisian seafood and meat products. Int. J. Food Microbiol. 2008, 121, 342–351. [Google Scholar] [CrossRef]
- Poirier, S.; Rué, O.; Peguilhan, R.; Coeuret, G.; Zagorec, M.; Champomier-Vergès, M.-C.; Loux, V.; Chaillou, S. Deciphering intra-species bacterial diversity of meat and seafood spoilage microbiota using gyrB amplicon sequencing: A comparative analysis with 16S rDNA V3-V4 amplicon sequencing. PLoS ONE 2018, 13, e0204629. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinforma. Oxf. Engl. 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. Peer J. 2014, 2, e593. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 December 2021).
- Raimondi, S.; Luciani, R.; Sirangelo, T.M.; Amaretti, A.; Leonardi, A.; Ulrici, A.; Foca, G.; D’Auria, G.; Moya, A.; Zuliani, V.; et al. Microbiota of sliced cooked ham packaged in modified atmosphere throughout the shelf life: Microbiota of sliced cooked ham in MAP. Int. J. Food Microbiol. 2019, 289, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Vasilopoulos, C.; De Maere, H.; De Mey, E.; Paelinck, H.; De Vuyst, L.; Leroy, F. Technology-induced selection towards the spoilage microbiota of artisan-type cooked ham packed under modified atmosphere. Food Microbiol. 2010, 27, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zagdoun, M.; Coeuret, G.; N’Dione, M.; Champomier-Vergès, M.-C.; Chaillou, S. Large microbiota survey reveals how the microbial ecology of cooked ham is shaped by different processing steps. Food Microbiol. 2020, 91, 103547. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhou, J.; Oren, A.; Zhang, Y.-Z. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Yamamoto, K. Food processing by high hydrostatic pressure. Biosci. Biotechnol. Biochem. 2017, 81, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Tassou, C.C.; Galiatsatou, P.; Samaras, F.J.; Mallidis, C.G. Inactivation kinetics of a piezotolerant Staphylococcus aureus isolated from high-pressure-treated sliced ham by high pressure in buffer and in a ham model system: Evaluation in selective and non-selective medium. Innov. Food Sci. Emerg. Technol. 2007, 8, 478–484. [Google Scholar] [CrossRef]
- Verheyen, D.; Govaert, M.; Seow, T.K.; Ruvina, J.; Mukherjee, V.; Baka, M.; Skåra, T.; Van Impe, J.F.M. The complex effect of food matrix fat content on thermal inactivation of Listeria monocytogenes: Case study in emulsion and gelled emulsion model systems. Front. Microbiol. 2020, 10, 3149. [Google Scholar] [CrossRef]
- La Storia, A.; Ferrocino, I.; Torrieri, E.; Di Monaco, R.; Mauriello, G.; Villani, F.; Ercolini, D. A combination of modified atmosphere and antimicrobial packaging to extend the shelf-life of beefsteaks stored at chill temperature. Int. J. Food Microbiol. 2012, 158, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Illikoud, N.; Gohier, R.; Werner, D.; Barrachina, C.; Roche, D.; Jaffrès, E.; Zagorec, M. Transcriptome and volatilome analysis during growth of Brochothrix thermosphacta in food: Role of food substrate and strain specificity for the expression of spoilage functions. Front. Microbiol. 2019, 10, 2527. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample Name | Sampling Date 1 | Additives 2 | Total Counts 3 | LAB Counts 4 |
|---|---|---|---|---|
| HAM_A | UBD + 6 12 February 2015 | Dextrose Sodium nitrite E250 Potassium chloride E508 Sodium erythorbate E316 Potassium lactate E326 Sodium ascorbate E301 Sodium acetate E262 Diphosphates E450 Triphosphates E451 Polyphosphates E452 | 8.6 ± 0.4 | 8.2 ± 0.0 |
| HAM_B 5 | UBD + 5 18 May 2016 | Sugar Lactose Nitrite salt Common salt Spices | 7.2 ± 0.7 | 7.3 ± 0.5 |
| HAM_C | UBD − 1 11 June 2015 | Dextrose Sodium nitrite E250 Potassium chloride E508 Potassium lactate E326 Sodium acetate E262 Sodium ascorbate E316 Polyphosphates E452 | 8.6 ± 0.1 | 9.0 ± 0.1 |
| HAM_D | UBD − 4 21 August 2015 | Sodium nitrite E250 Potassium chloride E508 Potassium lactate E326 Sodium diacetate E262 Sodium ascorbate E316 | 10.3 ± 0.1 | 8.1 ± 0.1 |
| Storage Time in Days 1 | ||||
|---|---|---|---|---|
| Sample Name | Day 1 | Day 5 | Day 12 | Day 30 |
| HAM_A samples | ||||
| No treatment | 4.63 ± 0.09 | 7.32 ± 0.14 | 7.84 ± 0.10 | 8.15 ± 0.15 |
| HPP | 1.20 ± 0.14 | 3.74 ± 0.01 | 3.00 ± 1.00 | 4.02 ± 0.59 |
| L. lactis | 7.21 ± 0.62 | 9.18 ± 0.02 | 8.39 ± 0.12 | 8.87 ± 0.87 |
| HPP + L. lactis | 2.54 ± 0.45 | 3.97 ± 0.26 | 5.42 ± 1.27 | 7.60 ± 0.03 |
| HAM_B samples | ||||
| No treatment | 5.24 ± 0.18 | 6.10 ± 0.13 | 6.90 ± 0.33 | 6.50 ± 0.20 |
| HPP | 1.39 ± 0.09 | ND 2 | 5.33 ± 0.34 | 6.23 ± 0.42 |
| L. lactis | 7.25 ± 0.51 | 8.29 ± 0.01 | 7.82 ± 0.21 | 8.50 ± 0.50 |
| HPP + L. lactis | 2.75 ± 0.44 | 3.64 ± 0.07 | 7.11 ± 0.13 | 10.2 ± 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaillou, S.; Ramaroson, M.; Coeuret, G.; Rossero, A.; Anthoine, V.; Champomier-Vergès, M.; Moriceau, N.; Rezé, S.; Martin, J.-L.; Guillou, S.; et al. Combination of High-Pressure Treatment at 500 MPa and Biopreservation with a Lactococcus lactis Strain for Lowering the Bacterial Growth during Storage of Diced Cooked Ham with Reduced Nitrite Salt. Microorganisms 2022, 10, 456. https://doi.org/10.3390/microorganisms10020456
Chaillou S, Ramaroson M, Coeuret G, Rossero A, Anthoine V, Champomier-Vergès M, Moriceau N, Rezé S, Martin J-L, Guillou S, et al. Combination of High-Pressure Treatment at 500 MPa and Biopreservation with a Lactococcus lactis Strain for Lowering the Bacterial Growth during Storage of Diced Cooked Ham with Reduced Nitrite Salt. Microorganisms. 2022; 10(2):456. https://doi.org/10.3390/microorganisms10020456
Chicago/Turabian StyleChaillou, Stéphane, Mihanta Ramaroson, Gwendoline Coeuret, Albert Rossero, Valérie Anthoine, Marie Champomier-Vergès, Nicolas Moriceau, Sandrine Rezé, Jean-Luc Martin, Sandrine Guillou, and et al. 2022. "Combination of High-Pressure Treatment at 500 MPa and Biopreservation with a Lactococcus lactis Strain for Lowering the Bacterial Growth during Storage of Diced Cooked Ham with Reduced Nitrite Salt" Microorganisms 10, no. 2: 456. https://doi.org/10.3390/microorganisms10020456