
Genomic Comparison, Phylogeny and Taxonomic Reevaluation of the Ectothiorhodospiraceae and Description of Halorhodospiraceae fam. nov. and Halochlorospira gen. nov.


Abstract
:1. Introduction
2. Material and Methods
2.1. Genome Sequences Used in this Study
2.2. Genomic DNA Extraction and Sequencing
2.3. Whole Genome Comparison
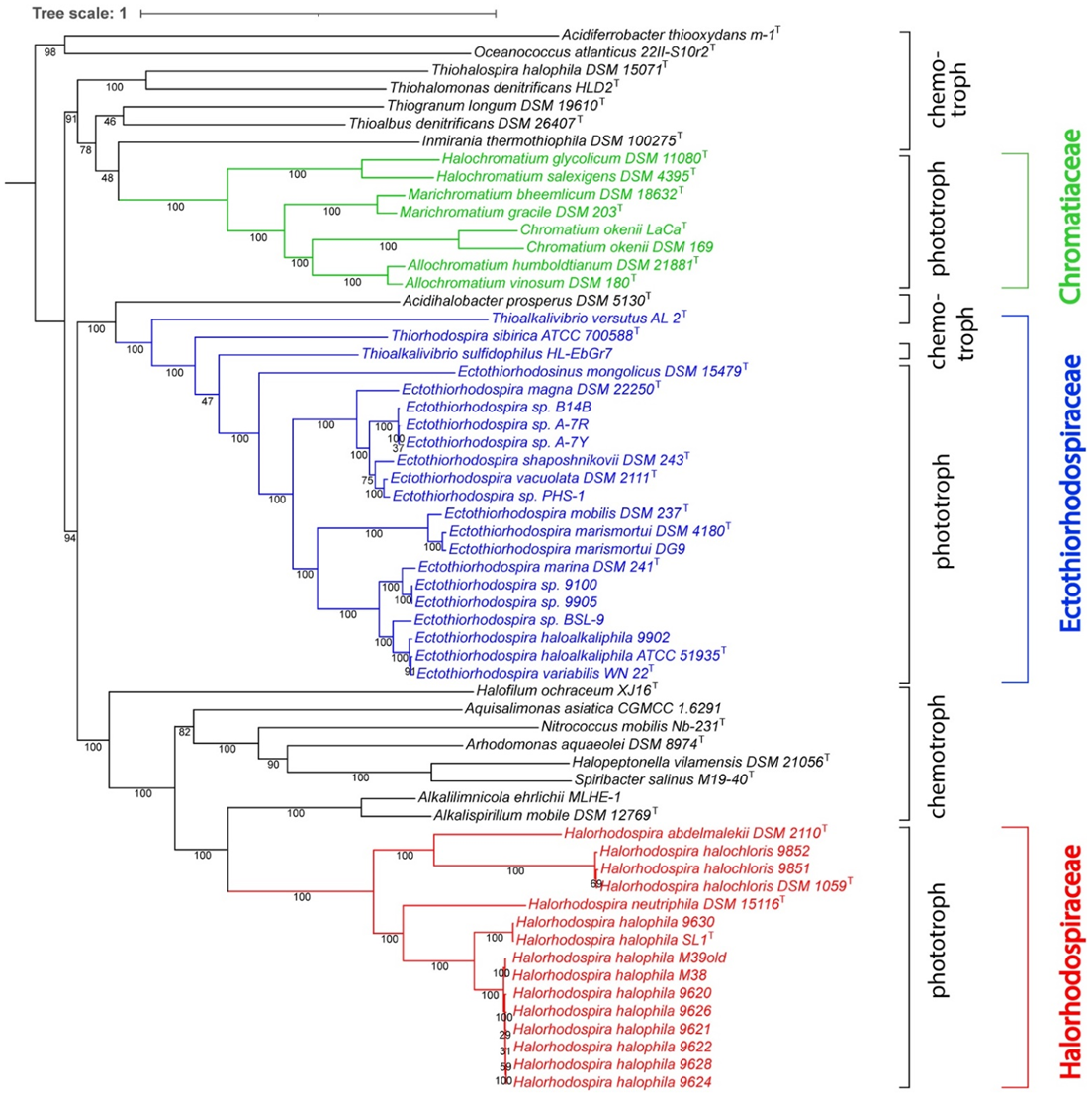
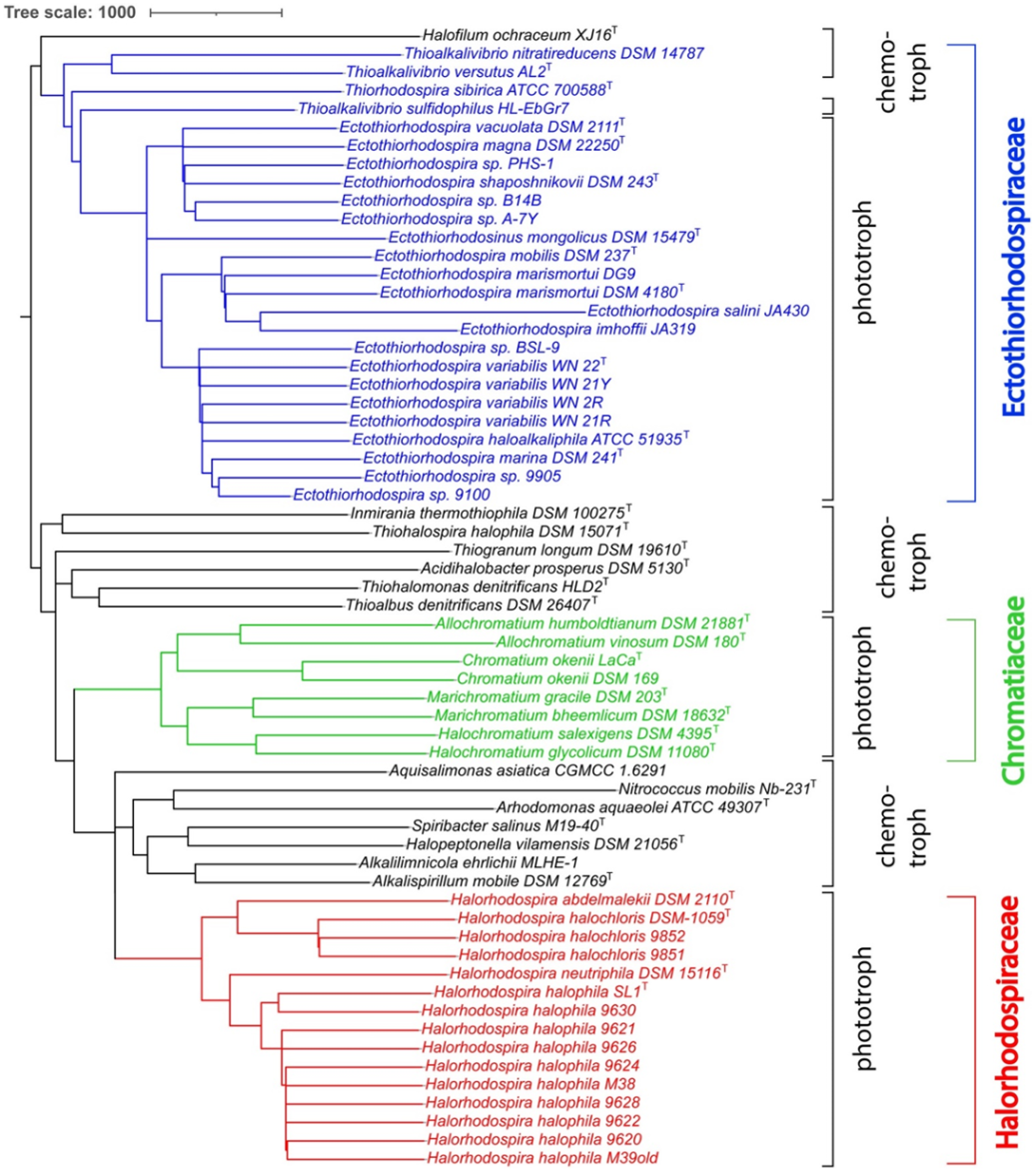
3. Results and Discussion
3.1. Genomes of Ectothiorhodospiraceae
3.2. Genomes of Halorhodospira Species
3.3. Genome-Delineated Properties
3.3.1. Sulfur and Thiosulfate Oxidation
3.3.2. Glutathione
3.3.3. Carboxysome Genes
3.3.4. Nitric Oxide Reduction
3.3.5. HIPIP
3.3.6. Cytochrome b5
3.3.7. Arsenic Oxidation
3.3.8. Photoactive Yellow Protein—PYP
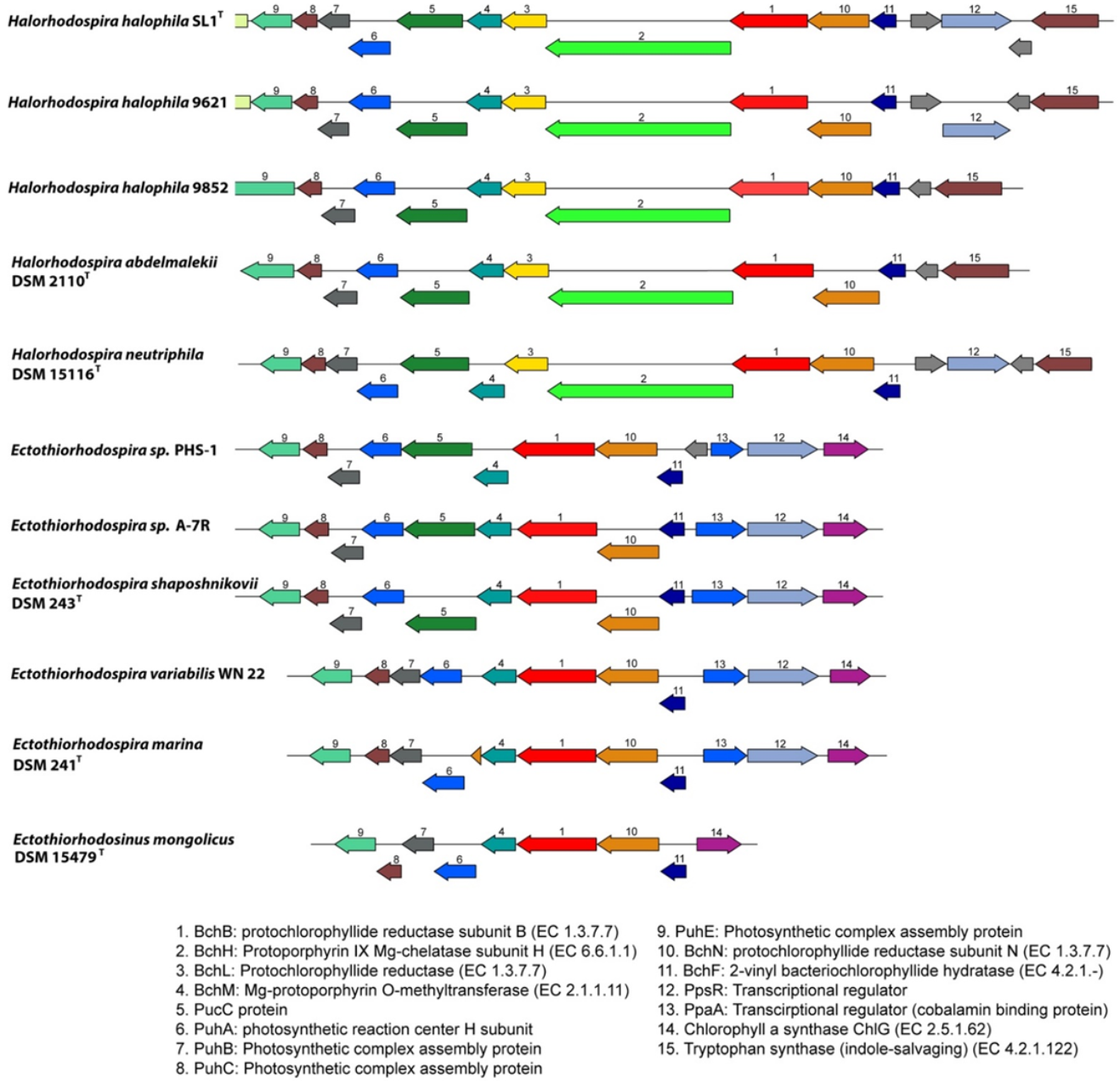
3.3.9. Photosynthesis Gene Clusters
3.4. Habitats and Environmental Distribution
4. Systematic Conclusions
5. Emended Description of the Family Ectothiorhodospiraceae Imhoff 1984a, 33VP
6. Description of the Family Halorhodospiraceae. fam. nov.
6.1. Emended Description of the Genus Halorhodospira
6.2. Description of the Genus Halochlorospira gen. nov.
6.3. Description of Halochlorospira halochloris comb. nov.
6.4. Descrition of Halochlorospira abdelmalekii comb. nov.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pfennig, N.; Trüper, H.G. Higher Taxa of the Phototrophic Bacteria. Int. J. Syst. Bacteriol. 1971, 21, 17–18. [Google Scholar] [CrossRef] [Green Version]
- Fowler, V.J.; Pfennig, N.; Schubert, W.; Stackebrandt, E. Towards a phylogeny of phototrophic purple sulfur bacteria? 16S rRNA oligonucleotide cataloguing of 11 species of Chromatiaceae. Arch. Microbiol. 1984, 139, 382–387. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Fowler, V.J.; Schubert, W.; Imhoff, J.F. Towards a phylogeny of phototrophic purple sulfur bacteria—The genus Ectothiorhodospira. Arch. Microbiol. 1984, 137, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F. Reassignment of the Genus Ectothiorhodospira Pelsh 1936 to a New Family, Ectothiorhodospiraceae fam. nov., and Emended Description of the Chromatiaceae Bavendamm 1924. Int. J. Syst. Bacteriol. 1984, 34, 338–339. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F. Family Ectothiorhodospiraceae. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, Part B; pp. 41–43. [Google Scholar]
- Imhoff, J.F. Genus Ectothiorhodospira. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, Part B, pp. 43–48. [Google Scholar]
- Imhoff, J.F. Genus Halorhodospira. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, Part B, pp. 49–52. [Google Scholar]
- Imhoff, J.F. The family Ectothiorhodospiraceae. In The Prokaryotes. An Evolving Electronic Resource for the Microbiological Community, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer Verlag: New York, NY, USA, 1999. [Google Scholar]
- Thiemann, B.; Imhoff, J.F. Differentiation of Ectothiorhodospiraceae Based on Their Fatty Acid Composition. Syst. Appl. Microbiol. 1996, 19, 223–230. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Bias-Imhoff, U. Lipids, Quinones and Fatty Acids of Anoxygenic PHOTOTROPHIC bacteria. In Anoxygenic Photosynthetic Bacteria; Blankenship, R.E., Madigan, M.T., Bauer, C.E., Eds.; Kluwer Academic Publ.: Dordrecht, The Netherlands, 1995; pp. 179–205. [Google Scholar]
- Zahr, M.; Fobel, B.; Mayer, H.; Imhoff, J.F.; Campos, V.P.; Weckesser, J. Chemical composition of the lipopolysaccharides of Ectothiorhodospira shaposhnikovii, Ectothiorhodospira mobilis and Ectothiorhodospira halophila. Arch. Microbiol. 1992, 157, 499–504. [Google Scholar]
- Weckesser, J.; Drews, G.; Mayer, H. Lipopolysaccharides of photosynthetic bacteria. Ann. Rev. Microbiol. 1979, 33, 215–239. [Google Scholar] [CrossRef]
- Weckesser, J.; Mayer, H.; Schulz, G. Anoxygenic Phototrophic Bacteria: Model Organisms for Studies on Cell Wall Macromolecules. In Anoxygenic Photosynthetic Bacteria; Blankenship, R.E., Madigan, M.T., Bauer, C.E., Eds.; Kluwer Academic Publ.: Dordrecht, The Netherlands, 1995; pp. 207–230. [Google Scholar]
- Imhoff, J.F.; Süling, J. The phylogenetic relationship among Ectothiorhodospiraceae: A reevaluation of their taxonomy on the basis of 16S rDNA analyses. Arch. Microbiol. 1996, 165, 106–113. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase. Microorganisms 2019, 7, 576. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Photosynthesis Is Widely Distributed among Proteobacteria as Demonstrated by the Phylogeny of PufLM Reaction Center Proteins. Front. Microbiol. 2018, 8, 2679. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Keller, A.; Neulinger, S.C. Osmotic Adaptation and Compatible Solute Biosynthesis of Phototrophic Bacteria as Revealed from Genome Analyses. Microorganisms 2019, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Han, J.; Lapidus, A.; Cheng, J.F.; Goodwin, L.; Pitluck, S.; Peters, L.; Land, M.L.; Hauser, L.; Vogl, K.; et al. The draft genome of Thiorhodospira sibirica ATCC 700588. 2011; unpublished. [Google Scholar]
- Saltikov, C.W.; Zargar, K.; Conrad, A.; Bernick, D.; Lowe, T.M.; Stolc, V.; Hoeft, S.; Oremland, R.S.; Stolz, J. Ectothiorhodospira sp. PHS-1, whole genome shotgun sequence. 2012; unpublished. [Google Scholar]
- Bryant, D.A.; Huntemann, M.; Han, J.; Chen, A.; Kyrpides, N.; Mavromatis, K.; Markowitz, V.; Palaniappan, K.; Ivanova, N.; Schaumberg, A.; et al. Ectothiorhodospira haloalkaliphila ATCC 51935, whole genome shotgun sequence. DOE Joint Genome Institute. 2013; unpublished. [Google Scholar]
- Hernandez-Maldonado, J.; Stoneburner, B.; Boren, A.; Miller, L.; Rosen, M.; Oremland, R.S.; Saltikov, C.W. Genome sequence of the photoarsenotrophic bacterium Ectothiorhodospira sp. strain BSL-9, isolated from a hypersaline alkaline arsenic-rich extreme environment. Genome Announc. 2016, 4, e01139-16. [Google Scholar] [CrossRef] [Green Version]
- Challacombe, J.F.; Majid, S.; Deole, R.; Brettin, T.S.; Bruce, D.; Delano, S.F.; Detter, J.C.; Gleasner, C.D.; Han, C.S.; Misra, M.; et al. Complete genome sequence of Halorhodospira halophila SL1. Stand. Genom. Sci. 2013, 8, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Tsukatani, Y.; Hirose, Y.; Harada, J.; Yonekawa, C.; Tamiaki, H. Unusual features in the photosynthetic machinery of Halorhodospira halochloris DSM 1059 revealed by complete genome sequencing. Photosynth. Res. 2019, 140, 311–319. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Muntyan, M.S.; Pantaleeva, A.N.; Muyzer, G. Thioalkalivibrio sulfidophilus sp. nov., a haloalkaliphilic, sulfur-oxidizing gammaproteobacterium from alkaline habitats. Int. J. Syst. Evol. Microbiol. 2012, 62, 1884–1889. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F.; Hashwa, F.; Trüper, H.G. Isolation of extremely halophilic phototrophic bacteria from the alkaline Wadi Natrun, Egypt. Arch. Hydrobiol. 1978, 84, 381–388. [Google Scholar]
- Gorlenko, V.M.; Bryantseva, I.A.; Rabold, S.; Tourova, T.P.; Rubtsova, D.; Smirnova, E.; Thiel, V.; Imhoff, J.F. A novel alkaliphilic and halophilic purple sulfur bacterium Ectothiorhodospira variabilis from soda lakes. Int. J. Syst. Evol. Microbiol. 2009, 59, 658–664. [Google Scholar] [CrossRef]
- Ventura, S.; Viti, C.; Pastorelli, R.; Giovannetti, L. Revision of species delineation in the genus Ectothiorhodospira. Int. J. Syst. Evol. Microbiol. 2000, 50, 583–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, S.; Giovanetti, L.; Gori, A.; Viti, C.; Materassi, R. Total DNA restriction pattern and quinone composition of members of the family Ectothiorhodospiraceae. System. Appl. Microbiol. 1993, 16, 405–410. [Google Scholar] [CrossRef]
- Tourova, T.P.; Spiridonova, E.M.; Berg, I.; Slobodova, N.V.; Boulygina, E.S.; Sorokin, D.Y. Phylogeny and evolution of the family Ectothiorhodospiraceae based on comparison of 16S rRNA, cbbL and nifH gene sequences. Int. J. Syst. Evol. Microbiol. 2007, 57, 2387–2398. [Google Scholar] [CrossRef] [PubMed]
- McCann, S.H.; Boren, A.; Hernandez-Maldonado, J.; Stoneburner, B.; Saltikov, C.W.; Stolz, J.F.; Oremland, R.S. Arsenite as an Electron Donor for Anoxygenic Photosynthesis: Description of Three Strains of Ectothiorhodospira from Mono Lake, California and Big Soda Lake, Nevada. Life 2016, 7, E1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlenko, V.M.; Burganskaya, E.I.; Bryantseva, I.A. Phototrophic communities of the Berikei highly mineralized mesothermal sulfide springs (Dagestan, Russia). Microbiology 2019, 88, 146–155. [Google Scholar] [CrossRef]
- Gorlenko, V.M.; Bryantseva, I.A.; Panteleeva, E.E.; Turova, T.P.; Kolganova, T.V.; Makhneva, Z.K.; Moskalenko, A.A. Ectothio-rhodosinus mongolicum gen. nov., sp. nov., a new purple sulfur bacterium from a soda lake in Mongolia. Microbiology (Moscow) 2003, 73, 66–73. [Google Scholar] [CrossRef]
- Bryantseva, I.A.; Gorlenko, V.M.; Kompantseva, E.I.; Imhoff, J.F.; Süling, J.; Mityushina, L. Thiorhodospira sibirica gen. nov., sp. nov., a new alkaliphilic purple sulfur bacterium from a Siberian soda lake. Int. J. Syst. Evol. Microbiol. 1999, 49, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Bryantseva, I.A.; Tourova, T.P.; Kovaleva, O.L.; Kostrikina, N.A.; Gorlenko, V.M. Ectothiorhodospira magna sp. nov., a new large alkaliphilic purple sulfur bacterium. Microbiology 2010, 79, 780–790. [Google Scholar] [CrossRef]
- Raymond, J.C.; Sistrom, W.R. The isolation and preliminary characterization of a halophilic photosynthetic bacterium. Arch. Microbiol. 1967, 59, 255–268. [Google Scholar] [CrossRef]
- Raymond, J.C.; Sistrom, W.R. Ectothiorhodospira halophila: A new species of the genus Ectothiorhodospira. Archiv. Mikrobiol. 1969, 69, 121–126. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Trüper, H.G. Ectothiorhodospira halochloris sp. nov., a new extremely halophilic phototrophic bacterium containing bacteriochlorophyll b. Arch. Microbiol. 1977, 114, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F.; Truper, H.G. Ectothiorhodospira abdelmalekii sp. nov., a new halophilic and alkaliphilic phototrophic bacterium. Zentbl. Bakteriol. 1981, 2, 228–234. [Google Scholar] [CrossRef]
- Hirschler-Réa, A.; Matheron, R.; Riffaud, C.; Mouné, S.; Eatock, C.; Herbert, R.A.; Willison, J.C.; Caumette, P. Isolation and characterization of spirilloid purple phototrophic bacteria forming red layers in microbial mats of Mediterranean salterns: Description of Halorhodospira neutriphila sp. nov. and emendation of the genus Halorhodospira. Int. J. Syst. Evol. Microbiol. 2003, 53, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F.; Sahl, H.G.; Soliman, G.S.; Trüper, H.G. The Wadi Natrun: Chemical composition and microbial mass developments in alkaline brines of Eutrophic Desert Lakes. Geomicrobiol. J. 1979, 1, 219–234. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Singh, K.S.; Kirksey, J.; Hoff, W.D.; Deole, R. Draft Genome Sequence of the Extremely Halophilic Phototrophic Purple Sulfur Bacterium Halorhodospira halochloris. J. Genom. 2014, 2, 118–120. [Google Scholar] [CrossRef] [Green Version]
- Brune, D.C. Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina. Arch. Microbiol. 1995, 163, 391–399. [Google Scholar] [CrossRef]
- Pattaragulwanit, K.; Brune, D.C.; Trüper, H.G.; Dahl, C. Molecular genetic evidence for extracytoplasmic localization of sulfur globules in Chromatium vinosum. Arch. Microbiol. 1998, 169, 434–444. [Google Scholar] [CrossRef]
- Dahl, C.; Franz, B.; Hensen, D.; Kesselheim, A.; Zigann, R. Sulfite oxidation in the purple sulfur bacterium Allochromatium vinosum: Identification of SoeABC as a major player and relevance of SoxYZ in the process. Microbiology 2013, 159, 2626–2638. [Google Scholar] [CrossRef] [Green Version]
- Dahl, C. Cytoplasmic sulfur trafficking in sulfur-oxidizing prokaryotes. IUBMB Life 2015, 67, 268–274. [Google Scholar] [CrossRef]
- Fahey, R.C. Novel Thiols of Prokaryotes. Annu. Rev. Microbiol. 2001, 55, 333–356. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, R.G.; Newton, G.L.; Sherrill, C.; Fahey, R.C. Glutathione amide and its perthiol in anaerobic sulfur bacteria. J. Bacteriol. 1996, 78, 4742–4746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Petegem, F.; De Vos, D.; Savvides, S.; Vergauwen, B.; van Beeumen, J. Understanding nicotinamide dinucleotide cofactor and substrate specificity in class I flavoprotein disulfide oxidoreductases: Crystallographic analysis of a glutathione amide reductase. J. Mol. Biol. 2007, 374, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Kerfeld, C.A.; Melnicki, M.R. Assembly, function and evolution of cyanobacterial carboxysomes. Curr. Opin. Plant Biol. 2016, 31, 66–75. [Google Scholar] [CrossRef] [Green Version]
- MacCready, J.S.; Tran, L.; Basalla, J.L.; Hakim, P.; Vecchiarelli, A.G. The McdAB system positions α-carboxysomes in proteobacteria. Mol. Microbiol. 2021, 116, 277–297. [Google Scholar] [CrossRef]
- Venturoli, G.; Mamedov, M.D.; Mansy, S.S.; Musiani, F.; Strocchi, M.; Francia, F.; Semenov, A.Y.; Cowan, J.A.; Ciurli, S. Electron transfer from HiPIP to the photooxidized tetraheme cytochrome subunit of Allochromatium vinosum reaction center: New insights from site-directed mutagenesis and computational studies. Biochemistry 2004, 43, 437–445. [Google Scholar] [CrossRef]
- Nagashima, K.V.P.; Matsuura, K.; Shimada, K.; Verméglio, A. High-Potential Iron−Sulfur Protein (HiPIP) Is the Major Electron Donor to the Reaction Center Complex in Photosynthetically Growing Cells of the Purple Bacterium Rubrivivax gelatinosus. Biochemistry 2002, 41, 14028–14032. [Google Scholar] [CrossRef]
- Przysiecki, C.T.; Meyer, T.E.; Cusanovich, M.A. Circular dichroism and redox properties of high redox potential ferredoxins. Biochemistry 1985, 24, 2542–2549. [Google Scholar] [CrossRef]
- Lieutaud, C.; Alric, J.; Bauzan, M.; Nitschke, W.; Schoepp-Cothenet, B. Study of the high-potential iron sulfur protein in Halo-rhodospira halophila confirms that it is distinct from cytochrome c as electron carrier. Proc. Natl. Acad. Sci. USA 2005, 102, 3260–3265. [Google Scholar] [CrossRef] [Green Version]
- Azai, C.; Tsukatani, Y.; Itoh, S.; Oh-Oka, H. C-type cytochromes in the photosynthetic electron transfer pathways in green sulfur bacteria and heliobacteria. Photosynth. Res. 2010, 104, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Kostanjevecki, V.; Leys, D.; van Driessche, G.; Meyer, T.E.; Cusanovich, M.A.; Fischer, U.; Guisez, Y.; van Beeumen, J. Structure and characterization of Ectothiorhodospira vacuolata cytochrome b(558), a prokaryotic homologue of cytochrome b(5). J. Biol. Chem. 1999, 274, 35614–35620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, K.; Schicklberger, M.; Gescher, J. Dissimilatory reduction of extracellular electron acceptors in anaerobic respiration. Appl. Environ. Microbiol. 2012, 78, 913–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, T.E.; Kyndt, J.A.; Memmi, S.; Moser, T.; Colón-Acevedo, B.; Devreese, B.; Van Beeumen, J.J. The growing family of photoactive yellow proteins and their presumed functional roles. Photochem. Photobiol. Sci. 2012, 11, 1495–1514. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, J.A.; Fitch, J.C.; Meyer, T.E.; Cusanovich, M.A. The Photoactivated PYP Domain of Rhodospirillum centenum Ppr Accelerates the Recovery of the Bacteriophytochrome Domain after White Light Illumination. Biochemistry 2007, 46, 8256–8262. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, J.A.; Fitch, J.C.; Meyer, T.E.; Cusanovich, M.A. Thermochromatium tepidum Photoactive Yellow Protein/Bacteriophytochrome/Diguanylate Cyclase: Characterization of the PYP Domain. Biochemistry 2005, 44, 4755–4764. [Google Scholar] [CrossRef]
- Tourova, T.P.; Slobodova, N.V.; Bumazhkin, B.K.; Kolganova, T.V.; Muyzer, G.; Sorokin, D.Y. Analysis of community composition of sulfur-oxidizing bacteria in hypersaline and soda lakes using soxB as a functional marker. FEMS Microbiol. Ecol. 2013, 84, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Gorlenko, V.M.; Namsaraev, B.B.; Namsaraev, Z.B.; Lysenko, A.M.; Eshinimaev, B.T.; Khmelenina, V.N.; Trotsenko, Y.A.; Kuenen, J.G. Prokaryotic communities of the north-eastern Mongolian soda lakes. Hydrobiologia 2004, 522, 235–248. [Google Scholar] [CrossRef]
- Imhoff, J.F. Anoxygenic Phototrophic Bacteria from Extreme Environments. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 427–480. [Google Scholar]
- Jannasch, H.W. Die bakterielle Rotfärbung der Salzseen des Wadi Natrun. Arch. Hydrobiol. 1957, 53, 425–433. [Google Scholar] [CrossRef]
- Mesbah, N.M.; Abou-El-Ela, S.H.; Wiegel, J. Novel and unexpected prokaryotic diversity in water and sediments of the alkaline, hypersaline lakes of the Wadi an Natrun, Egypt. Microbiol. Ecol. 2007, 54, 598–617. [Google Scholar] [CrossRef]
- Tank, M.; Thiel, V.; Imhoff, J.F. Phylogenetic relationship of phototrophic purple sulfur bacteria according to pufL and pufM genes. Int. Microbiol. 2009, 12, 175–185. [Google Scholar]
- Thiel, V.; Tank, M.; Neulinger, S.C.; Gehrmann, L.; Dorador, C.; Imhoff, J.F. Unique communities of anoxygenic phototrophic bacteria in saline lakes of Salar de Atacama (Chile). Evidence for a new phylogenetic lineage of phototrophic Gammaproteobacteria from pufLM gene analyses. FEMS Microbiol. Ecol. 2010, 74, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, K.B.; Hedrich, S.; Johnson, D.B. Acidiferrobacter thiooxydans, gen. nov. sp. nov.; an acidophilic, thermo-tolerant, facultatively anaerobic iron- and sulfur-oxidizer of the family Ectothiorhodospiraceae. Extremophiles 2011, 15, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Shinohara, A.; Fukui, M. Sulfurifustis variabilis gen. nov., sp. nov., a sulfur oxidizer isolated from a lake, and proposal of Acidiferrobacteraceae fam. nov. and Acidiferrobacterales ord. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 3709–3713. [Google Scholar] [CrossRef]
- Issotta, F.; Moya-Beltran, A.; Mena, C.; Covarrubias, P.C.; Thyssen, C.; Bellenberg, S.; Sand, W.; Quatrinia, R.; Vera, M. Insights into the biology of acidophilic members of the Acidiferrobacteraceae family derived from comparative genomic analyses. Res. Microbiol. 2018, 169, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lai, Q.; Liu, X.; Sun, F.; Du, Y.; Li, G.; Shao, Z. Maricoccus atlantica gen. nov. sp. nov., isolated from deep sea sediment of the Atlantic Ocean. Antonie van Leeuwenhoek 2013, 104, 1073–1081, Erratum in Antonie van Leeuwenhoek 2014, 105, 439. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Zhao, J.-X.; Sang, J.; Chen, G.-J.; Du, Z.-J. Halofilum ochraceum gen. nov., sp. nov., a gammaproteobacterium isolated from a marine solar saltern. Int. J. Syst. Evol. Microbiol. 2017, 67, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; van Pelt, S.; Tourova, T.P.; Takaichi, S.; Muyzer, G. Acetonitrile degradation under haloalkaline conditions by Natronocella acetinitrilica gen. nov., sp.no. Microbiology 2007, 153, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Trotsenko, Y.A.; Doronina, N.V.; Tourova, T.P.; Galinski, E.A.; Kolganova, T.V.; Muyzer, G. Methylohalomonas lacus gen. nov., sp. nov. and Methylonatrum kenyenese gen. nov., sp. nov., methylotrophic Gammaproteobacteria from hypersaline lakes. Int. J. Syst. Evol. Microbiol. 2007, 57, 2762–2769. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Suzuki, K.-I.; Urabe, T.; Sugihara, M.; Tanaka, K.; Hamada, M.; Hanada, S. Thioprofunndum hispidum sp. nov., an obligately chemolithoautotrophic sulfur-oxidizing gammaproteobacterium isolated from the hydrothermal field on Suiyo Seamount, and proposal of Thioalkalispiraceae fam. nov. in the order Chromatiales. Int. J. Syst. Evol. Microbiol. 2011, 61, 2412–2418. [Google Scholar] [CrossRef] [Green Version]
- Barco, R.A.; Garrity, G.M.; Scott, J.J.; Amend, J.P.; Nealson, K.H.; Emerson, D. A genus definition for Bacteria and Archaea based on a standard genome relatedness index. mBio 2020, 11, e02475-19. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Guan, Y.; Li, F.; Zhao, Y.; Feng, C.; Tang, N. Biomass and pigments production of a newly isolated photosynthetic bacterium Ectothiorhodospira shaposhnikovii P2 from saline wastewater. Int. J. Environ. Sci. Technol. 2018, 16, 7487–7496. [Google Scholar] [CrossRef]
- Ramana, V.V.; Sasikala, C.; Ramaprasad, E.V.V.; Ramana, C.V. Description of Ectothiorhodospira salini sp. nov. J. Gen. Appl. Microbiol. 2010, 56, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Genome Name | Strain | Alternate Strain | Type Strain | GenBank Accessions | Contigs | Genome Length | GC mol% | quinone | CDS | Coarse Consis-tency | Fine Consis-tency | Sequencing Depth | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ectothiorhodospira haloalkaliphila | ATCC 51935 | yes | AJUE00000000 | 36 | 3,445,226 | 63.0 | MK-7/Q-8 | 3155 | 95.3 | 93.9 | Unknown | [20] | |
| Ectothiorhodospira haloalkaliphila | 9902 | strain C | no | JAJNQR000000000 | 243 | 3,666,279 | 62.7 | 3602 | 95.4 | 93.7 | 106x | this study | |
| Ectothiorhodospira variabilis | DSM 21381 | WN 22 | yes | JAJNAO000000000 | 84 | 3,416,336 | 63.0 | 3250 | 94.3 | 92.8 | 119x | this study | |
| Ectothiorhodospira variabilis | WN 21Y | no | JAJNAP000000000 | 84 | 3,452,695 | 63.0 | 3302 | 94.3 | 92.9 | 145x | this study | ||
| Ectothiorhodospira variabilis | WN 21R | no | JAJNAQ000000000 | 71 | 3,484,384 | 63.0 | 3341 | 94.3 | 92.9 | 119x | this study | ||
| Ectothiorhodospira variabilis | WN 2R | no | JAJNAR000000000 | 73 | 3,484,893 | 63.0 | 3341 | 94.3 | 92.9 | 132x | this study | ||
| Ectothiorhodospira sp. | BSL-9 | no | CP011994 | 1 | 3,550,080 | 63.0 | 3350 | 95.7 | 93.1 | 298x | [21] | ||
| Ectothiorhodospira sp. | 9100 | YC6.1 | no | JAJNAN000000000 | 161 | 3,238,183 | 62.3 | 3169 | 93.3 | 92 | 103x | this study | |
| Ectothiorhodospira sp. | 9905 | no | JAJNAM000000000 | 180 | 3,202,442 | 62.3 | 3120 | 94.8 | 93.4 | 52x | this study | ||
| Ectothiorhodospira marina | DSM 241 | 9914 | yes | FOAA00000000 | 41 | 3,185,852 | 62.3 | MK-7/Q-8 | 3059 | 94.8 | 93.9 | 378x | [17] |
| Ectothiorhodospira mobilis | DSM 237 | yes | NRSK01000000 | 119 | 2,796,053 | 68.3 | MK-7/Q-8 | 2714 | 95.4 | 93.1 | 539x | [17] | |
| Ectothiorhodospira marismortui | DSM 4180 | 9410 | yes | FOUO00000000 | 35 | 2,624,954 | 68.2 | MK-7/Q-8 | 2474 | 94.6 | 93.7 | 363x | [17] |
| Ectothiorhodospira marismortui | DG9 | no | JAJOZD000000000 | 40 | 2,682,160 | 68.4 | 2522 | 94.7 | 93.7 | 139x | this study | ||
| Ectothiorhodospira vacuolata | DSM 2111 | Beta 1 | yes | JAJMLZ000000000 | 40 | 3,341,893 | 63.4 | MK-7/Q-7 | 3100 | 93.7 | 92.5 | 125x | this study |
| Ectothiorhodospira sp. | PHS-1 | no | AGBG00000000 | 114 | 2,943,210 | 63.7 | 2725 | 95.6 | 94.2 | 42x | [19] | ||
| Ectothiorhodospira shaposhnikovii | DSM 243 | 9710 | yes | NRSM01000000 | 154 | 3,788,226 | 62.4 | MK-7/Q-7 | 3743 | 96 | 93.9 | 160x | [17] |
| Ectothiorhodospira sp. | A-7R | no | JAJNQO000000000 | 47 | 3,282,361 | 63.0 | 3009 | 96.1 | 94.4 | 84x | this study | ||
| Ectothiorhodospira sp. | A-7Y | no | JAJNQN000000000 | 50 | 3,282,916 | 63.0 | 3019 | 95.9 | 94.3 | 52x | this study | ||
| Ectothiorhodospira sp. | B14B | no | JAJNQM000000000 | 164 | 3,422,169 | 62.9 | 3209 | 95.5 | 93 | 10x | this study | ||
| Ectothiorhodospira magna | DSM 22250 | B7-7 | yes | FOFO00000000 | 58 | 2,721,342 | 60.9 | 2559 | 94.7 | 93.6 | 438x | [17] | |
| Ectothiorhodosinus mongolicus | DSM15479 | M9 | yes | FTPK00000000 | 6 | 1,990,961 | 55.6 | 1867 | 95.2 | 94.5 | 593x | [17] | |
| Thiorhodospira sibirica | ATCC 700588 | yes | AGFD00000000 | 186 | 3,188,869 | 56.7 | 2842 | 93 | 92.1 | 30x | [18] | ||
| Halorhodospira abdelmalekii | DSM 2110 | yes | NRRN01000000 | 158 | 3,090,835 | 62.9 | MK-4 | 2914 | 94.4 | 93.2 | 122x | [17] | |
| Halorhodospira halochloris | DSM 1059 | 9850 | yes | AP017372 | 1 | 2,819,782 | 55.8 | MK-4 | 2699 | 94.4 | 93.7 | 123x | [23] |
| Halorhodospira halochloris | 9851 | no | JAJNRA000000000 | 141 | 2,883,546 | 56.1 | MK-4 | 2847 | 94.1 | 93.3 | 136x | this study | |
| Halorhodospira halochloris | 9852 | no | JAJNRB000000000 | 188 | 2,888,425 | 56.1 | 2861 | 93.6 | 92.8 | 91x | this study | ||
| Halorhodospira neutriphila | DSM 15116 | yes | NRSH01000000 | 287 | 2,394,163 | 72.0 | 2535 | 95.5 | 92.3 | 46x | [17] | ||
| Halorhodospira halophila | DSM 244 | SL1 | yes | CP000544 | 1 | 2,678,452 | 68.0 | MK-8 | 2414 | 99.7 | 99.3 | Unknown | [22] |
| Halorhodospira halophila | 9630 | no | NRRO00000000 | 45 | 2,682,427 | 67.9 | MK-8 | 2522 | 99.6 | 98.9 | 220x | [17] | |
| Halorhodospira halophila | 9620 | no | NHSE01000000 | 99 | 2,703,685 | 68.4 | 2600 | 99.2 | 98.3 | 49x | [17] | ||
| Halorhodospira halophila | 9621 | no | JAJNRD000000000 | 87 | 2,912,850 | 67.9 | 2705 | 99.6 | 99 | 70x | this study | ||
| Halorhodospira halophila | 9622 | no | JAJNRE000000000 | 121 | 2,909,722 | 67.9 | 2776 | 99.6 | 99 | 80x | this study | ||
| Halorhodospira halophila | 9624 | no | JAJNRC000000000 | 20 | 2,779,891 | 68.1 | MK-8 | 2599 | 97.6 | 96.6 | 117x | this study | |
| Halorhodospira halophila | 9626 | no | NHSN01000000 | 45 | 2,857,375 | 68.2 | 2720 | 99.5 | 98.9 | 72x | [17] | ||
| Halorhodospira halophila | 9628 | no | JAJNRF000000000 | 27 | 2,787,144 | 68.1 | 2598 | 99.4 | 99 | 68x | this study | ||
| Halorhodospira halophila | M38 | no | JAJNQP000000000 | 101 | 2,748,046 | 68.3 | 2572 | 99.4 | 98.8 | 143x | this study | ||
| Halorhodospira halophila | M39old | no | JAJNQQ000000000 | 105 | 2,747,719 | 68.3 | 2571 | 99.4 | 98.8 | 135x | this study |
| Ect. vacuolata DSM 2111 | |||||||||||||||||||
| 96.3 | Ect. PHS-1 | ||||||||||||||||||
| 90.5 | 90.3 | Ect. shaposhnikovii DSM 243 | |||||||||||||||||
| 89.4 | 89.1 | 88.1 | Ect. B14B | ||||||||||||||||
| 89.2 | 88.7 | 87.8 | 99 | Ect. A-7Y | |||||||||||||||
| 88.9 | 88.7 | 87.8 | 99.1 | 99.9 | Ect. A-7R | ||||||||||||||
| 86.2 | 85.9 | 84.6 | 83.4 | 83.3 | 83.6 | Ect. magna DSM 22250 | |||||||||||||
| 76.8 | 76.8 | 76.5 | 76.4 | 76.4 | 76.5 | 76 | Ect. BSL-9 | ||||||||||||
| 76.7 | 76.6 | 76.4 | 76 | 76.1 | 76.3 | 75.9 | 91.5 | Ect. haloalkaliphila ATCC 51935 | |||||||||||
| 76.5 | 76.5 | 76.2 | 76 | 76.1 | 76.1 | 76 | 91.3 | 98 | Ect. haloalkaliphila 9902 | ||||||||||
| 76.6 | 76.6 | 76.6 | 75.9 | 76 | 76.2 | 75.8 | 91.2 | 98.3 | 98.2 | Ect. variabilis WN 22 | |||||||||
| 76.1 | 76.1 | 76 | 75.7 | 75.9 | 76.1 | 75.5 | 86.9 | 86.8 | 86.8 | 86.6 | Ect. marina DSM 241 | ||||||||
| 76 | 76.3 | 75.6 | 75.5 | 75.7 | 75.9 | 75.5 | 87.3 | 86.9 | 86.8 | 86.7 | 96.7 | Ect. YC6.1 | |||||||
| 75.9 | 76.1 | 75.7 | 75.7 | 75.7 | 75.8 | 75.5 | 87.2 | 86.8 | 86.8 | 86.7 | 96.8 | 99.9 | Ect. 9905 | ||||||
| 76 | 75.8 | 75.4 | 75.1 | 75 | 75.3 | 74.9 | 76.7 | 76.2 | 76.3 | 75.9 | 76.1 | 75.9 | 75.9 | Ect. mobilis DSM 237 | |||||
| 75.9 | 75.8 | 75.4 | 75 | 75 | 75.3 | 74.9 | 76.7 | 76.4 | 76.4 | 76.1 | 76.1 | 76 | 76 | 92.6 | Ect. marismortui DSM 4180 | ||||
| 75.5 | 75.7 | 75 | 75 | 75 | 75.3 | 74.9 | 76.2 | 75.9 | 76.2 | 75.9 | 76 | 76 | 76 | 92.8 | 98 | Ect. marismortui DG9 | |||
| 70 | 70 | 69.8 | 69.9 | 69.8 | 69.9 | 69.9 | 70.2 | 70.2 | 70.1 | 70.1 | 70.1 | 70 | 70 | 69.5 | 69.3 | 69.5 | Ets. mongolicus DSM 15479 | ||
| 69.9 | 69.8 | 69.8 | 69.7 | 69.5 | 69.7 | 70 | 69.5 | 69.3 | 69.3 | 69.3 | 69.2 | 69.5 | 69.4 | 69.5 | 69.3 | 69.2 | 68.3 | Trs. siberica ATCC700588 | |
| Hlr. halophila SL1 | |||||||||||||||
| 99.99 | Hlr. halophila BN9630 | ||||||||||||||
| 85.8 | 85.8 | Hlr. halophila BN9626 | |||||||||||||
| 85.8 | 85.8 | 98 | Hlr. halophila BN9620 | ||||||||||||
| 85.7 | 85.7 | 98.5 | 98.1 | Hlr. halophila BN9628 | |||||||||||
| 85.7 | 85.7 | 98.5 | 98.1 | 100 | Hlr. halophila BN9624 | ||||||||||
| 85.7 | 85.7 | 98.1 | 98.5 | 98.4 | 98.4 | Hlr. halophila BN9622 | |||||||||
| 85.7 | 85.7 | 98.2 | 97.9 | 98.7 | 98.7 | 98.1 | Hlr. halophila BN9621 | ||||||||
| 85.6 | 85.5 | 97.4 | 97.9 | 97.4 | 97.4 | 98 | 97.3 | Hlr. halophila M39old | |||||||
| 85.6 | 85.5 | 97.3 | 97.8 | 97.4 | 97.4 | 98 | 97.3 | 100 | Hlr. halophila M38 | ||||||
| 77.4 | 77.4 | 77.4 | 77.4 | 77.6 | 77.6 | 77.6 | 77.6 | 77.7 | 77.6 | Hlr. neutriphila DSM 15116 | |||||
| 73.3 | 73.3 | 73.7 | 73.7 | 73.6 | 73.6 | 73.6 | 73.5 | 73.4 | 73.4 | 74.2 | Hlr. abdelmalekii BN9840 | ||||
| 70.6 | 70.5 | 70.9 | 71.1 | 71.2 | 71.2 | 71.3 | 71 | 70.8 | 70.8 | 70.7 | 72.7 | Hlr. halochloris BN9852 | |||
| 70.6 | 70.5 | 70.7 | 70.9 | 71 | 71 | 71 | 70.2 | 70.7 | 70.8 | 70.7 | 72.5 | 97.6 | Hlr. halochloris BN9850 | ||
| 70.5 | 70.4 | 70.7 | 71 | 70.9 | 70.9 | 70.8 | 71 | 70.7 | 70.8 | 70.6 | 72.5 | 97.6 | 99.94 | Hlr halochloris BN9851 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imhoff, J.F.; Kyndt, J.A.; Meyer, T.E. Genomic Comparison, Phylogeny and Taxonomic Reevaluation of the Ectothiorhodospiraceae and Description of Halorhodospiraceae fam. nov. and Halochlorospira gen. nov. Microorganisms 2022, 10, 295. https://doi.org/10.3390/microorganisms10020295
Imhoff JF, Kyndt JA, Meyer TE. Genomic Comparison, Phylogeny and Taxonomic Reevaluation of the Ectothiorhodospiraceae and Description of Halorhodospiraceae fam. nov. and Halochlorospira gen. nov. Microorganisms. 2022; 10(2):295. https://doi.org/10.3390/microorganisms10020295
Chicago/Turabian StyleImhoff, Johannes F., John A. Kyndt, and Terrance E. Meyer. 2022. "Genomic Comparison, Phylogeny and Taxonomic Reevaluation of the Ectothiorhodospiraceae and Description of Halorhodospiraceae fam. nov. and Halochlorospira gen. nov." Microorganisms 10, no. 2: 295. https://doi.org/10.3390/microorganisms10020295